When Neurons Encounter Nanoobjects: Spotlight on Calcium Signalling

Abstract
:1. Introduction
2. NPs and the Access to the Nervous System
3. NPs and Neuronal Calcium Signalling
4. Environmental NPs (UFPs)

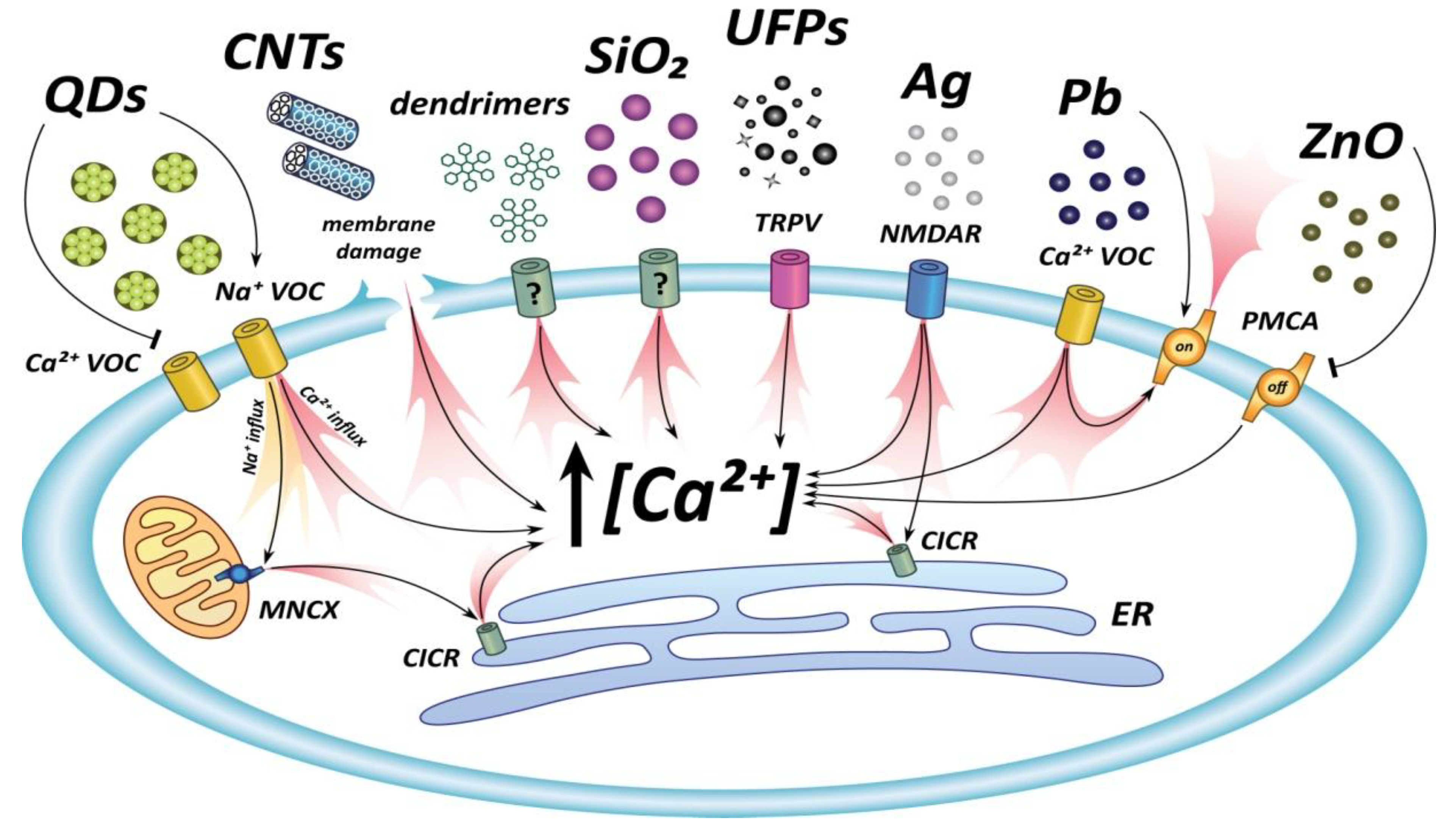{kind=link}
| NP | Diameter | Concentration | Cell Type | Mechanism/Target | [Ca2+]i Response | Ref. |
|---|---|---|---|---|---|---|
| PM (UFPs-FPs) | Nano-micro (0.2–10 µm) | 5–20 µg/mL | Mouse peripheral neurons | Influx through TRPV channels | [Ca2+]i oscillations | [40,41] |
| Ag (PVP coated) | 25 nm | 0.01–2.5 µg/mL | Rat cerebellar neurons | Influx through membrane channels | Increase in basal [Ca2+]i | [31] |
| Ag (PVP coated) | 5–35 nm | ≥2.5 µg/mL | Rat cerebellar neurons | Influx through NMDA receptors; release from intracellular stores | Increase in [Ca2+]i | [45] |
| Ag (peptide coated) | 20–40 nm | 5–20 µg/mL | Rat brain: Glial cells Neurons | n.d. | Increase in [Ca2+]i Oscillatory increase in [Ca2+]i | [44] |
| Au (peptide coated) | 20–40 nm | 5–20 µg/mL | Rat brain: Neurons | -- | No change in [Ca2+]i | [44] |
| PbS | 38 nm | n.d. (chronic uptake by rats) | Rat hippocampal neurons | Increase of expression of L-type calcium channels and of PMCA calcium pumps (compensatory) | Increase in basal [Ca2+]i | [30] |
| ZnO | 30 nm | 2.5–10 µg/mL | Rat retinal neurons | Inhibition of PMCA2 | Increase in basal [Ca2+]i | [48] |
| SiO2 | 50 nm 200 nm | ≤15 µg/mL ≥73 µg/mL ≤150 µg/mL | GT1-7 neuronal line | Influx through membrane channels | Increase in [Ca2+]i small transients strong, sustained increase small transients | [51] |
| Uncoated CdSe QDs | 2.38 nm | ≥10 nM | Rat hippocampal neurons | Calcium influx through Na channels; release from intracellular stores | Sustained [Ca2+]I increase | [53,54] |
| CdSe/ZnS | 10 nm | ≥16 nM (24 h preincubation) | Rat Chromaffin cells | Reduced currents through voltage-dependent calcium channels | n.d. (indirect electrophysiological evidence) | [55] |
| CNTs | 50–100 nm (length > 10 µm) | 30–263 µg/mL | Rat Chromaffin cells | Opening of non-selective channels (membrane damage); altered properties of calcium activated K+ channels | n.d. (indirect electrophysiological evidence) | [59] |
| Dendrimers | nanometers | 0.1 mg/mL | Rat hippocampal slices: glial cells neurons | n.d. n.d. | Increase in [Ca2+]i Transient Long lasting | [62] |
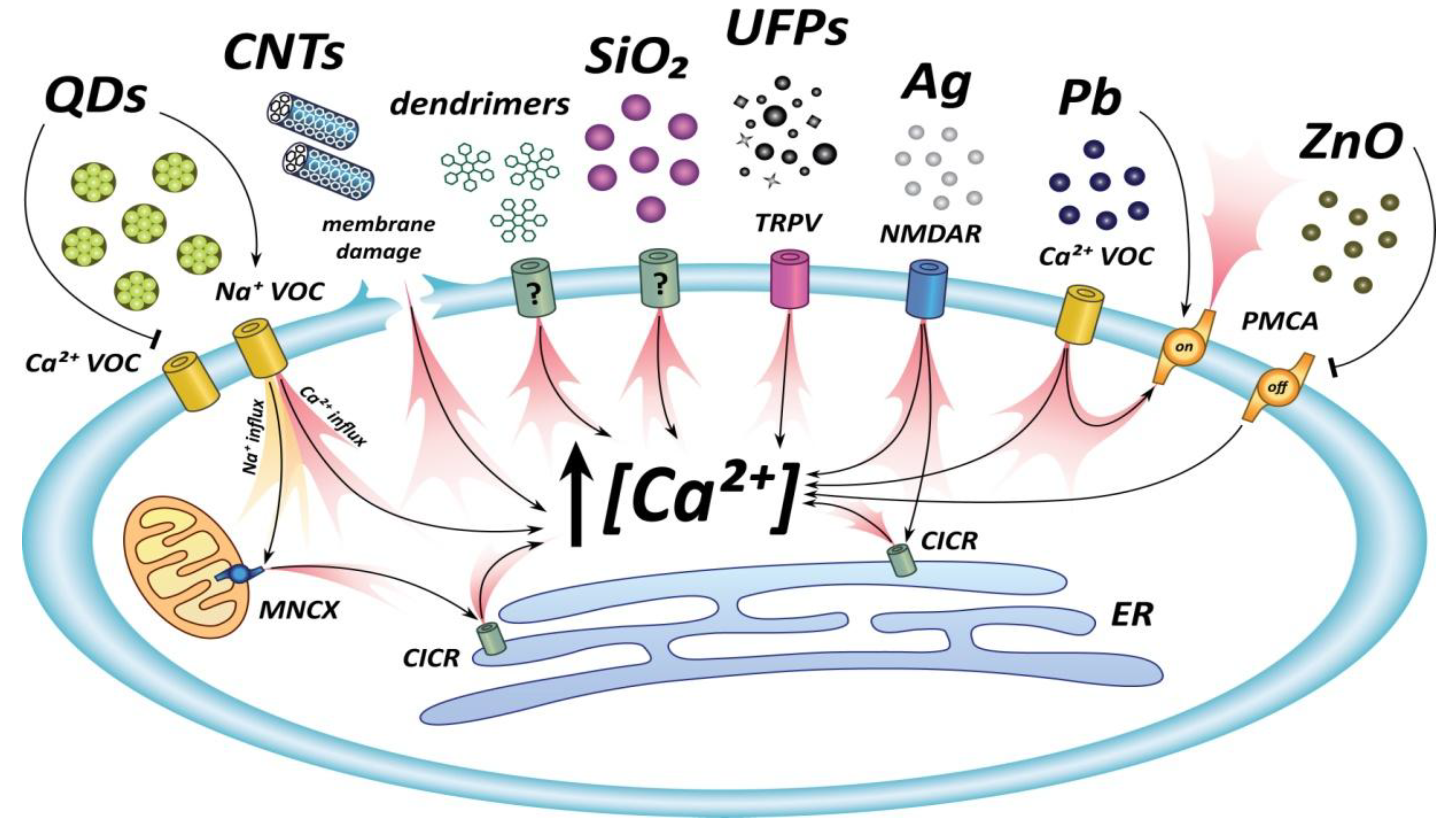
5. Inorganic Nanoparticles
5.1. Silver
5.2. Gold
5.3. Lead Sulphide
5.4. Zinc Oxide
5.5. Silica
5.6. Quantum Dots
5.7. Nanotubes
6. Organic Nanoparticles
7. NPs and Neuronal Networks
8. Conclusions
- -
- in general, most NPs interfere with neuronal calcium homeostasis by interactions at the plasmamembrane; only a few reports of a role of internalization in this context are available.
- -
- accordingly, influx from the extracellular medium is the main mechanism involved in both transient and long lasting increases in [Ca2+]i, with a minor role for release from intracellular compartments;
- -
- generation of ROS, considered as a trigger of neuronal damage and eventually death, has been shown in most cases to be downstream of the changes in [Ca2+]i;
- -
- a relationship can be established between reversible/irreversible or long lasting perturbations in [Ca2+]i and non-toxicity/toxicity.
Abbreviations
| ASICS | acid-sensitive chloride channels |
| BBB | blood-brain barrier |
| CB | carbon black |
| CICR | calcium induced calcium release |
| CNS | central nervous system |
| CNTs | carbon nanotubes |
| FPs | fine particles |
| GnRH | gonadotropin releasing hormone |
| MEAs | multi-electrode arrays |
| MNCX | mitochondrial Na+-Ca2+ exchangers |
| MWCNTs | multiwalled carbon nanotubes |
| NMDA | N-Methyl-d-Aspartate |
| nPM | nanosized particulate matter |
| NPs | nanoparticles |
| NS | nervous system |
| PAMAM | polyamidoammine |
| PM | particulate matter |
| PMCA | plasmamembrane calcium ATPase |
| PVP | polyvinylpirrolidone |
| QDs | quantum dots |
| ROFA | residual oil fly ash |
| ROS | reactive oxygen species |
| SOCE | store operated calcium entry |
| SWCNTs | single walled carbon nanotubes |
| TRPV | transient receptor potential-vanilloid |
| TTX | tetrodotoxin |
| VR1 | vanilloid receptor 1 |
| UFPs | ultrafine particles |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stone, V.; Johnston, H.; Clift, M.J. Air pollution, ultrafine and nanoparticle toxicology: Cellular and molecular interactions. IEEE Trans. Nanobiosci. 2007, 6, 331–340. [Google Scholar] [CrossRef]
- Conner, M.W.; Lam, H.F.; Rogers, A.E.; Fitzgerald, S.; Amdur, M.O. Lung injury in guinea pigs caused by multiple exposures to submicron zinc oxide mixed with sulfur dioxide in a humidified furnace. J. Toxicol. Environ. Health 1985, 16, 101–114. [Google Scholar]
- Davis, D.A.; Akopian, G.; Walsh, J.P.; Sioutas, C.; Morgan, T.E.; Finch, C.E. Urban air pollutants reduce synaptic function of CA1 neurons via an NMDA/NȮ pathway in vitro. J. Neurochem. 2013, 127, 509–519. [Google Scholar] [CrossRef]
- Kubiak, C.; Couvreur, P.; Manil, L.; Clausse, B. Increased cytotoxicity of nanoparticle-carried Adriamycin in vitro and potentiation by verapamil and amiodarone. Biomaterials 1989, 10, 553–556. [Google Scholar]
- Huang, C.C.; Aronstam, R.S.; Chen, D.R.; Huang, Y.W. Oxidative stress, calcium homeostasis, and altered gene expression in human lung epithelial cells exposed to ZnO nanoparticles. Toxicol. In Vitro 2010, 24, 45–55. [Google Scholar] [CrossRef]
- Miyata, R.; van Eeden, S.F. The innate and adaptive immune response induced by alveolar macrophages exposed to ambient particulate matter. Toxicol. Appl. Pharmacol. 2011, 257, 209–226. [Google Scholar] [CrossRef]
- Kendall, M.; Holgate, S. Health impact and toxicological effects of nanomaterials in the lung. Respirology 2012, 17, 743–758. [Google Scholar] [CrossRef]
- Nemmar, A.; Hoylaerts, M.F.; Nemery, B. Effects of particulate air pollution on hemostasis. Clin. Occup. Environ. Med. 2006, 5, 865–881. [Google Scholar]
- Oberdörster, G.; Elder, A.; Rinderknecht, A. Nanoparticles and the brain: Cause for concern? J. Nanosci. Nanotechnol. 2009, 9, 4996–5007. [Google Scholar]
- Karmakar, A.; Zhang, Q.; Zhang, Y. Neurotoxicity of nanoscale materials. J. Food Drug Anal. 2014, 22, 147–160. [Google Scholar] [CrossRef]
- Oberdörster, G.; Sharp, Z.; Atudorei, V.; Elder, A.; Gelein, R.; Kreyling, W.; Cox, C. Translocation of inhaled ultrafine particles to the brain. Inhal. Toxicol. 2004, 16, 437–445. [Google Scholar] [CrossRef]
- Elder, A.; Gelein, R.; Silva, V.; Feikert, T.; Opanashuk, L.; Carter, J.; Potter, R.; Maynard, A.; Ito, Y.; Finkelstein, J.; et al. Translocation of inhaled ultrafine manganese oxide particles to the central nervous system. Environ. Health Perspect. 2006, 114, 1172–1178. [Google Scholar] [CrossRef]
- Sharma, H.S.; Sharma, A. Neurotoxicity of engineered nanoparticles from metals. CNS Neurol. Disord. Drug Targets 2012, 11, 65–80. [Google Scholar] [CrossRef]
- Levesque, S.; Taetzsch, T.; Lull, M.E.; Johnson, J.A.; McGraw, C.; Block, M.L. The role of MAC1 in diesel exhaust particle-induced microglial activation and loss of dopaminergic neuron function. J. Neurochem. 2013, 125, 756–776. [Google Scholar] [CrossRef]
- Davis, D.A.; Bortolato, M.; Godar, S.C.; Sander, T.K.; Iwata, N.; Pakbin, P.; Shih, J.C.; Berhane, K.; McConnell, R.; Sioutas, C.; et al. Prenatal exposure to urban air nanoparticles in mice causes altered neuronal differentiation and depression-like responses. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Allen, J.L.; Liu, X.; Weston, D.; Conrad, K.; Oberdörster, G.; Cory-Slechta, D.A. Consequences of developmental exposure to concentrated ambient ultrafine particle air pollution combined with the adult paraquat and maneb model of the Parkinson’s disease phenotype in male mice. Neurotoxicology 2014, 41, 80–88. [Google Scholar] [CrossRef]
- Allen, J.L.; Conrad, K.; Oberdörster, G.; Johnston, C.J.; Sleezer, B.; Cory-Slechta, D.A. Developmental exposure to concentrated ambient particles and preference for immediate reward in mice. Environ. Health Perspect. 2013, 121, 32–38. [Google Scholar] [CrossRef]
- Block, M.L.; Elder, A.; Auten, R.L.; Bilbo, S.D.; Chen, H.; Chen, J.C.; Cory-Slechta, D.A.; Costa, D.; Diaz-Sanchez, D.; Dorman, D.C.; et al. The outdoor air pollution and brain health workshop. Neurotoxicology 2012, 33, 972–984. [Google Scholar] [CrossRef]
- Gao, H.; Pang, Z.; Jiang, X. Targeted delivery of nano-therapeutics for major disorders of the central nervous system. Pharm. Res. 2013, 30, 2485–2498. [Google Scholar] [CrossRef]
- Kim, J.S.; Yoon, T.J.; Yu, K.N.; Kim, B.G.; Park, S.J.; Kim, H.W.; Lee, K.H.; Park, S.B.; Lee, J.K.; Cho, M.H. Toxicity and tissue distribution of magnetic nanoparticles in mice. Toxicol. Sci. 2006, 89, 338–347. [Google Scholar]
- Shilo, M.; Motiei, M.; Hana, P.; Popovtzer, R. Transport of nanoparticles through the blood-brain barrier for imaging and therapeutic applications. Nanoscale 2014, 6, 2146–2152. [Google Scholar]
- Gendelman, H.E.; Mosley, R.L.; Boska, M.D.; McMillan, J. The promise of nanoneuromedicine. Nanomedicine 2014, 9, 171–176. [Google Scholar] [CrossRef]
- Arundine, M.; Tymianski, M. Molecular mechanisms of calcium-dependent neurodegeneration in excitotoxicity. Cell Calcium 2003, 34, 325–337. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Gleichmann, M.; Mattson, M.P. Neuronal calcium homeostasis and dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Rodríguez, J.J.; Parpura, V. Calcium signalling in astroglia. Mol. Cell. Endocrinol. 2012, 353, 45–56. [Google Scholar] [CrossRef]
- Vandebriel, R.J.; de Jong, W.H. A review of mammalian toxicity of ZnO nanoparticles. Nanotechnol. Sci. Appl. 2012, 5, 61–71. [Google Scholar] [CrossRef]
- Roy, R.; Kumar, S.; Tripathi, A.; Das, M.; Dwivedi, P.D. Interactive threats of nanoparticles to the biological system. Immunol. Lett. 2014, 158, 79–87. [Google Scholar] [CrossRef]
- Huang, J.Y.; Lu, Y.M.; Wang, H.; Liu, J.; Liao, M.H.; Hong, L.J.; Tao, R.R.; Ahmed, M.M.; Liu, P.; Liu, S.S.; et al. The effect of lipid nanoparticle PEGylation on neuroinflammatory response in mouse brain. Biomaterials 2013, 34, 7960–7970. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, H.; Li, Q.; Wang, Q.; Zhang, W.; Chen, Y.; Wang, D.; Cai, Y. Effect of lead sulfide nanoparticles exposure on calcium homeostasis in rat hippocampus neurons. J. Inorg. Biochem. 2013, 126, 70–75. [Google Scholar] [CrossRef]
- Yin, N.; Liu, Q.; Liu, J.; He, B.; Cui, L.; Li, Z.; Yun, Z.; Qu, G.; Liu, S.; Zhou, Q.; et al. Silver nanoparticle exposure attenuates the viability of rat cerebellum granule cells through apoptosis coupled to oxidative stress. Small 2013, 9, 1831–1841. [Google Scholar]
- Miletto, I.; Gilardino, A.; Zamburlin, P.; Dalmazzo, S.; Lovisolo, D.; Caputo, G.; Viscardi, G.; Martra, G. Highly bright and photostable cyanine dye-doped silica nanoparticles. Dyes Pigment. 2010, 84, 121–127. [Google Scholar] [CrossRef]
- Accomasso, L.; Cibrario Rocchietti, E.; Raimondo, S.; Catalano, F.; Alberto, G.; Giannitti, A.; Minieri, V.; Turinetto, V.; Orlando, L.; Saviozzi, S.; et al. Fluorescent silica nanoparticles improveoptical imaging of stem cells allowing direct discrimination between live and early-stage apoptotic cells. Small 2012, 8, 3192–3200. [Google Scholar] [CrossRef]
- Corazzari, I.; Gilardino, A.; Dalmazzo, S.; Fubini, B.; Lovisolo, D. Localization of CdSe/ZnS quantum dots in the lysosomal acidic compartment of cultured neurons and its impact on viability: Potential role of ion release. Toxicol. In Vitro 2013, 27, 752–759. [Google Scholar] [CrossRef]
- Izak-Nau, E.; Kenesei, K.; Murali, K.; Voetz, M.; Eiden, S.; Puntes, V.F.; Duschl, A.; Madarász, E. Interaction of differently functionalized fluorescent silica nanoparticles with neural stem- and tissue-type cells. Nanotoxicology 2013, 12. [Google Scholar] [CrossRef]
- Veronesi, B.; Oortgiesen, M.; Carter, J.D.; Devlin, R.B. Particulate matter initiates inflammatory cytokine release by activation of capsaicin and acid receptors in a human bronchial epithelial cell line. Toxicol. Appl. Pharmacol. 1999, 154, 106–115. [Google Scholar] [CrossRef]
- Alexander, S.P.; Mathie, A.; Peters, J.A. Guide to Receptors and Channels (GRAC), 5th Edition. Br. J. Pharmacol. 2011, 164, S1–S2. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Benham, C.D.; Randall, A.; Davis, J.B. The diversity in the vanilloid (TRPV) receptor family of ion channels. Trends Pharmacol. Sci. 2002, 23, 183–191. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12. [Google Scholar] [CrossRef]
- Oortgiesen, M.; Veronesi, B.; Eichenbaum, G.; Kiser, P.F.; Simon, S.A. Residual oil fly ash and charged polymers activate epithelial cells and nociceptive sensory neurons. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L683–L695. [Google Scholar]
- Veronesi, B.; de Haar, C.; Roy, J.; Oortgiesen, M. Particulate matter inflammation and receptor sensitivity are target cell specific. Inhal. Toxicol. 2002, 14, 159–183. [Google Scholar] [CrossRef]
- Gillespie, P.; Tajuba, J.; Lippmann, M.; Chen, L.C.; Veronesi, B. Particulate matter neurotoxicity in culture is size-dependent. Neurotoxicology 2013, 36, 112–117. [Google Scholar] [CrossRef]
- Chen, X.; Schluesener, H.J. Nanosilver: A nanoproduct in medical application. Toxicol. Lett. 2008, 176, 1–12. [Google Scholar] [CrossRef]
- Haase, A.; Rott, S.; Mantion, A.; Graf, P.; Plendl, J.; Thünemann, A.F.; Meier, W.P.; Taubert, A.; Luch, A.; Reiser, G. Effects of silver nanoparticles on primary mixed neural cell cultures: Uptake, oxidative stress and acute calcium responses. Toxicol. Sci. 2012, 126, 457–468. [Google Scholar]
- Ziemińska, E.; Stafiej, A.; Strużyńska, L. The role of the glutamatergic NMDA receptor in nanosilver-evoked neurotoxicity in primary cultures of cerebellar granule cells. Toxicology 2014, 315, 38–48. [Google Scholar] [CrossRef]
- Alon, N.; Miroshnikov, Y.; Perkas, N.; Nissan, I.; Gedanken, A.; Shefi, O. Substrates coated with silver nanoparticles as a neuronal regenerative material. Int. J. Nanomed. 2014, 9, 23–31. [Google Scholar]
- Paviolo, C.; Haycock, J.W.; Cadusch, P.J.; McArthur, S.L.; Stoddart, P.R. Laser exposure of gold nanorods can induce intracellular calcium transients. J. Biophotonics 2013, 6. [Google Scholar] [CrossRef]
- Guo, D.; Bi, H.; Wang, D.; Wu, Q. Zinc oxide nanoparticles decrease the expression and activity of plasma membrane calcium ATPase, disrupt the intracellular calcium homeostasis in rat retinal ganglion cells. Int. J. Biochem. Cell Biol. 2013, 45, 1849–1859. [Google Scholar] [CrossRef]
- Wang, H.J.; Growcock, A.C.; Tang, T.H.; O’Hara, J.; Huang, Y.W.; Aronstam, R.S. Zinc oxide nanoparticle disruption of store-operated calcium entry in a muscarinic receptor signaling pathway. Toxicol. In Vitro 2010, 24, 1953–1961. [Google Scholar]
- Lu, C.W.; Hung, Y.; Hsiao, J.K.; Yao, M.; Chung, T.H.; Lin, Y.S.; Wu, S.H.; Hsu, S.C.; Liu, H.M.; Mou, C.Y.; et al. Bifunctional magnetic silica nanoparticles for highly efficient human stem cell labeling. Nano Lett. 2007, 7, 149–154. [Google Scholar] [CrossRef]
- Ariano, P.; Zamburlin, P.; Gilardino, A.; Mortera, R.; Onida, B.; Tomatis, M.; Ghiazza, M.; Fubini, B.; Lovisolo, D. Interaction of spherical silica nanoparticles with neuronal cells: Size-dependent toxicity and perturbation of calcium homeostasis. Small 2011, 7, 766–774. [Google Scholar] [CrossRef]
- Shao, L.; Gao, Y.; Yan, F. Semiconductor quantum dots for biomedicial applications. Sensors 2011, 11, 11736–11751. [Google Scholar] [CrossRef]
- Tang, M.; Xing, T.; Zeng, J.; Wang, H.; Li, C.; Yin, S.; Yan, D.; Deng, H.; Liu, J.; Wang, M.; et al. Unmodified CdSe quantum dots induce elevation of cytoplasmic calcium levels and impairment of functional properties of sodium channels in rat primary cultured hippocampal neurons. Environ. Health Perspect. 2008, 116, 915–922. [Google Scholar] [CrossRef]
- Tang, M.; Wang, M.; Xing, T.; Zeng, J.; Wang, H.; Ruan, D.Y. Mechanisms of unmodified CdSe quantum dot-induced elevation of cytoplasmic calcium levels in primary cultures of rat hippocampal neurons. Biomaterials 2008, 29, 4383–4391. [Google Scholar] [CrossRef]
- Gosso, S.; Gavello, D.; Giachello, C.N.; Franchino, C.; Carbone, E.; Carabelli, V. The effect of CdSe-ZnS quantum dots on calcium currents and catecholamine secretion in mouse chromaffin cells. Biomaterials 2011, 32, 9040–9050. [Google Scholar]
- Fabbro, A.; Bosi, S.; Ballerini, L.; Prato, M. Carbon nanotubes: Artificial nanomaterials to engineer single neurons and neuronal networks. ACS Chem. Neurosci. 2012, 3, 611–618. [Google Scholar]
- Fattahi, P.; Yang, G.; Kim, G.; Abidian, M.R. A review of organic and inorganic biomaterials for neural interfaces. Adv. Mater. 2014, 26, 1846–1885. [Google Scholar]
- Park, S.Y.; Kang, B.S.; Hong, S. Improved neural differentiation of human mesenchymal stem cells interfaced with carbon nanotube scaffolds. Nanomedicine 2013, 8, 715–723. [Google Scholar] [CrossRef]
- Gavello, D.; Vandael, D.H.; Cesa, R.; Premoselli, F.; Marcantoni, A.; Cesano, F.; Scarano, D.; Fubini, B.; Carbone, E.; Fenoglio, I.; et al. Altered excitability of cultured chromaffin cells following exposure to multi-walled carbon nanotubes. Nanotoxicology 2012, 6, 47–60. [Google Scholar] [CrossRef]
- Jakubek, L.M.; Marangoudakis, S.; Raingo, J.; Liu, X.; Lipscombe, D.; Hurt, R.H. The inhibition of neuronal calcium ion channels by trace levels of yttrium released from carbon nanotubes. Biomaterials 2009, 30, 6351–6537. [Google Scholar] [CrossRef]
- Huang, R.Q.; Qu, Y.H.; Ke, W.L.; Zhu, J.H.; Pei, Y.Y.; Jiang, C. Efficient gene delivery targeted to the brain using a transferrin-conjugated polyethyleneglycol modified polyamidoamine dendrimer. FASEB J. 2007, 21, 1117–1125. [Google Scholar] [CrossRef]
- Nyitrai, G.; Héja, L.; Jablonkai, I.; Pál, I.; Visy, J.; Kardos, J. Polyamidoamine dendrimer impairs mitochondrial oxidation in brain tissue. J. Nanobiotechnology 2013, 11. [Google Scholar] [CrossRef]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981, 391, 85–100. [Google Scholar] [CrossRef]
- Gramowski, A.; Flossdorf, J.; Bhattacharya, K.; Jonas, L.; Lantow, M.; Rahman, Q.; Schiffmann, D.; Weiss, D.G.; Dopp, E. Nanoparticles induce changes of the electrical activity of neuronal networks on microelectrode array neurochips. Environ. Health Perspect. 2010, 118, 1363–1369. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lovisolo, D.; Gilardino, A.; Ruffinatti, F.A. When Neurons Encounter Nanoobjects: Spotlight on Calcium Signalling. Int. J. Environ. Res. Public Health 2014, 11, 9621-9637. https://doi.org/10.3390/ijerph110909621
Lovisolo D, Gilardino A, Ruffinatti FA. When Neurons Encounter Nanoobjects: Spotlight on Calcium Signalling. International Journal of Environmental Research and Public Health. 2014; 11(9):9621-9637. https://doi.org/10.3390/ijerph110909621
Chicago/Turabian StyleLovisolo, Davide, Alessandra Gilardino, and Federico Alessandro Ruffinatti. 2014. "When Neurons Encounter Nanoobjects: Spotlight on Calcium Signalling" International Journal of Environmental Research and Public Health 11, no. 9: 9621-9637. https://doi.org/10.3390/ijerph110909621
APA StyleLovisolo, D., Gilardino, A., & Ruffinatti, F. A. (2014). When Neurons Encounter Nanoobjects: Spotlight on Calcium Signalling. International Journal of Environmental Research and Public Health, 11(9), 9621-9637. https://doi.org/10.3390/ijerph110909621