Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula



Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Analyses
2.2. Anticoagulant Activity by Activated Partial Thromboplastin Time (aPTT) and Prothrombin Time (PT) Assays
2.3. Antioxidant Activity
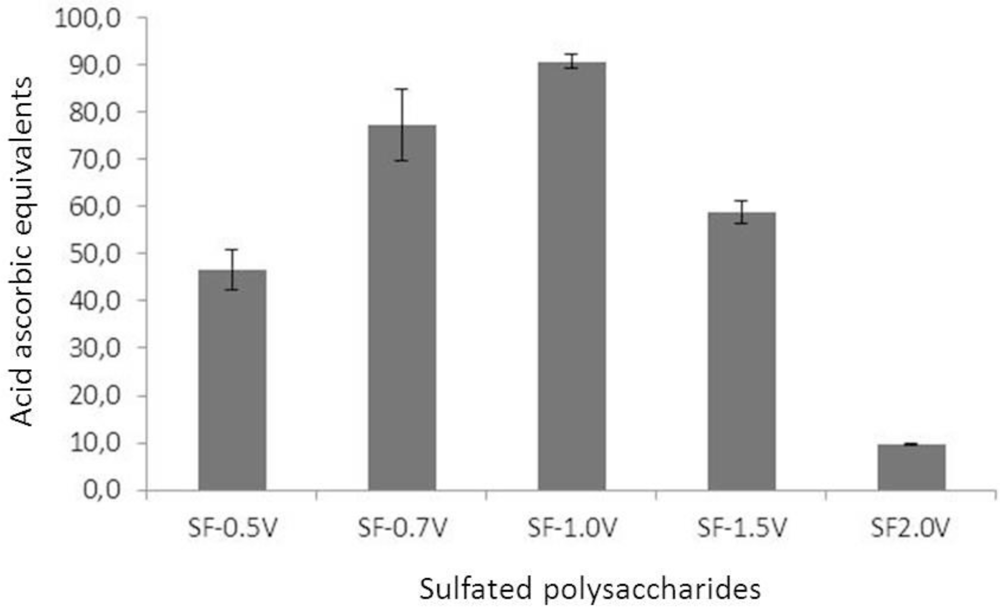
2.3.1. Total Antioxidant Capacity (TAC)
2.3.2. Hydroxyl and Superoxide Radical Scavenging
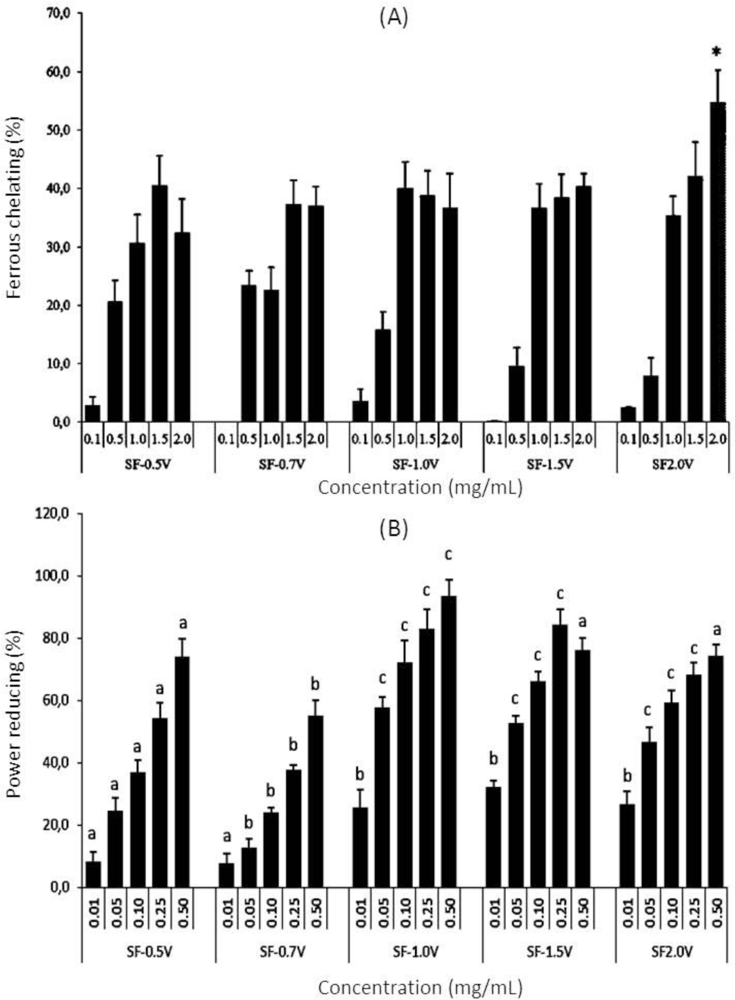
2.3.3. Chelating Effect on Ferrous Ions
2.3.4. Reducing Power
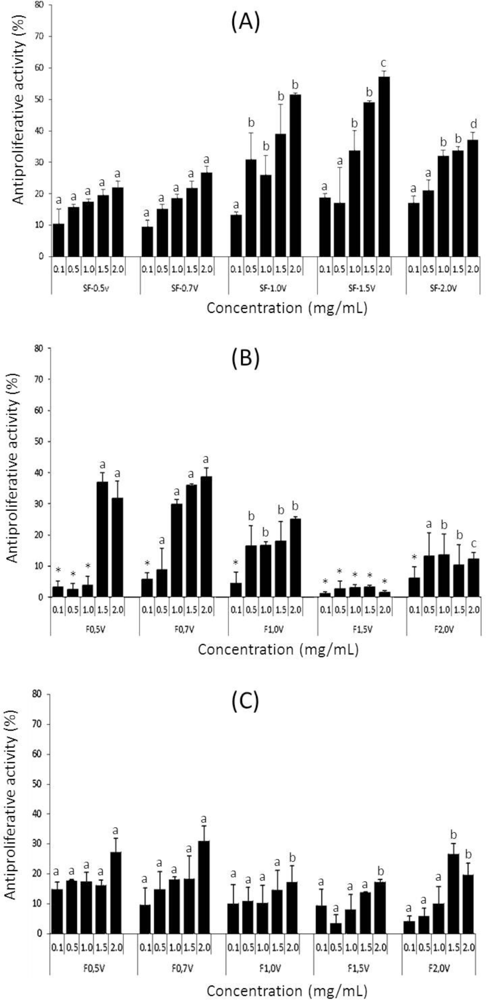
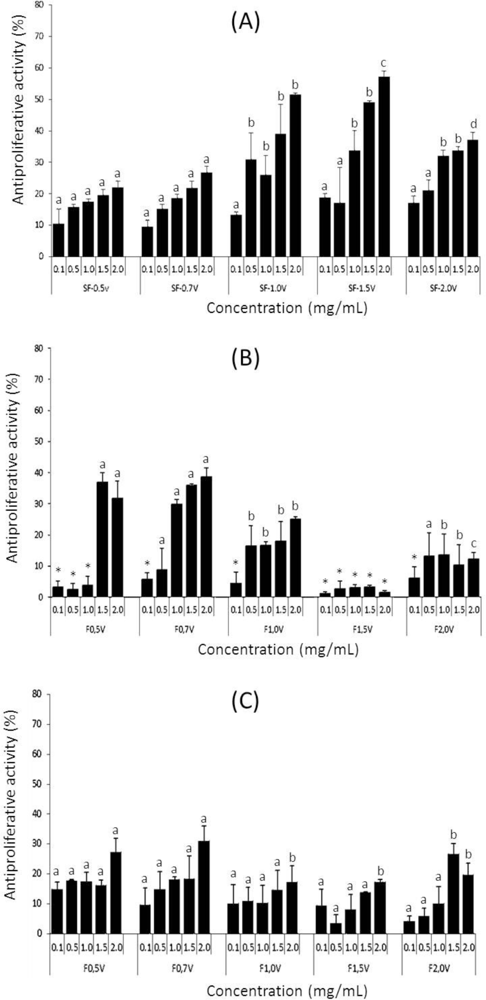
2.4. Antiproliferative Activity
3. Experimental Section
3.1. Materials
3.2. Extraction of Sulfated Polysaccharide
3.3. Chemical Analysis and Monosaccharide Composition
3.4. Molecular Weight Determination
3.5. Anticoagulant Activity
3.6. Antioxidant Activity
3.6.1. Determination of Total Antioxidant Capacity
3.6.2. Hydroxyl Radical Scavenging Activity Assay
3.6.3. Superoxide Radical Scavenging Activity Assay
3.6.4. Ferrous Ion [Fe(II)] Chelating Activity
3.6.5. Reducing Power
3.7. Antiproliferative Activity
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Almeida-Lima, J; Costa, LS; Silva, NB; Melo-Silveira, RF; Silva, FV; Felipe, MBMC; Medeiros, SRBM; Leite, EL; Rocha, HAO. Evaluating the possible genotoxic, mutagenic and tumor cell proliferation-inhibition effects of a non-anticoagulant, but antithrombotic algal heterofucan. J Appl Toxicol 2010, 30, 708–715. [Google Scholar]
- Costa, LS; Fidelis, GP; Cordeiro, SL; Oliveira, RM; Sabry, DA; Câmara, RBG; Nobre, LTDB; Costa, MSSP; Almeida-Lima, J; Farias, EHC; Leite, EL; Rocha, HAO. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed Pharmacother 2010, 64, 21–28. [Google Scholar]
- Barroso, EM; Costa, LS; Medeiros, VP; Cordeiro, SL; Costa, MS; Franco, CR; Nader, HB; Leite, EL; Rocha, HAO. A non-anticoagulant heterofucan has antithrombotic activity in vivo. Planta Med 2008, 74, 712–718. [Google Scholar]
- Cumashi, A; Ushakova, NA; Preobrazhenskaya, ME; D’Incecco, A; Picooli, A; Totani, L; Tinari, N; Morozevich, GE; Berman, AE; Bilan, MI; Usov, AI; Ustyuzhanina, NE; Grachev, AA; Sanderson, CJ; Kelly, M; Rabinovich, GA; Iacobelli, S; Nifantiev, NE. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Hoshino, T; Hayashi, T; Hayashi, K; Hamada, J; Lee, JB; Sankawa, U. An antivirally active sulfated polysaccharide from Sargassum horneri (TURNER) C. AGARDH. Biol Pharm Bull 1998, 21, 730–734. [Google Scholar]
- Sinha, S; Astani, A; Ghosh, T; Schnitzler, P; Ray, B. Polysaccharides from Sargassum tenerrimum: structural features, chemical modification and anti-viral activity. Phytochemistry 2010, 71, 235–242. [Google Scholar]
- Zhu, W; Ooi, VEC; Chan, PKS; Ang, POJ. Isolation and characterization of a sulfated polysaccharide from the brown alga Sargassum patens and determination of its anti-herpes activity. Biochem Cell Biol 2003, 81, 25–33. [Google Scholar]
- Stevan, FR; Oliveira, MB; Bucchi, DF; Noseda, MD; Iacomini, M; Duarte, ME. Cytotoxic effects against HeLa cells of polysaccharides from seaweeds. J Submicrosc Cytol Pathol 2001, 33, 477–484. [Google Scholar]
- Josephine, A; Veena, CK; Amudha, G; Preetha, SP; Sundarapandian, R; Varalakshmi, P. Sulphated polysaccharides: new insight in the prevention of cyclosporine A-induced glomerular injury. Basic Clin Pharmacol Toxicol 2007, 101, 9–15. [Google Scholar]
- Rocha, HAO; Moraes, FA; Trindade, ES; Franco, CR; Torquato, RJ; Veiga, SS; Valente, AP; Mourão, PA; Leite, EL; Nader, HB; Dietrich, CP. Structural and hemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schröederi. An ideal antithrombotic agent? J Biol Chem 2005, 280, 41278–41288. [Google Scholar]
- Rioux, LE; Turgeon, SL; Beaulieu, M. Structural characterization of laminaran and galactofucan extracted from the brown seaweed. Saccharina longicrucis Phytochemistry 2010, 71, 1586–1595. [Google Scholar]
- Medeiros, VP; Queiroz, KC; Cardoso, ML; Monteiro, GR; Oliveira, FW; Chavante, SF; Guimaraes, LA; Rocha, HA; Leite, EL. Sulfated galactofucan from Lobophora variegata: anticoagulant and anti-inflammatory properties. Biochemistry (Mosc) 2008, 73, 1018–1024. [Google Scholar]
- Ponce, NMA; Pujol, CA; Damonte, EB; Flores, ML; Stortz, CA. Fucoidans from the brown seaweed Adenocystis utricularis: extraction methods, antiviral activity and structural studies. Carbohydr Res 2003, 338, 153–165. [Google Scholar]
- Duarte, MER; Cardoso, MA; Noseda, MD; Cerezo, AS. Structural studies on fucoidans from the brown seaweed. Sargassum stenophyllum Carbohydr Res 2001, 333, 281–293. [Google Scholar]
- Kamakar, P; Ghosh, T; Sinha, S; Saha, S; Mandal, P; Ghosal, KG; Ray, B. Polysaccharides from the brown seaweed Padina tetrastromatica: Characterization of a sulfated fucan. Carbohydr Polym 2009, 78, 416–421. [Google Scholar]
- Zhang, Z; Wang, F; Wang, X; Liu, X; Hou, Y; Zhang, Q. Extraction of the polysaccharides from five algae and their potential antioxidant activity in vitro. Carbohydr Polym 2010, 82, 118–121. [Google Scholar]
- Pomim, VK. Review: an overview about the structure-function relationship of marine sulfated homopolysaccharides with regular chemical structures. Biopolymers 2009, 91, 601–609. [Google Scholar]
- Li, B; Lu, F; Wei, X; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Zhang, Q; Yu, P; Li, Z; Zhang, H; Xu, Z; Li, P. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J Appl Phycol 2003, 15, 301–310. [Google Scholar]
- Qi, H; Zhang, Q; Zhao, T; Chen, R; Zhang, H; Niu, X; Li, Z. Antioxidant activity of different sulfate content derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta) in vitro. Int J Biol Macromol 2005, 37, 195–199. [Google Scholar]
- Souza, MCR; Marques, CT; Dore, CMG; Silva, FRF; Rocha, HAO; Leite, EL. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J Appl Phycol 2007, 19, 153–160. [Google Scholar]
- Hu, T; Liu, L; Chen, Y; Wu, J; Wang, S. Antioxidant activity of sulfated polysaccharide fractions extracted from Undaria pinnatifida in vitro. Int J Biol Macromol 2010, 46, 193–198. [Google Scholar]
- Camara, RBG; Costa, LS; Fidelis, GP; Nobre, LTDB; Dantas-Santos, N; Cordeiro, SL; Costa, MSSP; Alves, LG; Rocha, HAO. Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities. Mar Drugs 2011, 9, 124–138. [Google Scholar]
- Zhang, Z; Zhang, Q; Wang, J; Zhang, H; Niu, X; Li, P. Preparation of the different derivatives of the low-molecular-weight porphyran from Porphyra haitanensis and their antioxidant activities in vitro. Int J Biol Macromol 2009, 45, 22–26. [Google Scholar]
- Nagamine, T; Hayakawa, K; Kusakabe, T; Takada, H; Nakazato, K; Hisanaga, E; Iha, M. Inhibitory Effect of Fucoidan on Huh7 Hepatoma Cells Through Downregulation of CXCL12. Nut Can 2009, 61, 340–347. [Google Scholar]
- Jiang, Z; Okimura, T; Yokose, T; Yamasaki, Y; Yamaguchi, K; Oda, T. Effects of sulfated fucan, ascophyllan, from the brown Alga Ascophyllum nodosum on various cell lines: A comparative study on ascophyllan and fucoidan. J Biosc Bioeng 2010, 110, 113–117. [Google Scholar]
- Lin, E; Chou, H; Kuo, P; Huang, Y. Antioxidant and antiproliferative activities of methanolic extracts of Perilla frutescens. J Med Plant Res 2010, 4, 477–483. [Google Scholar]
- Holtkamp, AD; Kelly, S; Ulber, R; Lang, S. Fucoidans and fucoidanases-focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl Microbiol Biotechnol 2009, 82, 1–11. [Google Scholar]
- Ye, J; Li, Y; Teruya, K; Katakura, Y; Ichikawa, A; Eto, H; Hosoi, M; Hosoi, M; Nishimoto, S; Shirahata, S. Enzyme-digested fucoidan extracts derived from seaweed mozuku of cladosiphon novae-caledoniae kylin inhibit invasion and angiogenesis of tumor cells. Cytotechnology 2005, 47, 117–126. [Google Scholar]
- Dubois, M; Gilles, KA; Hamilton, JK; Rebers, PA; Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956, 28, 250–256. [Google Scholar]
- Dodgson, KS; Price, RG. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem J 1962, 84, 106–110. [Google Scholar]
- Spector, J. Refinement of the coomassie blue method of protein quantification. A simple and linear spectrophotometric assay of 0.5 to 50 μg of protein. Anal Biochem 1978, 86, 142–143. [Google Scholar]
- Somogyi, M. Notes on sugar determination. J Biol Chem 1952, 195, 19–23. [Google Scholar]
- Farndale, RW; Buttle, DJ; Barrett, A. Improved quantitation and discrimination of sulphated glycosaminoglycans by use of dimethylmethylene blue. J Biochim Biophys Acta 1986, 883, 173–177. [Google Scholar]
- Albuquerque, IRL; Queiroz, KCS; Alves, LG; Santos, EA; Leite, EL; Rocha, HAO. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz J Med Biol Res 2004, 37, 167–171. [Google Scholar]
- Silva, GDF; Silva, SRS; Barbosa, LCA; Duarte, LP; Ribeiro, SMR; Queiroz, JH; Vieira, SA; Oliveira, ML. Antioxidant activity of Maytenus imbricata Mart., Celastraceae. Rev Bras Farmacogn 2009, 19, 530–536. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Sulfated Polysaccharides | Total sugar (%) | Sulfate (%) | Protein (%) | Molar ratio | |||||
|---|---|---|---|---|---|---|---|---|---|
| Fuc | Gal | Glc | Man | Xyl | Gluc acid | ||||
| SF-0.5v | 41.4 | 10.2 | 0.6 | 1.0 | 1.5 | 0.5 | 0.4 | 1.0 | 1.1 |
| SF-0.7v | 46.2 | 10.8 | 0.5 | 1.0 | 1.2 | 0.7 | 0.2 | 0.7 | 0.7 |
| SF-1.0v | 59.1 | 12.6 | 0.3 | 1.0 | 1.3 | 0.5 | 0.1 | 0.3 | 0.7 |
| SF-1.5v | 64.9 | 12.3 | 0.2 | 1.0 | 1.1 | 0.3 | - | 0.1 | 0.5 |
| SF-2.0v | 66.0 | 17.7 | 0.4 | 1.0 | 2.2 | 0.5 | 0.6 | 0.2 | - |
| Sulfated polysaccharides | Concentration (mg/mL) | Scavenging (%) | |
|---|---|---|---|
| OH• | O2− | ||
| SF-0.5v | 0.05 | 0 ± 0 | 0 ± 0 |
| 0.10 | 0 ± 0 | 0 ± 0 | |
| 0.25 | 0 ± 0 | 0 ± 0 | |
| 0.50 | 0 ± 0 | 0 ± 0 | |
| SF-0.7v | 0.05 | 5.3 ± 3.5 a | 11.1 ± 0.4 a |
| 0.10 | 15.8 ± 2.9 b | 14.2 ± 0.4 b | |
| 0.25 | 23.0 ± 1.5 c | 16.3 ± 0.6 b | |
| 0.50 | 26.2 ± 1.8 c | 19.3 ± 0.7 b | |
| SF-1.0v | 0.05 | 10.8 ± 2.2 a | 0 ± 0 |
| 0.10 | 17.1 ± 2.9 b | 0 ± 0 | |
| 0.25 | 22.5 ± 1.5 c | 0 ± 0 | |
| 0.50 | 26.7 ± 1.8 c | 0 ± 0 | |
| SF-1.5v | 0.05 | 4.9 ± 0.9 a | 0 ± 0 |
| 0.10 | 9.2 ± 0.7 b | 0 ± 0 | |
| 0.25 | 12.4 ± 1.5 b | 0 ± 0 | |
| 0.50 | 12.7 ± 4.8 b | 0 ± 0 | |
| SF-2.0v | 0.05 | 0 ± 0 | 0 ±0 |
| 0.10 | 0 ± 0 | 5.0 ± 0.7 a | |
| 0.25 | 0 ± 0 | 9.0 ± 1.1 b | |
| 0.50 | 0 ± 0 | 12.2 ± 1.2 b | |
| Gallic acid | 0.05 | 11.6 ± 1,7 a | 28.9 ± 3.8 a |
| 0.10 | 43.6 ± 2.4 b | 41.8 ± 4.7 b | |
| 0.25 | 64.3 ± 3.0 c | 72.1 ± 2.9 c | |
| 0.50 | 93.7 ± 3.7 d | 86.3 ± 3.1 d | |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Costa, L.S.; Fidelis, G.P.; Telles, C.B.S.; Dantas-Santos, N.; Camara, R.B.G.; Cordeiro, S.L.; Costa, M.S.S.P.; Almeida-Lima, J.; Melo-Silveira, R.F.; Oliveira, R.M.; et al. Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952-966. https://doi.org/10.3390/md9060952
Costa LS, Fidelis GP, Telles CBS, Dantas-Santos N, Camara RBG, Cordeiro SL, Costa MSSP, Almeida-Lima J, Melo-Silveira RF, Oliveira RM, et al. Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula. Marine Drugs. 2011; 9(6):952-966. https://doi.org/10.3390/md9060952
Chicago/Turabian StyleCosta, Leandro Silva, Gabriel Pereira Fidelis, Cinthia Beatrice Silva Telles, Nednaldo Dantas-Santos, Rafael Barros Gomes Camara, Sara Lima Cordeiro, Mariana Santana Santos Pereira Costa, Jailma Almeida-Lima, Raniere Fagundes Melo-Silveira, Ruth Medeiros Oliveira, and et al. 2011. "Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula" Marine Drugs 9, no. 6: 952-966. https://doi.org/10.3390/md9060952
APA StyleCosta, L. S., Fidelis, G. P., Telles, C. B. S., Dantas-Santos, N., Camara, R. B. G., Cordeiro, S. L., Costa, M. S. S. P., Almeida-Lima, J., Melo-Silveira, R. F., Oliveira, R. M., Albuquerque, I. R. L., Andrade, G. P. V., & Rocha, H. A. O. (2011). Antioxidant and Antiproliferative Activities of Heterofucans from the Seaweed Sargassum filipendula. Marine Drugs, 9(6), 952-966. https://doi.org/10.3390/md9060952