Effects of High Salt Stress on Secondary Metabolite Production in the Marine-Derived Fungus Spicaria elegans

Abstract
:1. Introduction
2. Results and Discussion
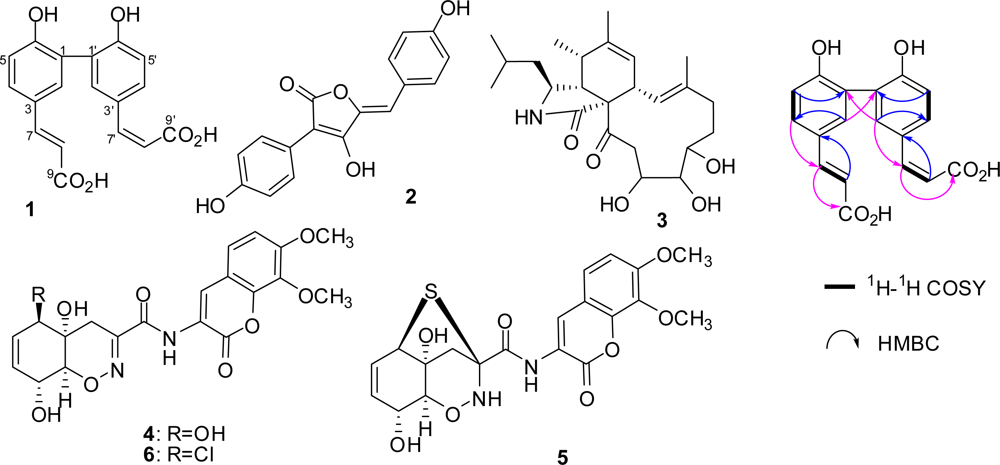
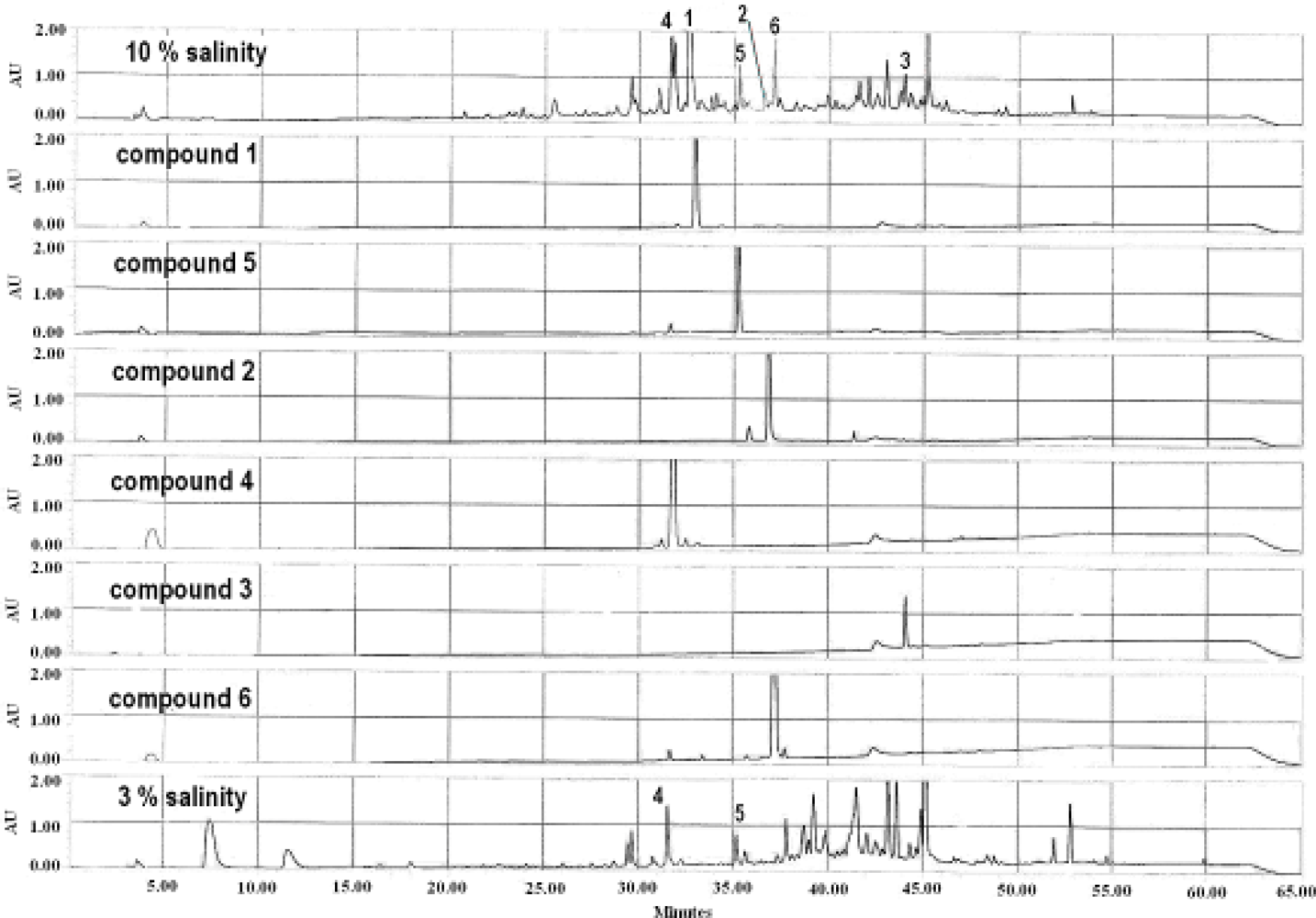
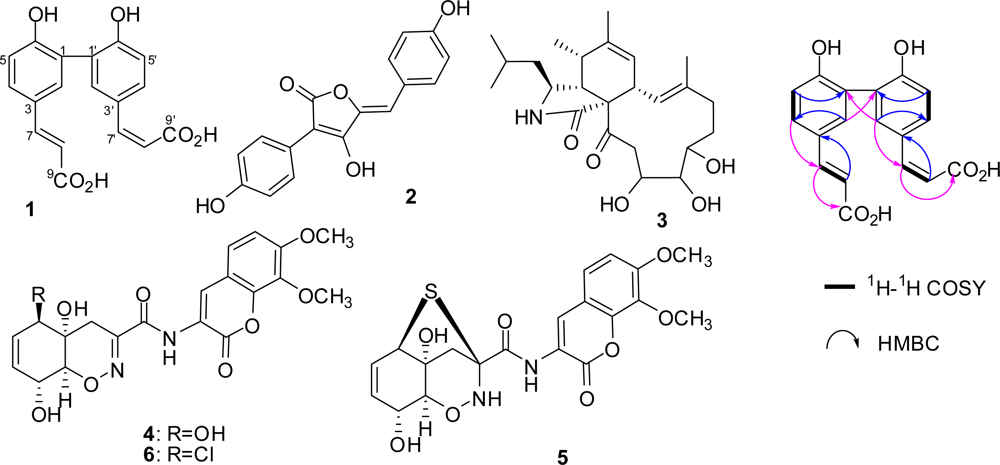
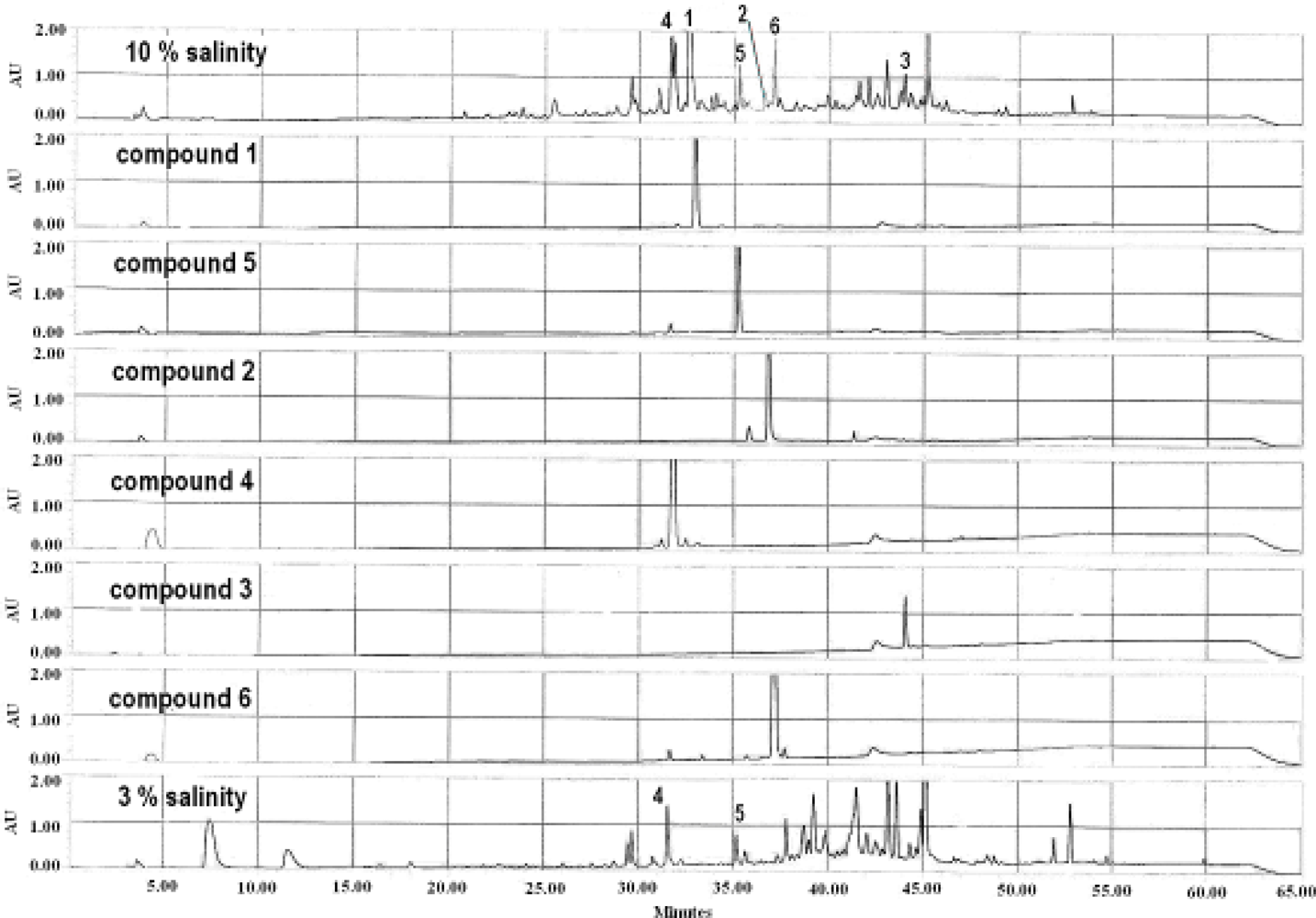
2.1. Identification of Metabolites from S. Elegans in 10% Saline Conditions
2.2. Effect of Salt Stress on Secondary Metabolite Production
2.3. Antimicrobial Effects of Secondary Metabolites
3. Experimental Section
3.1. General Experimental Procedures
3.2. Fungal Material
3.3. Fermentation and Extraction
3.4. Purification
3.5. Bioassays
4. Conclusions
Acknowledgments
References
- Larsen, TO; Smedsgaard, J; Nielsen, KF; Hansen, ME; Frisvad, JC. Phenotypic taxonomy and metabolite profiling in microbial drug discovery. Nat Prod Rep 2005, 22, 672–695. [Google Scholar]
- Liu, R; Gu, QQ; Zhu, WM; Cui, CB; Fan, GT. Trichodermamide A and aspergillazine A, two cytotoxic modified dipeptides from a marine-derived fungus. Spicaria elegans. Arch Pharm Res 2005, 28, 1042–1046. [Google Scholar]
- Liu, R; Gu, QQ; Zhu, WM; Cui, CB; Fan, GT; Fang, YC; Zhu, TJ; Liu, HB. 10-Phenyl-[12]-cytochalasins Z7, Z8, and Z9 from the marine-derived fungus. Spicaria elegans. J Nat Prod 2006, 69, 871–875. [Google Scholar]
- Liu, R; Lin, ZJ; Zhu, TJ; Fang, YC; Gu, QQ; Zhu, WM. Novel open-chain cytochalsins from the marine-derived fungus. Spicaria elegans. J Nat Prod 2008, 71, 1127–1132. [Google Scholar]
- Wang, WL; Lu, ZY; Tao, HW; Zhu, TJ; Fang, YC; Gu, QQ; Zhu, WM. Isoechinulin-type alkaloids, variecolorins A-L, from halotolerant. Aspergillus variecolor. J Nat Prod 2007, 70, 1558–1564. [Google Scholar]
- Wang, WL; Zhu, TJ; Tao, HW; Lu, ZY; Fang, YC; Gu, QQ; Zhu, WM. Three novel, structurally unique spirocyclic alkaloids from the halotolerant B-17 fungal strain of Aspergillus variecolor. Chem Biodivers 2007, 4, 2913–2919. [Google Scholar]
- Lu, ZY; Lin, ZJ; Wang, WL; Du, L; Zhu, TJ; Fang, YC; Gu, QQ; Zhu, WM. Citrinin dimers from the halotolerant fungus Penicillium citrinum B-57. J Nat Prod 2008, 71, 543–546. [Google Scholar]
- Zheng, JK; Zhu, HJ; Hong, K; Wang, Y; Liu, PP; Wang, X; Peng, XP; Zhu, WM. Novel cyclic hexapeptides from marine-derived fungus, Aspergillus sclerotiorum PT06-1. Org Lett 2009, 11, 5262–5265. [Google Scholar]
- Zheng, JK; Xu, ZH; Wang, Y; Hong, K; Liu, PP; Zhu, WM. Cyclic tripeptides from the halotolerant fungus Aspergillus sclerotiorum PT06-1. J Nat Prod 2010, 73, 1133–1137. [Google Scholar]
- Sugiyama, H; Ojima, N; Kobayashi, M; Senda, Y; Ishiyama, J; Seto, S. C-13 NMR Spectra of Aspulvinones. Agric Biol Chem 1979, 43, 403–404. [Google Scholar]
- Naruse, N; Yamamoto, H; Murata, S; Sawada, Y; Fukagawa, Y; Oki, T. Aspochalasin E, a new antibiotic isolated from a fungus. J Antibiot 1993, 46, 679–681. [Google Scholar]
- Garo, E; Starks, CM; Jensen, PR; Fenical, W; Lobkovsky, E; Clardy, J. Trichodermamides A and B, cytotoxic modified dipeptides from the marine-derived fungus Trichoderma virens. J Nat Prod 2003, 66, 423–426. [Google Scholar]
- Lin, ZJ; Zhu, TJ; Zhang, GJ; Wei, HJ; Gu, QQ. Deoxy-cytochalasins from a marine-derived fungus Spicaria elegans. Can J Chem 2009, 87, 486–489. [Google Scholar]
- Lin, ZJ; Zhu, TJ; Chen, L; Gu, QQ. Three new aspochalasin derivatives from the marine-derived fungus Spicaria elegans. Chin Chem Lett 2010, 21, 824–826. [Google Scholar]
- Tresner, HD; Hayes, JA. Sodium chloride tolerance of terrestrial fungi. Appl Microbiol 1971, 22, 210–213. [Google Scholar]
- Yu, JH; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu Rev Phytopathol 2005, 43, 437–458. [Google Scholar]
- Zaika, LL. Spices and herbs: Their antimicrobial activity and its determination. J Food Safety 1988, 9, 97–118. [Google Scholar]

{kind=link}
{kind=link}
| MIC (mM) | |||||
|---|---|---|---|---|---|
| Compound | Enterobacter aerogenes | Escherichia coli | Pseudomonas aeruginosa | Staphylococcus aureus | Candida albicans |
| 1 | 0.153 | 0.038 | 0.767 | 1.534 | 0.383 |
| 2 | 0.084 | 0.338 | 3.378 | 3.378 | 0.844 |
| 3 | 0.059 | 0.239 | 1.193 | 2.387 | 0.298 |
| 4 | 0.029 | 0.231 | 0.231 | 0.231 | 0.289 |
| 5 | 0.056 | 0.223 | 2.227 | 0.223 | 0.557 |
| 6 | 0.056 | 0.222 | 0.222 | 0.222 | 0.278 |
| Ciprofloxacin lactate | 0.001 | 0.004 | 0.030 | 0.001 | − |
| Ketoconazole | − | − | − | − | 0.005 |
| position | δC | δH (J in Hz) | HMBC (H→C) | 1H-1H COSY |
|---|---|---|---|---|
| 1 | 126.0 s | |||
| 2 | 131.9 d | 7.40 (1H, d, 2.3) | 1′, 4, 7 | |
| 3 | 125.0 s | |||
| 4 | 128.6 d | 7.52 (1H, dd, 8.7, 2.3) | 2, 6, 7 | 5 |
| 5 | 116.0 d | 6.91 (1H, d, 8.7) | 1, 3 | 4 |
| 6 | 157.2 s | |||
| 7 | 144.3 d | 7.52 (1H, d, 15.6) | 2, 4, 9 | 8 |
| 8 | 115.3 d | 6.29 (1H, d, 15.6) | 3, 9 | 7 |
| 9 | 167.9 s | |||
| 1′ | 124.8 s | |||
| 2′ | 134.2 d | 7.57 (1H, d, 2.3) | 1, 4′, 7′ | |
| 3′ | 125.5 s | |||
| 4′ | 131.1 d | 7.64 (1H, dd, 8.7, 2.3) | 2′, 6′, 7′ | 5′ |
| 5′ | 115.1 d | 6.87 (1H, d, 8.7) | 1′, 3′ | 4′ |
| 6′ | 156.0 s | |||
| 7′ | 141.5 d | 6.80 (1H, d, 12.8) | 2′, 4′, 9′ | 8′ |
| 8′ | 117.0 d | 5.74 (1H, d, 12.8) | 3′, 9′ | 7′ |
| 9′ | 167.7 s |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.; Lu, Z.; Sun, K.; Zhu, W. Effects of High Salt Stress on Secondary Metabolite Production in the Marine-Derived Fungus Spicaria elegans. Mar. Drugs 2011, 9, 535-542. https://doi.org/10.3390/md9040535
Wang Y, Lu Z, Sun K, Zhu W. Effects of High Salt Stress on Secondary Metabolite Production in the Marine-Derived Fungus Spicaria elegans. Marine Drugs. 2011; 9(4):535-542. https://doi.org/10.3390/md9040535
Chicago/Turabian StyleWang, Yi, Zhenyu Lu, Kunlai Sun, and Weiming Zhu. 2011. "Effects of High Salt Stress on Secondary Metabolite Production in the Marine-Derived Fungus Spicaria elegans" Marine Drugs 9, no. 4: 535-542. https://doi.org/10.3390/md9040535
APA StyleWang, Y., Lu, Z., Sun, K., & Zhu, W. (2011). Effects of High Salt Stress on Secondary Metabolite Production in the Marine-Derived Fungus Spicaria elegans. Marine Drugs, 9(4), 535-542. https://doi.org/10.3390/md9040535