Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting


Abstract
:1. Introduction
2. Methodology
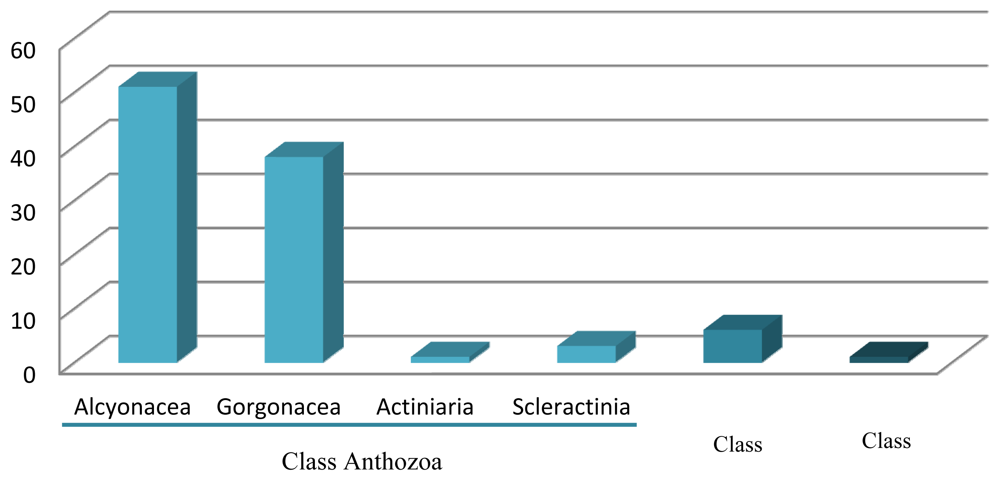
3. Class Anthozoa
3.1. Order Alcyonacea (Soft Corals)
3.2. Order Gorgonacea (Sea Fans)
3.3. Other Orders
4. Class Hydrozoa
5. Class Scyphozoa
6. Other Classes
7. Exploring the Unexplored and Being Creative: Future Perspectives for the Bioprospecting of Cnidarians
8. Conclusions
Acknowledgements
References
- Jain, R; Sonawane, S; Mandrekar, N. Marine organisms: Potential source for drug discovery. Curr. Sci 2008, 94, 292. [Google Scholar]
- Fenical, W; Jensen, PR; Palladino, MA; Lam, KS; Lloyd, GK; Potts, BC. Discovery and development of the anticancer agent salinosporamide A (NPI-0052). Bioorg. Med. Chem 2009, 17, 2175–2180. [Google Scholar]
- Hill, RT; Fenical, W. Pharmaceuticals from marine natural products: Surge or ebb? Curr. Opin. Biotechnol 2010, 21, 777–779. [Google Scholar]
- Glaser, KB; Mayer, AMS. A renaissance in marine pharmacology: From preclinical curiosity to clinical reality. Biochem. Pharmacol 2009, 78, 440–448. [Google Scholar]
- Fusetani, N. Biotechnological potential of marine natural products. Pure Appl. Chem 2010, 82, 17–26. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1999, 16, 155–198. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1998, 15, 113–158. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1997, 14, 259–302. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1996, 13, 75–125. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1995, 12, 223–269. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1994, 11, 355–395. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1993, 10, 497–539. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1991, 8, 97–147. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1990, 7, 269–309. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1988, 5, 613–663. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1987, 4, 539–576. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1986, 3, 1–33. [Google Scholar]
- Faulkner, DJ. Marine pharmacology. Antonie Van Leeuwenhoek 2000, 77, 135–145. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 2001, 18, 1–49. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 2002, 19, 1–48. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2003, 20, 1–48. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2004, 21, 1–49. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2005, 22, 15–61. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2006, 23, 26–78. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2007, 24, 31–86. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2008, 25, 35–94. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2009, 26, 170–244. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2010, 27, 165–237. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2011, 28, 196–268. [Google Scholar]
- Mayer, A; Hamann, M. Marine Pharmacology in 2000: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar. Biotechnol 2004, 6, 37–52. [Google Scholar]
- Mayer, AMS; Hamann, MT. Marine pharmacology in 2001–2002: Marine compounds with anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol 2005, 140, 265–286. [Google Scholar]
- Mayer, AMS; Gustafson, KR. Marine pharmacology in 2001–2002: Antitumour and cytotoxic compounds. Eur. J. Cancer 2004, 40, 2676–2704. [Google Scholar]
- Mayer, AMS; Rodríguez, AD; Berlinck, RGS; Hamann, MT. Marine pharmacology in 2003–2004: Marine compounds with anthelmintic antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol 2007, 145, 553–581. [Google Scholar]
- Mayer, AMS; Gustafson, KR. Marine pharmacology in 2003–2004: Anti-tumour and cytotoxic compounds. Eur. J. Cancer 2006, 42, 2241–2270. [Google Scholar]
- Mayer, AMS; Rodriguez, AD; Berlinck, RGS; Hamann, MT. Marine pharmacology in 2005–2006: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta Gen. Subj 2009, 1790, 283–308. [Google Scholar]
- Mayer, AMS; Gustafson, KR. Marine pharmacology in 2005–2006: Antitumour and cytotoxic compounds. Eur. J. Cancer 2008, 44, 2357–2387. [Google Scholar]
- Mayer, AMS; Rodríguez, AD; Berlinck, RGS; Fusetani, N. Marine pharmacology in 2007–2008: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol 2011, 153, 191–222. [Google Scholar]
- Mayer, AMS; Hamann, MT. Marine pharmacology in 1999: Compounds with antibacterial, anticoagulant, antifungal, anthelmintic, anti-inflammatory, antiplatelet, antiprotozoal and antiviral activities affecting the cardiovascular, endocrine, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol 2002, 132, 315–339. [Google Scholar]
- Sipkema, D; Franssen, MCR; Osinga, R; Tramper, J; Wijffels, RH. Marine sponges as pharmacy. Mar. Biotechnol 2005, 7, 142–162. [Google Scholar]
- Newman, DJ; Cragg, GM. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod 2004, 67, 1216–1238. [Google Scholar]
- Turk, T; Kem, WR. The phylum Cnidaria and investigations of its toxins and venoms until 1990. Toxicon 2009, 54, 1031–1037. [Google Scholar]
- Daly, M; Brugler, MR; Cartwright, P; Collins, AG; Dawson, MN; Fautin, DG; France, SC; McFadden, CS; Opresko, DM; Rodriguez, E; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar]
- Boyd, MR; Paull, KD; Rubinstein, LR. Data display and analysis strategies for the NCI disease-oriented in vitro antitumor drug screen. In Cytotoxic Anticancer Drugs: Models and Concepts for Drug Discovery and Development; Springer: Dordrecht, The Netherlands, 1992; p. 20. [Google Scholar]
- El-Gamal, AAH; Chiang, C-Y; Huang, S-H; Wang, S-K; Duh, C-Y. Xenia diterpenoids from the Formosan soft coral Xenia blumi. J. Nat. Prod 2005, 68, 1336–1340. [Google Scholar]
- Duh, CY; El-Gamal, AAH; Chu, CJ; Wang, SK; Dai, CF. New cytotoxic constituents from the Formosan soft corals Clavularia viridis and Clavularia violacea. J. Nat. Prod 2002, 65, 1535–1539. [Google Scholar]
- Appeltans, W; Bouchet, P; Boxshall, GA; Fauchald, K; Gordon, DP; Hoeksema, BW; Poore, GCB; van Soest, RWM; Stöhr, S; Walter, TC; Costello, MJ. Cnidaria. World Register of Marine Species. 2010. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=1267 accessed on 4 January 2011.
- Fautin, D. Personal communication by e-mail, University of Kansas: Lawrence, KS, USA, 2011.
- Strukelj, B; Lenarcic, B; Gruden, K; Pungercar, J; Rogelj, B; Turk, V; Bosch, D; Jongsma, MA. Equistatin, a protease inhibitor from the sea anemone Actinia equina, is composed of three structural and functional domains. Biochem. Biophys. Res. Commun 2000, 269, 732–736. [Google Scholar]
- Fontana, A; Ciavatta, ML; Cimino, G. Cladocoran A and B: Two novel γ-hydroxybutenolide sesterterpenes from the Mediterranean coral Cladocora cespitosa. J. Org. Chem 1998, 63, 2845–2849. [Google Scholar]
- Miyaoka, H; Yamanishi, M; Mitome, H. PLA2 inhibitory activity of marine sesterterpenoids cladocorans, their diastereomers and analogues. Chem. Pharm. Bull 2006, 54, 268–270. [Google Scholar]
- Meyer, M; Delberghe, F; Liron, F; Guillaume, M; Valentin, A; Guyot, M. An antiplasmodial new (bis)indole alkaloid from the hard coral Tubastraea sp. Nat. Prod. Res 2009, 23, 178–182. [Google Scholar]
- Hooper, GJ; Davies-Coleman, MT. New metabolites from the South African soft coral Capnella thyrsoidea. Tetrahedron 1995, 51, 9973–9984. [Google Scholar]
- Bhakuni, DS; Rawat, DS. Bioactive Marine Natural Products; Springer: Dordrecht, The Netherlands, 2005; p. 396. [Google Scholar]
- Wu, SL; Su, JH; Wen, ZH; Hsu, CH; Chen, BW; Dai, CF; Kuo, YH; Sheu, JH. Simplexins A–I, eunicellin-based diterpenoids from the soft coral Klyxum simplex. J. Nat. Prod 2009, 72, 994–1000. [Google Scholar]
- Chen, BW; Wu, YC; Chiang, MY; Su, JH; Wang, WH; Fan, TY; Sheu, JH. Eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Tetrahedron 2009, 65, 7016–7022. [Google Scholar]
- Nguyen, HT; Chau, VM; Phan, VK; Hoang, TH; Nguyen, HN; Nguyen, XC; Tran, HQ; Nguyen, XN; Hyun, JH; Kang, HK; et al. Chemical components from the Vietnamese soft coral Lobophytum sp. Arch. Pharm. Res 2010, 33, 503–508. [Google Scholar]
- Rashid, MA; Gustafson, KR; Boyd, MR. HIV-inhibitory cembrane derivatives from a Philippines collection of the soft coral Lobophytum species. J. Nat. Prod 2000, 63, 531–533. [Google Scholar]
- Chao, C-H; Wen, Z-H; Wu, Y-C; Yeh, H-C; Sheu, J-H. Cytotoxic and anti-inflammatory cembranoids from the soft coral Lobophytum crassum. J. Nat. Prod 2008, 71, 1819–1824. [Google Scholar]
- Coval, SJ; Patton, RW; Petrin, JM; James, L; Rothofsky, ML; Lin, SL; Patel, M; Reed, JK; McPhail, AT; Bishop, WR. A cembranolide diterpene farnesyl protein transferase inhibitor from the marine soft coral Lobophytum cristagalli. Bioorg. Med. Chem. Lett 1996, 6, 909–912. [Google Scholar]
- Cheng, S-Y; Wen, Z-H; Chiou, S-F; Hsu, C-H; Wang, S-K; Dai, C-F; Chiang, MY; Duh, C-Y. Durumolides A–E, anti-inflammatory and antibacterial cembranolides from the soft coral Lobophytum durum. Tetrahedron 2008, 64, 9698–9704. [Google Scholar]
- Cheng, SY; Wen, ZH; Wang, SK; Chiou, SF; Hsu, CH; Dai, CF; Chiang, MY; Duh, CY. Unprecedented hemiketal cembranolides with anti-inflammatory activity from the soft coral Lobophytum durum. J. Nat. Prod 2009, 72, 152–155. [Google Scholar]
- Huang, HC; Chao, CH; Kuo, YH; Sheu, JH. Crassocolides G–M, cembranoids from the Formosan soft coral Sarcophyton crassocaule. Chem. Biodivers 2009, 6, 1232–1242. [Google Scholar]
- Fusetani, N. Research toward drugs from the sea. New J. Chem 1990, 14, 721–728. [Google Scholar]
- Dmitrenok, AS; Radhika, P; Anjaneyulu, V; Subrahmanyam, C; Subba Rao, PV; Dmitrenok, PS; Boguslavsky, VM. New lipids from the soft corals of the Andaman Islands. Russ. Chem. Bull 2003, 52, 1868–1872. [Google Scholar]
- Lin, YS; Chen, CH; Liaw, CC; Chen, YC; Kuo, YH; Shen, YC. Cembrane diterpenoids from the Taiwanese soft coral Sinularia flexibilis. Tetrahedron 2009, 65, 9157–9164. [Google Scholar]
- Michalek, K; Bowden, BF. A natural algacide from soft coral Sinularia flexibilis (Coelenterata, Octocorallia, Alcyonacea). J. Chem. Ecol 1997, 23, 259–273. [Google Scholar]
- Ahmed, AF; Hsieh, Y-T; Wen, Z-H; Wu, Y-C; Sheu, J-H. Polyoxygenated sterols from the Formosan soft coral Sinularia gibberosa. J. Nat. Prod 2006, 69, 1275–1279. [Google Scholar]
- Lu, Y; Huang, CY; Lin, YF; Wen, ZH; Su, JH; Kuo, YH; Chiang, MY; Sheu, JH. Anti-inflammatory cembranoids from the soft corals Sinularia querciformis and Sinularia granosa. J. Nat. Prod 2008, 71, 1754–1759. [Google Scholar]
- Yabe, T; Yamada, H; Shimomura, M; Miyaoka, H; Yamada, Y. Induction of choline acetyltransferase activity in cholinergic neurons by stolonidiol: structure-activity relationship. J. Nat. Prod 2000, 63, 433–435. [Google Scholar]
- Iwashima, M; Matsumoto, Y; Takahashi, H; Iguchi, K. New marine cembrane-type diterpenoids from the Okinawan soft coral Clavularia koellikeri. J. Nat. Prod 2000, 63, 1647–1652. [Google Scholar]
- Lin, YS; Khalil, AT; Chiou, SH; Kuo, YC; Cheng, YB; Liaw, CC; Shen, YC. Bioactive marine prostanoids from octocoral Clavularia viridis. Chem. Biodivers 2008, 5, 784–792. [Google Scholar]
- Watanabe, K; Sekine, M; Takahashi, H; Iguchi, K. New halogenated marine prostanoids with cytotoxic activity from the Okinawan soft coral Clavularia viridis. J. Nat. Prod 2001, 64, 1421–1425. [Google Scholar]
- Shen, YC; Cheng, YB; Lin, YC; Guh, JH; Teng, CM; Ko, CL. New prostanoids with cytotoxic activity from Taiwanese octocoral Clavularia viridis. J. Nat. Prod 2004, 67, 542–546. [Google Scholar]
- Chiang, P-C; Chien, C-L; Pan, S-L; Chen, W-P; Teng, C-M; Shen, Y-C; Guh, J-H. Induction of endoplasmic reticulum stress and apoptosis by a marine prostanoid in human hepatocellular carcinoma. J. Hepatol 2005, 43, 679–686. [Google Scholar]
- Iwashima, M; Nara, K; Nakamichi, Y; Iguchi, K. Three new chlorinated marine steroids, yonarasterols G, H and I, isolated from the Okinawan soft coral Clavularia viridis. Steroids 2001, 66, 25–32. [Google Scholar]
- Verbitski, SM; Mullally, JE; Fitzpatrick, FA; Ireland, CM. Punaglandins, chlorinated prostaglandins, function as potent Michael receptors to inhibit ubiquitin isopeptidase activity. J. Med. Chem 2004, 47, 2062–2070. [Google Scholar]
- Tomono, Y; Hirota, H; Fusetani, N. Isogosterones A–D, antifouling 13,17-Secosteroids from an octocoral Dendronephthya sp. J. Org. Chem 1999, 64, 2272–2275. [Google Scholar]
- Grote, D; Hanel, F; Dahse, HM; Seifert, K. Capnellenes from the soft coral Dendronephthya rubeola. Chem. Biodivers 2008, 5, 1683–1693. [Google Scholar]
- Peukert, K; Staller, P; Schneider, A; Carmichael, G; Hanel, F; Eilers, M. An alternative pathway for gene regulation by Myc. EMBO J 1997, 16, 5672–5686. [Google Scholar]
- Hermeking, H. The MYC oncogene as a cancer drug target. Curr. Cancer Drug Targets 2003, 3, 163–175. [Google Scholar]
- Cheng, SY; Huang, KJ; Wang, SK; Wen, ZH; Hsu, CH; Dai, CF; Duh, CY. New terpenoids from the soft corals Sinularia capillosa and Nephthea chabroli. Org. Lett 2009, 11, 4830–4833. [Google Scholar]
- Cheng, SY; Wen, ZH; Wang, SK; Chiang, MY; El-Gamal, AAH; Dai, CF; Duh, CY. Revision of the absolute configuration at C(23) of Lanostanoids and isolation of secondary metabolites from Formosan soft coral Nephthea erecta. Chem. Biodivers 2009, 6, 86–95. [Google Scholar]
- Lin, YC; Abd El-Razek, MH; Hwang, TL; Chiang, MY; Kuo, YH; Dai, CF; Shen, YC. Asterolaurins A–F, xenicane diterpenoids from the Taiwanese soft coral Asterospicularia laurae. J. Nat. Prod 2009, 72, 1911–1916. [Google Scholar]
- Duh, C-Y; El-Gamal, AAH; Wang, S-K; Dai, C-F. Novel terpenoids from the Formosan soft coral Cespitularia hypotentaculata. J. Nat. Prod 2002, 65, 1429–1433. [Google Scholar]
- Bishara, A; Rudi, A; Goldberg, I; Benayahu, Y; Kashman, Y. Novaxenicins A–D and xeniolides I–K, seven new diterpenes from the soft coral Xenia novaebrittanniae. Tetrahedron 2006, 62, 12092–12097. [Google Scholar]
- Konig, GM; Wright, AD. New cembranoid diterpenes from the soft coral Sarcophyton ehrenbergi. J. Nat. Prod 1998, 61, 494–496. [Google Scholar]
- Chang, CH; Wen, ZH; Wang, SK; Duh, CY. Capnellenes from the Formosan soft coral Capnella imbricata. J. Nat. Prod 2008, 71, 619–621. [Google Scholar]
- Duh, CY; Hou, RS. Cytotoxic cembranoids from the soft corals Sinularia gibberosa and Sarcophyton trocheliophorum. J. Nat. Prod 1996, 59, 595–598. [Google Scholar]
- Su, JH; Ahmed, AF; Sung, PJ; Chao, CH; Kuo, YH; Sheu, JH. Manaarenolides A–I, diterpenoids from the soft coral Sinularia manaarensis. J. Nat. Prod 2006, 69, 1134–1139. [Google Scholar]
- Norton, RS; Kazlauskas, R. C-13 NMR-study of flexibilide, an anti-inflammatory agent from a soft coral. Experientia 1980, 36, 276–278. [Google Scholar]
- Williams, DH; Faulkner, DJ. Two practical syntheses of an anti-inflammatory sesquiterpene furoic acid from Sinularia spp. Tetrahedron 1996, 52, 4245–4256. [Google Scholar]
- Weinheimer, AJ; Matson, JA; Hossain, MB; van der Helm, D. Marine anticancer agents: sinularin and dihydrosinularin, new cembranolides from the soft coral Sinularia flexibilis. Tetrahedron Lett 1977, 18, 2923–2926. [Google Scholar]
- Li, GQ; Zhang, YL; Deng, ZW; van Ofwegen, LP; Proksch, P; Lin, WH. Cytotoxic cembranoid Diterpenes from a soft coral Sinularia gibberosa. J. Nat. Prod 2005, 68, 649–652. [Google Scholar]
- Aceret, TL; Coll, JC; Uchio, Y; Sammarco, PW. Antimicrobial activity of the diterpenes flexibilide and sinulariolide derived from Sinularia flexibilis Quoy and Gaimard 1833 (Coelenterata: Alcyonacea, Octocorallia). Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol 1998, 120, 121–126. [Google Scholar]
- Aceret, TL; Brown, L; Miller, J; Coll, JC; Sammarco, PW. Cardiac and vascular responses of isolated rat tissues treated with diterpenes from Sinularia flexibilis (Coelenterata: Octocorallia). Toxicon 1996, 34, 1165–1171. [Google Scholar]
- Sheikh, YM; Singy, G; Kaisin, M; Eggert, H; Djerassi, C; Tursch, B; Daloze, D; Braekman, JC. Terpenoids—LXXI: Chemical studies of marine invertebrates-XIV. Four representatives of a novel sesquiterpene class—the capnellane skeleton. Tetrahedron 1976, 32, 1171–1178. [Google Scholar]
- Kaisin, M; Sheikh, YM; Durham, LJ; Djerassi, C; Tursch, B; Daloze, D; Braekman, JC; Losman, D; Karlsson, R. Capnellane—a new tricyclic sesquiterpene skeleton from the soft coral Capnella imbricata. Tetrahedron Lett 1974, 15, 2239–2242. [Google Scholar]
- Kaisin, M; Braekman, JC; Daloze, D; Tursch, B. Novel acetoxycapnellenes from the alcyonacean Capnella imbricata. Tetrahedron 1985, 41, 1067–1072. [Google Scholar]
- Nakao, Y; Fusetani, N. Enzyme inhibitors from marine invertebrates. J. Nat. Prod 2007, 70, 689–710. [Google Scholar]
- Sata, NU; Sugano, M; Matsunaga, S; Fusetani, N. Sinulamide: An H,K-ATPase inhibitor from a soft coral Sinularia sp. Tetrahedron Lett 1999, 40, 719–722. [Google Scholar]
- Huang, H-C; Wen, Z-H; Chao, C-H; Ahmed, AF; Chiang, MY; Kuo, Y-H; Hsu, C-H; Sheu, J-H. Novel sesquiterpenoids from the Formosan soft coral Paralemnalia thyrsoides. Tetrahedron Lett 2006, 47, 8751–8755. [Google Scholar]
- Coll, JC. The chemistry and chemical ecology of octocorals (Coelenterata, Anthozoa, Octocorallia). Chem. Rev 1992, 92, 613–631. [Google Scholar]
- Maida, M; Sammarco, PW; Coll, JC. A diffusion chamber for assessing efficacy of natural anti-fouling defenses in marine organisms. J. Exp. Mar. Biol. Ecol 2006, 337, 59–64. [Google Scholar]
- Chambers, LD; Stokes, KR; Walsh, FC; Wood, RJK. Modern approaches to marine antifouling coatings. Surf. Coat. Technol 2006, 201, 3642–3652. [Google Scholar]
- Watanabe, K; Iwashima, M; Iguchi, K. New marine prostanoid carboxylate salts from the Okinawan soft coral Clavularia viridis. J. Nat. Prod 1996, 59, 980–982. [Google Scholar]
- Iwashima, M; Terada, I; Okamoto, K; Iguchi, K. Tricycloclavulone and clavubicyclone, novel prostanoid-related marine oxylipins, isolated from the Okinawan soft coral Clavularia viridis. J. Org. Chem 2002, 67, 2977–2981. [Google Scholar]
- Kusumi, T; Hamada, T; Hara, M; Ishitsuka, MO; Ginda, H; Kakisawa, H. Structure and absolute-configuration of Isoclavukerin-A, a component from an Okinawan soft coral. Tetrahedron Lett 1992, 33, 2019–2022. [Google Scholar]
- Davies, P; Maloney, AJF. Selective loss of central cholinergic neurons in Alzheimer’s disease. Lancet 1976, 308, 1403–1403. [Google Scholar]
- Whitehouse, P; Price, D; Struble, R; Clark, A; Coyle, J; Delon, M. Alzheimer’s disease and senile dementia: Loss of neurons in the basal forebrain. Science 1982, 215, 1237–1239. [Google Scholar]
- Bartus, R; Dean, R; Beer, B; Lippa, A. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar]
- Chiang, PC; Kung, FL; Huang, DM; Li, TK; Fan, JR; Pan, SL; Shen, YC; Guh, JH. Induction of Fas clustering and apoptosis by coral prostanoid in human hormone-resistant prostate cancer cells. Eur. J. Pharmacol 2006, 542, 22–30. [Google Scholar]
- Tsukamoto, S; Yokosawa, H. Inhibition of the ubiquitin-proteasome system by natural products for cancer therapy. Planta Med 2010, 76, 1064–1074. [Google Scholar]
- Chai, XY; Sun, JF; Tang, LY; Yang, XW; Li, YQ; Huang, H; Zhou, XF; Yang, B; Liu, YH. A novel cyclopentene derivative and a polyhydroxylated steroid from a South China sea gorgonian Menella sp. Chem. Pharm. Bull 2010, 58, 1391–1394. [Google Scholar]
- Sheu, J-H; Hung, K-C; Wang, G-H; Duh, C-Y. New Cytotoxic Sesquiterpenes from the Gorgonian Isis hippuris. J. Nat. Prod 2000, 63, 1603–1607. [Google Scholar]
- González, N; Barral, MA; Rodríguez, J; Jiménez, C. New cytotoxic steroids from the gorgonian Isis hippuris. Structure-activity studies. Tetrahedron 2001, 57, 3487–3497. [Google Scholar]
- Sheu, JH; Chao, CH; Wang, GH; Hung, KC; Duh, CY; Chiang, MY; Wu, YC; Wu, CC. The first A-nor-hippuristanol and two novel 4,5-secosuberosanoids from the Gorgonian Isis hippuris. Tetrahedron Lett 2004, 45, 6413–6416. [Google Scholar]
- Chao, C-H; Huang, L-F; Yang, Y-L; Su, J-H; Wang, G-H; Chiang, MY; Wu, Y-C; Dai, C-F; Sheu, J-H. Polyoxygenated steroids from the gorgonian Isis hippuris. J. Nat. Prod 2005, 68, 880–885. [Google Scholar]
- Sung, P-J; Chen, Y-P; Hwang, T-L; Hu, W-P; Fang, L-S; Wu, Y-C; Li, J-J; Sheu, J-H. Briaexcavatins C–F, four new briarane-related diterpenoids from the Formosan octocoral Briareum excavatum (Briareidae). Tetrahedron 2006, 62, 5686–5691. [Google Scholar]
- Sung, P-J; Su, J-H; Duh, C-Y; Chiang, MY; Sheu, J-H. Briaexcavatolides K–N, new briarane diterpenes from the Gorgonian Briareum excavatum. J. Nat. Prod 2001, 64, 318–323. [Google Scholar]
- Ospina, CA; Rodríguez, AD; Ortega-Barria, E; Capson, TL. Briarellins J–P and Polyanthellin A: New Eunicellin-based diterpenes from the gorgonian coral Briareum polyanthes and their antimalarial activity. J. Nat. Prod 2003, 66, 357–363. [Google Scholar]
- Shen, YC; Chen, YH; Hwang, TL; Guh, JH; Khalil, AT. Four new briarane diterpenoids from the gorgonian coral Junceella fragilis. Helv. Chim. Acta 2007, 90, 1391–1398. [Google Scholar]
- Qi, SH; Zhang, S; Qian, PY; Xu, HH. Antifeedant and antifouling briaranes from the South China Sea gorgonian Junceella juncea. Chem. Nat. Compd 2009, 45, 49–54. [Google Scholar]
- Targett, NM; Bishop, SS; McConnell, OJ; Yoder, JA. Antifouling agents against the benthic marine diatom Navicula salinicola: Homarine from the gorgonians Leptogorgia virgulata and L. setacea and analogs. J. Chem. Ecol 1983, 9, 817–829. [Google Scholar]
- Gerhart, DJ; Rittschof, D; Mayo, SW. Chemical ecology and the search for marine antifoulants—Studies of a predatory-prey symbiosis. J. Chem. Ecol 1988, 14, 1905–1917. [Google Scholar]
- He, HY; Kulanthaivel, P; Baker, BJ; Kalter, K; Darges, J; Cofield, D; Wolff, L; Adams, L. New antiproliferative and antiinflammatory 9,11-secosterols from the gorgonian Pseudopterogorgia sp. Tetrahedron 1995, 51, 51–58. [Google Scholar]
- Kate, AS; Pearson, JK; Ramanathan, B; Richard, K; Kerr, RG. Isolation, biomimetic synthesis, and cytotoxic activity of bis(pseudopterane) amines. J. Nat. Prod 2009, 72, 1331–1334. [Google Scholar]
- Ospina, CA; Rodríguez, AD; Zhao, H; Raptis, RG. Bipinnapterolide B, a bioactive oxapolycyclic diterpene from the Colombian gorgonian coral Pseudopterogorgia bipinnata. Tetrahedron Lett 2007, 48, 7520–7523. [Google Scholar]
- Ospina, CA; Rodríguez, AD; Sánchez, JA; Ortega-Barria, E; Capson, TL; Mayer, AMS. Caucanolides A–F, unusual antiplasmodial constituents from a Colombian collection of the gorgonian coral Pseudopterogorgia bipinnata. J. Nat. Prod 2005, 68, 1519–1526. [Google Scholar]
- Ata, A; Win, HY; Holt, D; Holloway, P; Segstro, EP; Jayatilake, GS. New antibacterial diterpenes from Pseudopterogorgia elisabethae. Helv. Chim. Acta 2004, 87, 1090–1098. [Google Scholar]
- Rodríguez, II; Rodríguez, AD; Wang, Y; Franzblau, SG. Ileabethoxazole: A novel benzoxazole alkaloid with antimycobacterial activity. Tetrahedron Lett 2006, 47, 3229–3232. [Google Scholar]
- Rodríguez, II; Rodríguez, AD. Homopseudopteroxazole, a new antimycobacterial diterpene alkaloid from Pseudopterogorgia elisabethae. J. Nat. Prod 2003, 66, 855–857. [Google Scholar]
- Wei, X; Rodríguez, II; Rodríguez, AD; Barnes, CL. Caribenols A and B, sea whip derived norditerpenes with novel tricarbocyclic skeletons. J. Org. Chem 2007, 72, 7386–7389. [Google Scholar]
- Rodriguez, AD; Ramirez, C; Rodriguez, II; Barnes, CL. Novel terpenoids from the West Indian sea whip Pseudopterogorgia elisabethae (Bayer). Elisapterosins A and B: Rearranged diterpenes possessing an unprecedented cagelike framework. J. Org. Chem 2000, 65, 1390–1398. [Google Scholar]
- Rodriguez, II; Rodriguez, AD; Zhao, H. Aberrarone: A gorgonian-derived diterpene from Pseudopterogorgia elisabethae. J. Org. Chem 2009, 74, 7581–7584. [Google Scholar]
- Marrero, J; Rodríguez, AD; Baran, P; Raptis, RG; Sánchez, JA; Ortega-Barria, E; Capson, TL. Bielschowskysin, a gorgonian-derived biologically active diterpene with an unprecedented carbon skeleton. Org. Lett 2004, 6, 1661–1664. [Google Scholar]
- McEnroe, FJ; Fenical, W. Structures and synthesis of some new antibacterial sesquiterpenoids from the gorgonian coral Pseudopterogorgia rigida. Tetrahedron 1978, 34, 1661–1664. [Google Scholar]
- Garzón, SP; Rodríguez, AD; Sánchez, JA; Ortega-Barria, E. Sesquiterpenoid metabolites with antiplasmodial activity from a Caribbean gorgonian coral Eunicea sp. J. Nat. Prod 2005, 68, 1354–1359. [Google Scholar]
- Shin, JH; Fenical, W. Fuscosides A–D: Antinflammatory diterpenoid glycosides of new structural classes from the Caribbean gorgonian Eunicea fusca. J. Org. Chem 1991, 56, 3153–3158. [Google Scholar]
- Kikuchi, H; Tsukitani, Y; Nakanishi, H; Shimizu, I; Saitoh, S; Iguchi, K; Yamada, Y. New butenolides from the gorgonian Euplexaura flava (Nutting). Chem. Lett 1982, 11, 233–236. [Google Scholar]
- Rodríguez, AD. The natural products chemistry of West Indian gorgonian octocorals. Tetrahedron 1995, 51, 4571–4618. [Google Scholar]
- Fenical, W. Marine soft corals of the genus Pseudopterogorgia: A resource for novel anti-inflammatory diterpenoids. J. Nat. Prod 1987, 50, 1001–1008. [Google Scholar]
- Marrero, J; Ospina, CA; Rodríguez, AD; Baran, P; Zhao, H; Franzblau, SG; Ortega-Barria, E. New diterpenes of the pseudopterane class from two closely related Pseudopterogorgia species: Isolation, structural elucidation, and biological evaluation. Tetrahedron 2006, 62, 6998–7008. [Google Scholar]
- Martelletti, P; Adriani, E; Bonini, S; Celestino, D; Lenti, L; Armaleo, C; Dipastena, A; Misasi, R; Giacovazzo, M. Basophil histamine-release and Leukotriene (LTB4-LTC4) production in cluster headache. Headache 1989, 29, 46–48. [Google Scholar]
- Jacobson, PB; Jacobs, RS. Fuscoside—An antiinflammatory marine natural product which selectively inhibits 5-lipoxygenase. Biochemical-studies in the human neutrophil. J. Pharmacol. Exp. Ther 1992, 262, 874–882. [Google Scholar]
- Nathan, C. Neutrophils and immunity: challenges and opportunities. Nat. Rev. Immunol 2006, 6, 173–182. [Google Scholar]
- Lacy, P; Eitzen, G. Control of granule exocytosis in neutrophils. Front. Biosci 2008, 13, 5559–5570. [Google Scholar]
- Pham, CTN. Neutrophil serine proteases: specific regulators of inflammation. Nat. Rev. Immunol 2006, 6, 541–550. [Google Scholar]
- Boukouvalas, J; Loach, RP. General, regiodefined access to alpha-substituted butenolides through metal-halogen exchange of 3-bromo-2-silyloxyfurans. Efficient synthesis of an anti-inflammatory gorgonian lipid. J. Org. Chem 2008, 73, 8109–8112. [Google Scholar]
- Sung, PJ; Sheu, JH; Xu, JP. Survey of briarane-type diterpenoids of marine origin. Heterocycles 2002, 57, 535–579. [Google Scholar]
- Sung, PJ; Chang, PC; Fang, LS; Sheu, JH; Chen, WC; Chen, YP; Lin, MR. Survey of briarane-related diterpenoids—part II. Heterocycles 2005, 65, 195–204. [Google Scholar]
- Wu, SL; Sung, PJ; Chiang, MY; Wu, JY; Sheu, JH. New polyoxygenated briarane diterpenoids, Briaexcavatolides O–R, from the Gorgonian Briareum excavatum. J. Nat. Prod 2001, 64, 1415–1420. [Google Scholar]
- Lenarcic, B; Ritonja, A; Strukelj, B; Turk, B; Turk, V. Equistatin, a new inhibitor of cysteine proteinases from Actinia equina, is structurally related to thyroglobulin type-1 domain. J. Biol. Chem 1997, 272, 13899–13903. [Google Scholar]
- Lenarcic, B; Turk, V. Thyroglobulin Type-1 domains in equistatin inhibit both Papain-like cysteine proteinases and Cathepsin D. J. Biol. Chem 1999, 274, 563–566. [Google Scholar]
- Brömme, D; Petanceska, S. Papain-Like Cysteine Proteases and Their Implications in Neurodegenerative Diseases. In Role of Proteases in the Pathophysiology of Neurodegenerative Diseases; Lajtha, A, Banik, NL, Eds.; Springer: New York, NY, USA, 2002; pp. 47–61. [Google Scholar]
- Wolf, M; Clark-Lewis, I; Buri, C; Langen, H; Lis, M; Mazzucchelli, L. Cathepsin D specifically cleaves the chemokines macrophage inflammatory protein-la, macrophage inflammatory protein-1 beta, and SLC that are expressed in human breast cancer. Am. J. Pathol 2003, 162, 1183–1190. [Google Scholar]
- NCBI. CTSD cathepsin D [Homo sapiens]. Entrez Gene—Genes and mapped phenotypes. 2010. Available online: http://www.ncbi.nlm.nih.gov/gene/1509 accessed on 7 December 2010.
- Muller, AJ; DuHadaway, JB; Donover, PS; Sutanto-Ward, E; Prendergast, GC. Inhibition of indoleamine 2,3-dioxygenase, an immunoregulatory target of the cancer suppression gene Bin1, potentiates cancer chemotherapy. Nat. Med 2005, 11, 312–319. [Google Scholar]
- Grohmann, U; Fallarino, F; Puccetti, P. Tolerance, DCs and tryptophan: much ado about IDO. Trends Immunol 2003, 24, 242–248. [Google Scholar]
- Pereira, A; Vottero, E; Roberge, M; Mauk, AG; Andersen, RJ. Indoleamine 2,3-dioxygenase inhibitors from the northeastern Pacific marine hydroid Garveia annulata. J. Nat. Prod 2006, 69, 1496–1499. [Google Scholar]
- Muller, AJ; Malachowski, WP; Prendergast, GC. Indoleamine 2,3-dioxygenase in cancer: Targeting pathological immune tolerance with small-molecule inhibitors. Expert Opin. Ther. Targets 2005, 9, 831–849. [Google Scholar]
- Seo, YW; Cho, KW; Rho, JR; Shin, JH; Kwon, BM; Bok, SH; Song, JI. Solandelactones A–I, lactonized cyclopropyl oxylipins isolated from the hydroid Solanderia secunda. Tetrahedron 1996, 52, 10583–10596. [Google Scholar]
- Ovchinnikova, TV; Balandin, SV; Aleshina, GM; Tagaev, AA; Leonova, YF; Krasnodembsky, ED; Men’shenin, AV; Kokryakov, VN. Aurelin, a novel antimicrobial peptide from jellyfish Aurelia aurita with structural features of defensins and channel-blocking toxins. Biochem. Biophys. Res. Commun 2006, 348, 514–523. [Google Scholar]
- Brinkman, DL; Burnell, JN. Biochemical and molecular characterisation of cubozoan protein toxins. Toxicon 2009, 54, 1162–1173. [Google Scholar]
- Synnes, M. Bioprospecting of organisms from the deep sea: Scientific and environmental aspects. Clean Technol. Environ. Policy 2007, 9, 53–59. [Google Scholar]
- Moura, CJ; Cunha, MR; Schuchert, P. Tubiclavoides striatum gen. nov et sp nov (Cnidaria: Hydrozoa) a new bathyal hydroid from the Gulf of Cadiz, north-east Atlantic Ocean. J. Mar. Biol. Assoc. UK 2007, 87, 421–428. [Google Scholar]
- Rodriguez, E; Lopez-Gonzalez, PJ; Daly, M. New family of sea anemones (Actiniaria, Acontiaria) from deep polar seas. Polar Biol 2009, 32, 703–717. [Google Scholar]
- Census of Marine Life. Available online: http://www.coml.org/discoveries/species/bathyal_hydroid accessed on 3 January 2011.
- Fautin, DG. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar]
- Clark, MR; Tittensor, D; Rogers, AD; Brewin, P; Schlacher, T; Rowden, A; Stocks, K; Consalvey, M. Seamounts, Deep-sea Corals and Fisheries: Vulnerability of Deep-Sea Corals to Fishing on Seamounts Beyond Areas of National Jurisdiction; UNEP-WCMC: Cambridge, UK, 2006. [Google Scholar]
- Rodriguez, E; Daly, M. Phylogenetic relationships among deep-sea and chemosynthetic sea anemones: Actinoscyphiidae and actinostolidae (Actiniaria: Mesomyaria). PLoS One 2010, 5, e10958. [Google Scholar]
- Skropeta, D. Deep-sea natural products. Nat. Prod. Rep 2008, 25, 1131–1166. [Google Scholar]
- Kingston, DGI. Modern natural products drug discovery and its relevance to biodiversity conservation. J. Nat. Prod 2010, 74, 496–511. [Google Scholar]
- Wabnitz, C; Taylor, M; Green, E; Razak, T. From Ocean to Aquarium; UNEP-WCMC: Cambridge, UK, 2003. [Google Scholar]
- White, RE. High-throughput screening in drug metabolism and pharmacokinetic support of drug discovery. Ann. Rev. Pharmacol. Toxicol 2000, 40, 133–157. [Google Scholar]
- Koehn, FE; Carter, GT. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov 2005, 4, 206–220. [Google Scholar]
- Li, JWH; Vederas, JC. Drug discovery and natural products: End of an Era or an endless frontier? Science 2009, 325, 161–165. [Google Scholar]
- Shafir, S; Van Rijn, J; Rinkevich, B. Coral nubbins as source material for coral biological research: A prospectus. Aquaculture 2006, 259, 444–448. [Google Scholar]
- Sella, I; Benayahu, Y. Rearing cuttings of the soft coral Sarcophyton glaucum (Octocorallia, Alcyonacea): Towards mass production in a closed seawater system. Aquaculture Res 2010, 41, 1748–1758. [Google Scholar]
- Qian, PY; Xu, Y; Fusetani, N. Natural products as antifouling compounds: recent progress and future perspectives. Biofouling 2010, 26, 223–234. [Google Scholar]
- Mendola, D. Aquaculture of three phyla of marine invertebrates to yield bioactive metabolites: process developments and economics. Biomol. Eng 2003, 20, 441–458. [Google Scholar]
- Paterson, I; Anderson, EA. The renaissance of natural products as drug candidates. Science 2005, 310, 451–453. [Google Scholar]
- Khosla, C; Keasling, JD. Timeline—Metabolic engineering for drug discovery and development. Nat. Rev. Drug Discov 2003, 2, 1019–1025. [Google Scholar]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep 2009, 26, 338–362. [Google Scholar]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep 2004, 21, 519–538. [Google Scholar]
- Shimomura, O; Johnson, FH; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol 1962, 59, 223–239. [Google Scholar]
- Schnitzler, C; Keenan, R; McCord, R; Matysik, A; Christianson, L; Haddock, S. Spectral diversity of fluorescent proteins from the Anthozoan Corynactis californica. Mar. Biotechnol 2008, 10, 328–342. [Google Scholar]
- Ip, DTM; Wong, KB; Wan, DCC. Characterization of novel orange fluorescent protein cloned from cnidarian tube anemone Cerianthus sp. Mar. Biotechnol 2007, 9, 469–478. [Google Scholar]
- Chan, MCY; Karasawa, S; Mizuno, H; Bosanac, I; Ho, D; Prive, GG; Miyawaki, A; Ikura, M. Structural characterization of a blue chromoprotein and its yellow mutant from the sea anemone Cnidopus japonicus. J. Biol. Chem 2006, 281, 37813–37819. [Google Scholar]
- Ai, HW; Henderson, JN; Remington, SJ; Campbell, RE. Directed evolution of a monomeric, bright and photostable version of Clavularia cyan fluorescent protein: Structural characterization and applications in fluorescence imaging. Biochem. J 2006, 400, 531–540. [Google Scholar]
- Goulding, A; Shrestha, S; Dria, K; Hunt, E; Deo, SK. Red fluorescent protein variants with incorporated non-natural amino acid analogues. Protein Eng. Des. Sel 2008, 21, 101–106. [Google Scholar]
- Tu, HB; Xiong, Q; Zhen, SL; Zhong, XF; Peng, LH; Chen, HP; Jiang, XY; Liu, WH; Yang, WL; Wei, JW; Dong, ML; Wu, WY; Xu, AL. A naturally enhanced green fluorescent protein from magnificent sea anemone (Heteractis magnifica) and its functional analysis. Biochem. Biophys. Res. Commun 2003, 301, 879–885. [Google Scholar]
- Pomponi, SA. The oceans and Human health: The discovery and development of marine-derived drugs. Oceanography 2001, 14, 78–87. [Google Scholar]
- Blunt, JW; Munro, MHG. Dictionary of Marine Natural Products with CD-ROM; Chapman & Hall/CRC, Taylor & Francis Group: Boca Raton, FL, USA, 2008; p. 119. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Order | |
|---|---|---|---|
| Cnidaria (≈11,287 species) | Anthozoa (≈7500 species) | Actiniaria Antipatharia Ceriantharia Corallimorpharia Scleractinia | Zoanthidea Alcyonacea Gorgonacea Helioporacea Pennatulacea |
| Cubozoa (≈36 species) | Carybdeida | Chirodropida | |
| Hydrozoa (≈3500 species) | Anthoathecata Leptothecata Siphonophorae Actinulida | Limnomedusae Narcomedusae Trachymedusae | |
| Polypodiozoa (1 species) | |||
| Scyphozoa (≈200 species) | Coronatae Rhizostomeae | Semaeostomeae | |
| Staurozoa (≈50 species) | Stauromedusae | ||
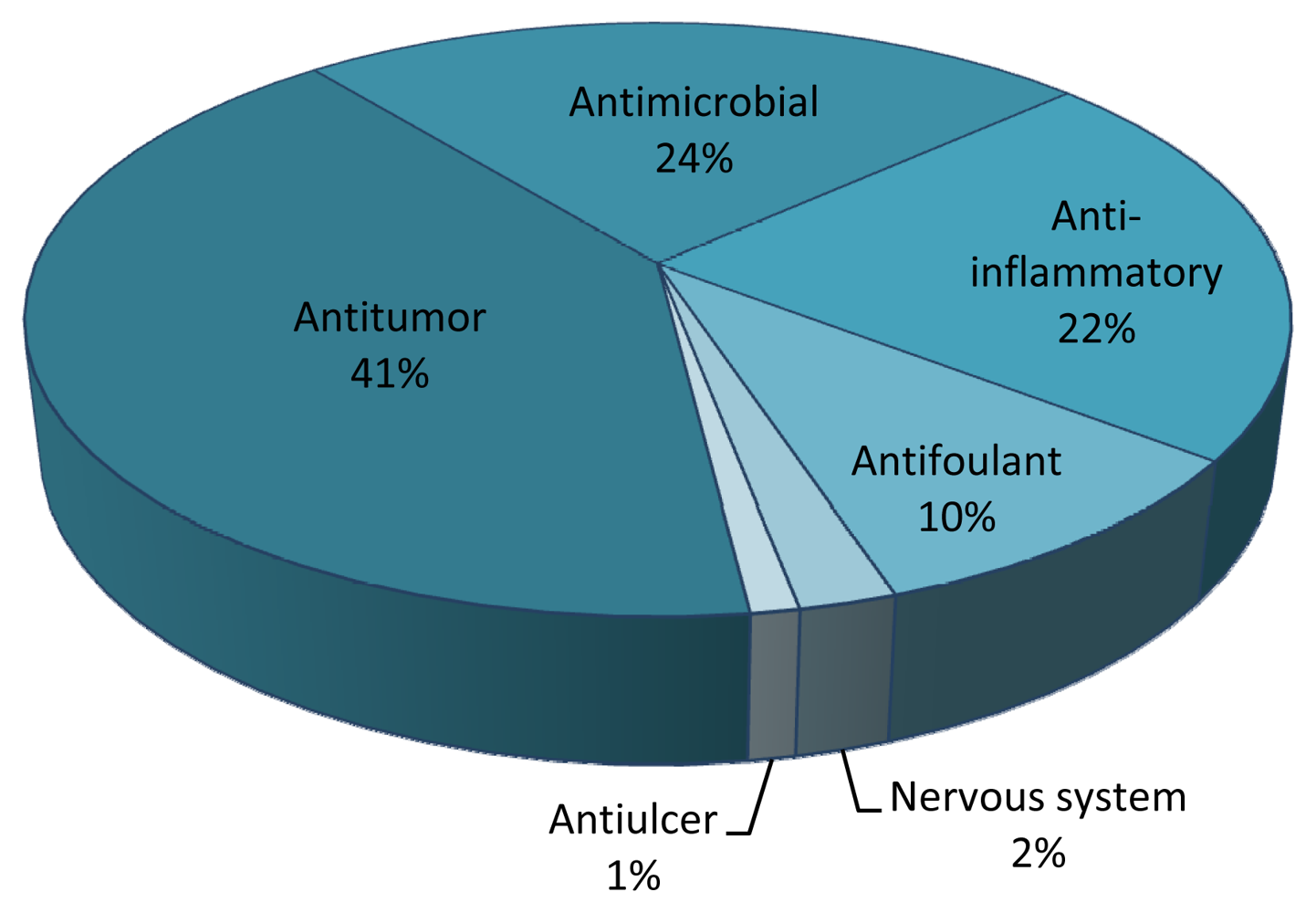
| Family and Species | Drug Class | Compound | Chemistry | Country | Ref. |
|---|---|---|---|---|---|
| Alcyoniidae | |||||
| Klyxum simplex | Anti-inflammatory | Simplexin E | Diterpenoid | TAIW | [54] |
| Klyxum simplex | Antitumor | Klysimplexin B and H | Diterpenoid | TAIW | [55] |
| Lobophytum sp. | Antitumor | Lobophytene | Diterpenoid | VN | [56] |
| Lobophytum sp. | Anti-HIV | Lobohedleolide | Diterpenoid | PHL | [57] |
| Lobophytum sp. | Anti-HIV | (7Z)-lobohedleolide, | Diterpenoid | PHL | [57] |
| Lobophytum sp. | Anti-HIV | 17-dimethylamino lobohedleolide | Diterpenoid | PHL | [57] |
| Lobophytum crassum | Anti-inflammatory | Crassumolides A and C | Terpenoid | TAIW | [58] |
| Lobophytum cristagalli | Antitumor | Cembranolide diterpene | Diterpenoid | RSC | [59] |
| Lobophytum durum | Anti-inflammatory | Durumolides A–C | Terpenoid | TAIW | [60] |
| Lobophytum durum | Anti-inflammatory | Durumhemiketalolide A–C | Cembranoid | TAIW | [61] |
| Sarcophyton crassocaule | Antitumor | Crassocolides H–M | Cembranoid | TAIW | [62] |
| Sinularia sp. | Antiulcer | Sinulide | Spermine | [63] | |
| Sinularia sp. | Antimicrobial | Lipids | Polyketide | RUS | [64] |
| Sinularia flexibilis | Antitumor | Flexilarin D | Cembranoid | TAIW | [65] |
| Sinularia flexibilis | Antifoulant | 11-episinulariolide | Diterpenoid | AUS | [66] |
| Sinularia gibberosa | Anti-inflammatory | Gibberoketosterol | Steroid | TAIW | [67] |
| Sinularia querciformis | Anti-inflammatory | Querciformolide C | Terpenoid | TAIW | [68] |
| Clavulariidae | |||||
| Clavularia sp. | Nervous system | Stolonidiol | Diterpenoid | JPN | [69] |
| Clavularia koellikeri | Antitumor | Cembrane-type diterpenoid | Diterpenoid | JPN | [70] |
| Clavularia viridis | Antitumor | Claviridic acid | Prostanoid | TAIW | [71] |
| Clavularia viridis | Antitumor | Clavulones | Prostanoid | TAIW | [71] |
| Clavularia viridis | Antitumor | Claviridenone | Prostanoid | TAIW | [45] |
| Clavularia viridis | Antitumor | Halogenated prostanoids | Prostanoid | JPN | [72] |
| Clavularia viridis | Antitumor | Bromovulone III | Prostanoid | TAIW | [73,74] |
| Clavularia viridis | Antitumor | Yonarasterols | Steroid | JPN | [75] |
| Clavularia viridis | Antitumor | Stoloniferone E | Steroid | TAIW | [45] |
| Telesto riisei | Antitumor | Punaglandins | Prostaglandin | USA | [76] |
| Nephtheidae | |||||
| Dendronephthya sp. | Antifoulant | Isogosterones A–D | Steroid | JPN | [77] |
| Dendronephthya rubeola | Antitumour | Capnell-9(12)-ene-8β,10α-diol | Sesquiterpenoid | DE | [78,79,80] |
| Nephthea chabroli | Antitumor | Chabranol | Terpenoid | TAIW | [81] |
| Nephthea erecta | Anti-inflammatory | Ergostanoids 1 and 3 | Ergostanoid | TAIW | [82] |
| Xeniidae | |||||
| Asterospicularia laurae | Antitumor | Asterolaurin A | Diterpenoid | TAIW | [83] |
| Cespitularia hypotentaculata | Antitumor | Cespitularin C | Diterpenoid | TAIW | [84] |
| Xenia novaebritanniae | Antibacterial | Xeniolide I | Diterpenoid | ISR | [85] |
| Xenia plicata | Antitumor | Blumiolide C | Diterpenoid | TAIW | [44] |
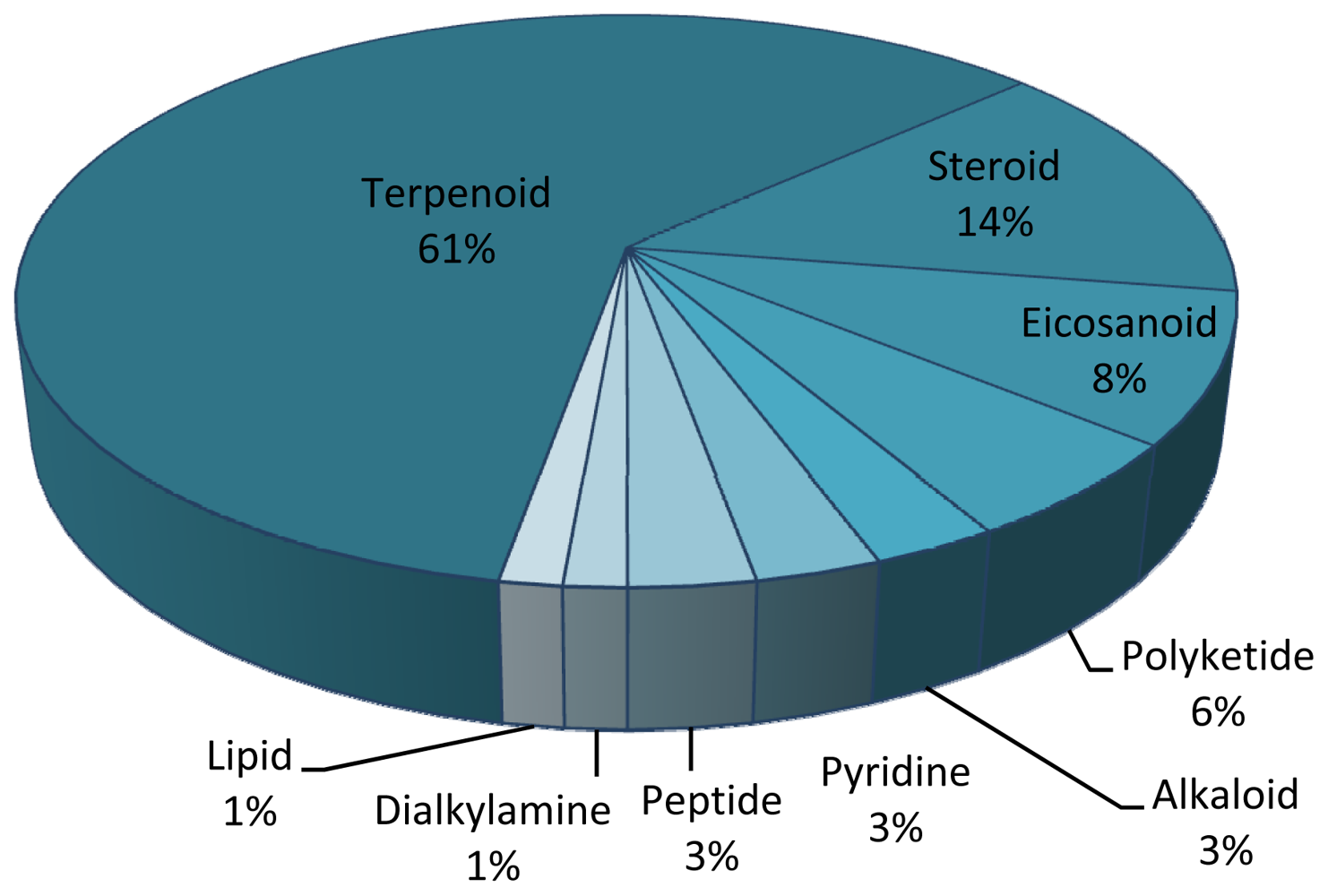
| Family and Species | Drug Class | Compound | Chemistry | Country | Ref. |
|---|---|---|---|---|---|
| Briareidae | |||||
| Briareum excavate | Anti-inflammatory | Briaexcavatin E | Diterpenoid | TAIW | [118] |
| Briareum excavate | Antitumor | Briaexcavatolides L and P | Diterpenoid | TAIW | [119] |
| Briareum asbestinum | Antimalarial | Briarellin D, K and L | Diterpenoid | PAN, USA | [120] |
| Ellisellidae | |||||
| Junceella fragilis | Anti-inflammatory | Frajunolides B and C | Terpenoid | TAIW | [121] |
| Junceella juncea | Antifoulant | Juncin ZII | Diterpenoid | TAIW | [122] |
| Gorgoniidae | |||||
| Leptogorgia setácea | Antifoulant | Homarine | Pyridine | GEO | [123] |
| Leptogorgia virgulata | Antifoulant | Homarine | Pyridine | GEO | [123] |
| Leptogorgia virgulata | Antifoulant | Pukalide | Diterpenoid | USA | [124] |
| Leptogorgia virgulata | Antifoulant | Epoxypukalide | Diterpenoid | USA | [124] |
| Pseudopterogorgia sp. | Antitumor | Secosterols | Sterol | USA | [125] |
| Pseudopterogorgia sp. | Anti-inflammatory | Secosterols | Sterol | USA | [125] |
| Pseudopterogorgia acerosa | Antitumor | Bis(pseudopterane) amine | Dialkylamine | BHS | [126] |
| Pseudopterogorgia bipinnata | Antituberculosis | Bipinnapterolide B | Terpenoid | USA | [127] |
| Pseudopterogorgia bipinnata | Antimalarial | Caucanolide A and D | Diterpenoid | COL, PAN, USA | [128] |
| Pseudopterogorgia elisabethae | Antimicrobial | Pseudopterosin X | Diterpenoid | USA | [129] |
| Pseudopterogorgia elisabethae | Antituberculosis | Ileabethoxazole | Diterpenoid | USA | [130] |
| Pseudopterogorgia elisabethae | Antituberculosis | Homopseudopteroxazole | Diterpenoid | USA | [131] |
| Pseudopterogorgia elisabethae | Antituberculosis | Caribenols A and B | Terpenoid | USA | [132] |
| Pseudopterogorgia elisabethae | Antituberculosis | Elisapterosin B | Diterpenoid | USA | [133] |
| Pseudopterogorgia elisabethae | Antimalarial | Aberrarone | Diterpenoid | COL | [134] |
| Pseudopterogorgia kallos | Antimalarial | Bielschowskysin | Diterpenoid | PAN, USA | [135] |
| Pseudopterogorgia kallos | Antitumor | Bielschowskysin | Diterpenoid | PAN, USA | [135] |
| Pseudopterogorgia rígida | Antimicrobial | Curcuphenol | Terpenoid | USA | [136] |
| Isididae | |||||
| Isis hippuris | Antitumor | Suberosenol B | Terpenoid | TAIW | [114] |
| Isis hippuris | Antitumor | Polyoxygenated steroids | Steroid | IND | [115,117] |
| Isis hippuris | Antitumor | A –nor-hippuristanol | Steroid | TAIW | [116] |
| Isis hippuris | Antitumor | Isishippuric acid B | Steroid | TAIW | [116] |
| Plexauridae | |||||
| Eunicea sp. | Antimalarial | Sesquiterpenoids | Sesquiterpenoid | COL, PAN, USA | [137] |
| Eunicea fusca | Anti-inflammatory | Fuscisides | Diterpenoid | USA | [138] |
| Euplexaura flava | Anti-inflammatory | Butenolide | Lipid | JPN | [139] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Mar. Drugs 2011, 9, 1860-1886. https://doi.org/10.3390/md9101860
Rocha J, Peixe L, Gomes NCM, Calado R. Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Marine Drugs. 2011; 9(10):1860-1886. https://doi.org/10.3390/md9101860
Chicago/Turabian StyleRocha, Joana, Luisa Peixe, Newton C.M. Gomes, and Ricardo Calado. 2011. "Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting" Marine Drugs 9, no. 10: 1860-1886. https://doi.org/10.3390/md9101860
APA StyleRocha, J., Peixe, L., Gomes, N. C. M., & Calado, R. (2011). Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Marine Drugs, 9(10), 1860-1886. https://doi.org/10.3390/md9101860