Antitumor Peptides from Marine Organisms

Abstract
:1. Introduction
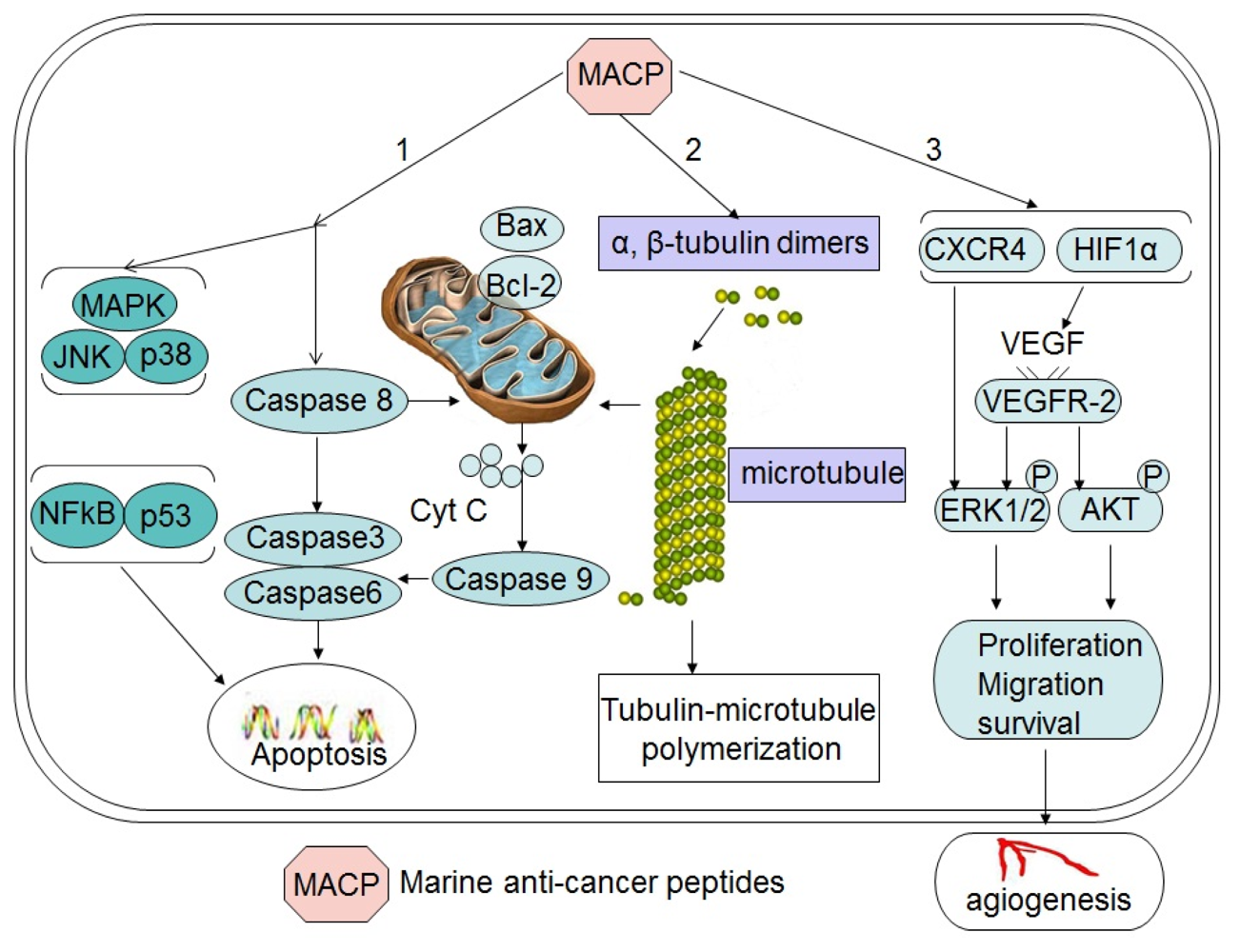
2. Peptides that Induce Apoptosis
2.1. Peptides that Activate the Intrinsic Mitochondrial Pathway
2.2. Peptides that Target the JNK or p38 MAPK Pathway
2.3. Peptides with an Unknown Mechanism of Apoptosis-Inducing Activity
3. Peptides that Affect the Tubulin-Microtubule Equilibrium
4. Peptides that Inhibit Angiogenesis
5. Peptides with Unknown Mechanism for Their Anti-Tumor Activity
6. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- de Vries, DJ; Beart, PM. Fishing for drugs from the sea: Status and strategies. Trends Pharmacol. Sci 1995, 16, 275–279. [Google Scholar]
- Reed, JC. Mechanisms of apoptosis. Am. J. Pathol 2000, 157, 1415–1430. [Google Scholar]
- Danial, NN; Korsmeyer, SJ. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar]
- Thompson, CB. Apoptosis in the pathogenesis and treatment of disease. Science 1995, 267, 1456–1462. [Google Scholar]
- Rowinsky, EK. Targeted induction of apoptosis in cancer management: The emerging role of tumor necrosis factor-related apoptosis-inducing ligand receptor activating agents. J. Clin. Oncol 2005, 23, 9394–9407. [Google Scholar]
- Call, JA; Eckhardt, SG; Camidge, DR. Targeted manipulation of apoptosis in cancer treatment. Lancet Oncol 2008, 9, 1002–1011. [Google Scholar]
- Iannolo, G; Conticello, C; Memeo, L; de Maria, R. Apoptosis in normal and cancer stem cells. Crit. Rev. Oncol. Hematol 2008, 66, 42–51. [Google Scholar]
- Burz, C; Berindan-Neagoe, I; Balacescu, O; Irimie, A. Apoptosis in cancer: Key molecular signaling pathways and therapy targets. Acta Oncol 2009, 48, 811–821. [Google Scholar]
- Fulda, S; Pervaiz, S. Apoptosis signaling in cancer stem cells. Int. J. Biochem. Cell Biol 2009, 42, 31–38. [Google Scholar]
- Ghobrial, IM; Witzig, TE; Adjei, AA. Targeting apoptosis pathways in cancer therapy. CA Cancer J. Clin 2005, 55, 178–194. [Google Scholar]
- Eberle, J; Fecker, LF; Forschner, T; Ulrich, C; Rowert-Huber, J; Stockfleth, E. Apoptosis pathways as promising targets for skin cancer therapy. Br. J. Dermatol 2007, 156S, 18–24. [Google Scholar]
- Ziegler, DS; Kung, AL. Therapeutic targeting of apoptosis pathways in cancer. Curr. Opin. Oncol 2008, 20, 97–103. [Google Scholar]
- Cheng, L; Wang, X; Zhang, J; Zhang, SB; Zheng, SQ; Zheng, J. Targeting apoptosis signaling pathways in cancer therapy. Zhonghua Bing Li Xue Za Zhi 2009, 38, 639–642. [Google Scholar]
- Lin, X; Liu, M; Hu, C; Liao, DJ. Targeting cellular proapoptotic molecules for developing anticancer agents from marine sources. Curr. Drug Targets 2010, 11, 708–715. [Google Scholar]
- von Schwarzenberg, K; Vollmar, AM. Targeting apoptosis pathways by natural compounds in cancer: Marine compounds as lead structures and chemical tools for cancer therapy. Cancer Lett 2010. [Google Scholar] [CrossRef]
- Abe, K; Kurakin, A; Mohseni-Maybodi, M; Kay, B; Khosravi-Far, R. The complexity of TNF-related apoptosis-inducing ligand. Ann. N. Y. Acad. Sci 2000, 926, 52–63. [Google Scholar]
- Ozoren, N; El-Deiry, WS. Cell surface Death Receptor signaling in normal and cancer cells. Semin. Cancer Biol 2003, 13, 135–147. [Google Scholar]
- Peter, ME; Krammer, PH. The CD95(APO-1/Fas) DISC and beyond. Cell Death Differ 2003, 10, 26–35. [Google Scholar]
- Thorburn, A. Death receptor-induced cell killing. Cell Signal 2004, 16, 139–144. [Google Scholar]
- Kroemer, G. Mitochondrial control of apoptosis: An introduction. Biochem. Biophys. Res. Commun 2003, 304, 433–435. [Google Scholar]
- Gupta, S; Kass, GE; Szegezdi, E; Joseph, B. The mitochondrial death pathway: A promising therapeutic target in diseases. J. Cell. Mol. Med 2009, 13, 1004–1033. [Google Scholar]
- Yip, KW; Reed, JC. Bcl-2 family proteins and cancer. Oncogene 2008, 27, 6398–6406. [Google Scholar]
- Okun, I; Balakin, KV; Tkachenko, SE; Ivachtchenko, AV. Caspase activity modulators as anticancer agents. Anticancer Agents Med. Chem 2008, 8, 322–341. [Google Scholar]
- Ghavami, S; Hashemi, M; Ande, SR; Yeganeh, B; Xiao, W; Eshraghi, M; Bus, CJ; Kadkhoda, K; Wiechec, E; Halayko, AJ; Los, M. Apoptosis and cancer: Mutations within caspase genes. J. Med. Genet 2009, 46, 497–510. [Google Scholar]
- Park, HJ; Kim, BC; Kim, SJ; Choi, KS. Role of MAP kinases and their cross-talk in TGF-beta1-induced apoptosis in FaO rat hepatoma cell line. Hepatology 2002, 35, 1360–1371. [Google Scholar]
- Kang, MH; Reynolds, CP. Bcl-2 inhibitors: Targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res 2009, 15, 1126–1132. [Google Scholar]
- Wong, WW; Puthalakath, H. Bcl-2 family proteins: The sentinels of the mitochondrial apoptosis pathway. IUBMB Life 2008, 60, 390–397. [Google Scholar]
- Braekman, JC; Daloze, D; Moussiaux, B; Riccio, R. Jaspamide from the marine sponge Jaspis johnstoni. J. Nat. Prod 1987, 50, 994–995. [Google Scholar]
- Odaka, C; Sanders, ML; Crews, P. Jasplakinolide induces apoptosis in various transformed cell lines by a caspase-3-like protease-dependent pathway. Clin. Diagn. Lab. Immunol 2000, 7, 947–952. [Google Scholar]
- Cioca, DP; Kitano, K. Induction of apoptosis and CD10/neutral endopeptidase expression by jaspamide in HL-60 line cells. Cell. Mol. Life Sci 2002, 59, 1377–1387. [Google Scholar]
- Nogle, LM; Gerwick, WH. Somocystinamide A, a novel cytotoxic disulfide dimer from a Fijian marine cyanobacterial mixed assemblage. Org. Lett 2002, 4, 1095–1098. [Google Scholar]
- Wrasidlo, W; Mielgo, A; Torres, VA; Barbero, S; Stoletov, K; Suyama, TL; Klemke, RL; Gerwick, WH; Carson, DA; Stupack, DG. The marine lipopeptide somocystinamide A triggers apoptosis via caspase 8. Proc. Natl. Acad. Sci. USA 2008, 105, 2313–2318. [Google Scholar]
- Schirmer, T; Huber, R; Schneider, M; Bode, W; Miller, M; Hackert, ML. Crystal structure analysis and refinement at 2.5 Å of hexameric C-phycocyanin from the cyanobacterium Agmenellum quadruplicatum: The molecular model and its implications for light-harvesting. J. Mol. Biol 1986, 188, 651–676. [Google Scholar]
- Li, B; Gao, MH; Zhang, XC; Chu, XM. Molecular immune mechanism of C-phycocyanin from Spirulina platensis induces apoptosis in HeLa cells in vitro. Biotechnol. Appl. Biochem 2006, 43, 155–164. [Google Scholar]
- Dhanasekaran, DN; Reddy, EP. JNK signaling in apoptosis. Oncogene 2008, 27, 6245–6251. [Google Scholar]
- Wagner, EF; Nebreda, AR. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar]
- Depenbrock, H; Peter, R; Faircloth, GT; Manzanares, I; Jimeno, J; Hanauske, AR. In vitro activity of aplidine, a new marine-derived anti-cancer compound, on freshly explanted clonogenic human tumour cells and haematopoietic precursor cells. Br. J. Cancer 1998, 78, 739–744. [Google Scholar]
- Urdiales, JL; Morata, P; de Castro, IN; Sánchez-Jiménez, F. Antiproliferative effect of dehydrodidemnin B (DDB), a depsipeptide isolated from Mediterranean tunicates. Cancer Lett 1996, 102, 31–37. [Google Scholar]
- Garcia-Fernandez, LF; Losada, A; Alcaide, V; Alvarez, AM; Cuadrado, A; Gonzalez, L; Nakayama, K; Nakayama, KI; Fernandez-Sousa, JM; Munoz, A; et al. Aplidin induces the mitochondrial apoptotic pathway via oxidative stress-mediated JNK and p38 activation and protein kinase C delta. Oncogene 2002, 21, 7533–7544. [Google Scholar]
- Rinehart, KL. Antitumor compounds from tunicates. Med. Res. Rev 2000, 20, 1–27. [Google Scholar]
- Faivre, S; Chieze, S; Delbaldo, C; Ady-Vago, N; Guzman, C; Lopez-Lazaro, L; Lozahic, S; Jimeno, J; Pico, F; Armand, JP; et al. Phase I and pharmacokinetic study of aplidine, a new marine cyclodepsipeptide in patients with advanced malignancies. J. Clin. Oncol 2005, 23, 7871–7880. [Google Scholar]
- Maroun, JA; Belanger, K; Seymour, L; Matthews, S; Roach, J; Dionne, J; Soulieres, D; Stewart, D; Goel, R; Charpentier, D; et al. Phase I study of Aplidine in a dailyx5 one-hour infusion every 3 weeks in patients with solid tumors refractory to standard therapy. A National Cancer Institute of Canada Clinical Trials Group study: NCIC CTG IND 115. Ann. Oncol 2006, 17, 1371–1378. [Google Scholar]
- Nalda-Molina, R; Valenzuela, B; Ramon-Lopez, A; Miguel-Lillo, B; Soto-Matos, A; Perez-Ruixo, JJ. Population pharmacokinetics meta-analysis of plitidepsin (Aplidin) in cancer subjects. Cancer Chemother. Pharmacol 2009, 64, 97–108. [Google Scholar]
- Rinehart, KL; Gloer, JB; Cook, JC; Mizsak, SA; Scahill, TA. Structures of the didemnins, antiviral and cytotoxic depsipeptides from a Caribbean tunicate. J. Am. Chem. Soc 1981, 103, 1857–1859. [Google Scholar]
- Rinehart, K, Jr; Gloer, J; Hughes, R, Jr; Renis, H; McGovren, J; Swynenberg, E; Stringfellow, D; Kuentzel, S; Li, L. Didemnins: Antiviral and antitumor depsipeptides from a caribbean tunicate. Science 1981, 212, 933–935. [Google Scholar]
- Schmitz, F; Bowden, B; Toth, S. Antitumor and cytotoxic compounds from marine organisms. Mar. Biotechnol 1993, 1, 197–308. [Google Scholar]
- Blunden, G. Biologically active compounds from marine organisms. Phytother. Res 2001, 15, 89–94. [Google Scholar]
- Grubb, DR; Wolvetang, EJ; Lawen, A. Didemnin B induces cell death by apoptosis: The fastest induction of apoptosis ever described. Biochem. Biophys. Res. Commun 1995, 215, 1130–1136. [Google Scholar]
- Johnson, KL; Lawen, A. Rapamycin inhibits didemnin B-induced apoptosis in human HL-60 cells: Evidence for the possible involvement of FK506-binding protein 25. Immunol. Cell Biol 1999, 77, 242–248. [Google Scholar]
- Vervoort, H; Fenical, W; Epifanio, RA. Tamandarins A and B: New cytotoxic depsipeptides from a Brazilian ascidian of the family Didemnidae. J. Org. Chem 2000, 65, 782–792. [Google Scholar]
- Chun, H; Davies, B; Hoth, D; Suffness, M; Plowman, J; Flora, K; Grieshaber, C; Leyland-Jones, B. Didemnin B. Investig. New Drugs 1986, 4, 279–284. [Google Scholar]
- Rinehart, KL; Holt, TG; Fregeau, NL; Keifer, PA; Wilson, GR; Perun, TJ; Sakai, R; Thompson, AG; Stroh, JG; Shield, LS; Seigler, DS; Li, LH; Martin, DG; Grimmelikhuijzen, CJP; Gäde, G. Bioactive Compounds from Aquatic and Terrestrial Sources. J. Nat. Prod 1990, 53, 771–792. [Google Scholar]
- Hwang, Y; Rowley, D; Rhodes, D; Gertsch, J; Fenical, W; Bushman, F. Mechanism of inhibition of a poxvirus topoisomerase by the marine natural product sansalvamide A. Mol. Pharmacol 1999, 55, 1049–1053. [Google Scholar]
- Pan, PS; Vasko, RC; Lapera, SA; Johnson, VA; Sellers, RP; Lin, CC; Pan, CM; Davis, MR; Ardi, VC; McAlpine, SR. A comprehensive study of Sansalvamide A derivatives: The structure-activity relationships of 78 derivatives in two pancreatic cancer cell lines. Bioorg. Med. Chem 2009, 17, 5806–5825. [Google Scholar]
- Hambley, TW; Hawkins, CJ; Lavin, MF; van den Brenk, A; Watters, DJ. Cycloxazoline: A cytotoxic cyclic hexapeptide from the ascidian lissoclinum bistratum. Tetrahedron 1992, 48, 341–348. [Google Scholar]
- Watters, DJ; Beamish, HJ; Marshall, KA; Gardiner, RA; Seymour, GJ; Lavin, MF. Accumulation of HL-60 leukemia cells in G2/M and inhibition of cytokinesis caused by two marine compounds, bistratene A and cycloxazoline. Cancer Chemother. Pharmacol 1994, 33, 399–409. [Google Scholar]
- Carroll, AR; Feng, Y; Bowden, BF; Coll, JC. Studies of Australian Ascidians. 5. Virenamides A–C, New Cytotoxic Linear Peptides from the Colonial Didemnid Ascidian Diplosoma virens. J. Org. Chem 1996, 61, 4059–4061. [Google Scholar]
- Hadfield, JA; Ducki, S; Hirst, N; McGown, AT. Tubulin and microtubules as targets for anticancer drugs. Prog. Cell Cycle Res 2003, 5, 309–325. [Google Scholar]
- Islam, MN; Iskander, MN. Microtubulin binding sites as target for developing anticancer agents. Mini Rev. Med. Chem 2004, 4, 1077–1104. [Google Scholar]
- Pettit, GR; Kamano, Y; Fujii, Y; Herald, CL; Inoue, M; Brown, P; Gust, D; Kitahara, K; Schmidt, JM; Doubek, DL; Michel, C. Marine animal biosynthetic constituents for cancer chemotherapy. J. Nat. Prod 1981, 44, 482–485. [Google Scholar]
- Luesch, H; Moore, RE; Paul, VJ; Mooberry, SL; Corbett, TH. Isolation of Dolastatin 10 from the Marine Cyanobacterium Symploca Species VP642 and Total Stereochemistry and Biological Evaluation of Its Analogue Symplostatin 1. J. Nat. Prod 2001, 64, 907–910. [Google Scholar]
- Bai, R; Pettit, GR; Hamel, E. Dolastatin 10, a powerful cytostatic peptide derived from a marine animal. Inhibition of tubulin polymerization mediated through the Vinca alkaloid binding domain. Biochem. Pharmacol 1990, 39, 1941–1949. [Google Scholar]
- Bai, RL; Pettit, GR; Hamel, E. Binding of dolastatin 10 to tubulin at a distinct site for peptide antimitotic agents near the exchangeable nucleotide and Vinca alkaloid sites. J. Biol. Chem 1990, 265, 17141–17149. [Google Scholar]
- Edler, MC; Fernandez, AM; Lassota, P; Ireland, CM; Barrows, LR. Inhibition of tubulin polymerization by vitilevuamide, a bicyclic marine peptide, at a site distinct from colchicine, the Vinca alkaloids, and dolastatin 10. Biochem. Pharmacol 2002, 63, 707–715. [Google Scholar]
- Cruz-Monserrate, Z; Vervoort, HC; Bai, R; Newman, DJ; Howell, SB; Los, G; Mullaney, JT; Williams, MD; Pettit, GR; Fenical, W; Hamel, E. Diazonamide A and a synthetic structural analog: Disruptive effects on mitosis and cellular microtubules and analysis of their interactions with tubulin. Mol. Pharmacol 2003, 63, 1273–1280. [Google Scholar]
- Lachia, M; Moody, CJ. The synthetic challenge of diazonamide A, a macrocyclic indole bis-oxazole marine natural product. Nat. Prod. Rep 2008, 25, 227–253. [Google Scholar]
- Schmidt, EW; Raventos-Suarez, C; Bifano, M; Menendez, AT; Fairchild, CR; Faulkner, DJ. Scleritodermin A, a cytotoxic cyclic peptide from the lithistid sponge Scleritoderma nodosum. J. Nat. Prod 2004, 67, 475–478. [Google Scholar]
- Liu, S; Cui, YM; Nan, FJ. Total synthesis of the originally proposed and revised structures of scleritodermin A. Org. Lett 2008, 10, 3765–3768. [Google Scholar]
- Anderson, HJ; Coleman, JE; Andersen, RJ; Roberge, M. Cytotoxic peptides hemiasterlin, hemiasterlin A and hemiasterlin B induce mitotic arrest and abnormal spindle formation. Cancer Chemother. Pharmacol 1997, 39, 223–226. [Google Scholar]
- Gamble, WR; Durso, NA; Fuller, RW; Westergaard, CK; Johnson, TR; Sackett, DL; Hamel, E; Cardellina Ii, JH; Boyd, MR. Cytotoxic and tubulin-interactive hemiasterlins from Auletta sp. and Siphonochalina spp. sponges. Bioorg. Med. Chem 1999, 7, 1611–1165. [Google Scholar]
- Loganzo, F; Discafani, CM; Annable, T; Beyer, C; Musto, S; Hari, M; Tan, X; Hardy, C; Hernandez, R; Baxter, M; et al. HTI-286, a synthetic analogue of the tripeptide hemiasterlin, is a potent antimicrotubule agent that circumvents P-glycoprotein-mediated resistance in vitro and in vivo. Cancer Res 2003, 63, 1838–1845. [Google Scholar]
- Yamashita, A; Norton, EB; Kaplan, JA; Niu, C; Loganzo, F; Hernandez, R; Beyer, CF; Annable, T; Musto, S; Discafani, C; et al. Synthesis and activity of novel analogs of hemiasterlin as inhibitors of tubulin polymerization: Modification of the A segment. Bioorg. Med. Chem. Lett 2004, 14, 5317–5322. [Google Scholar]
- Simmons, TL; Nogle, LM; Media, J; Valeriote, FA; Mooberry, SL; Gerwick, WH. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod 2009, 72, 1011–1016. [Google Scholar]
- Chevallier, C; Richardson, AD; Edler, MC; Hamel, E; Harper, MK; Ireland, CM. A new cytotoxic and tubulin-interactive milnamide derivative from a marine sponge Cymbastela sp. Org. Lett 2003, 5, 3737–3739. [Google Scholar]
- Ning, X; Zhao, J; Zhang, Y; Cao, S; Liu, M; Ling, P; Lin, X. A novel anti-tumor protein extracted from Meretrix meretrix Linnaeus induces cell death by increasing cell permeability and inhibiting tubulin polymerization. Int. J. Oncol 2009, 35, 805–812. [Google Scholar]
- Folkman, J. The role of angiogenesis in tumor growth. Semin. Cancer Biol 1992, 3, 65–71. [Google Scholar]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat. Med 1995, 1, 27–31. [Google Scholar]
- Bouck, N; Stellmach, V; Hsu, SC. How tumors become angiogenic. Adv. Cancer Res 1996, 69, 135–174. [Google Scholar]
- Folkman, J. Angiogenesis and angiogenesis inhibition: An overview. EXS 1997, 79, 1–8. [Google Scholar]
- Ferrara, N. VEGF: An update on biological and therapeutic aspects. Curr. Opin. Biotechnol 2000, 11, 617–624. [Google Scholar]
- Ferrara, N; Gerber, HP; LeCouter, J. The biology of VEGF and its receptors. Nat. Med 2003, 9, 669–676. [Google Scholar]
- Nakamura, S; Chikaraishi, Y; Tsuruma, K; Shimazawa, M; Hara, H. Ruboxistaurin, a PKCbeta inhibitor, inhibits retinal neovascularization via suppression of phosphorylation of ERK1/2 and Akt. Exp. Eye Res 2009, 90, 137–145. [Google Scholar]
- Ushio-Fukai, M. Redox signaling in angiogenesis: Role of NADPH oxidase. Cardiovasc. Res 2006, 71, 226–235. [Google Scholar]
- Chiavarina, B; Whitaker-Menezes, D; Migneco, G; Martinez-Outschoorn, UE; Pavlides, S; Howell, A; Tanowitz, HB; Casimiro, MC; Wang, C; Pestell, RG; et al. HIF1-alpha functions as a tumor promoter in cancer associated fibroblasts, and as a tumor suppressor in breast cancer cells: Autophagy drives compartment-specific oncogenesis. Cell Cycle 2010, 9, 3534–3551. [Google Scholar]
- Fukushima, K; Murata, M; Hachisuga, M; Tsukimori, K; Seki, H; Takeda, S; Asanoma, K; Wake, N. Hypoxia inducible factor 1 alpha regulates matrigel-induced endovascular differentiation under normoxia in a human extravillous trophoblast cell line. Placenta 2008, 29, 324–331. [Google Scholar]
- Lee, A; Langer, R. Shark cartilage contains inhibitors of tumor angiogenesis. Science 1983, 221, 1185–1187. [Google Scholar]
- Dupont, E; Brazeau, P; Juneau, C. Extracts of shark cartilage having an anti-angiogenic activity and an effect on tumor regression: Process of making thereof. US Patents 5,985,839, 16 November 1999. [Google Scholar]
- Lee, SY; Chung, SM. Neovastat (AE-941) inhibits the airway inflammation via VEGF and HIF-2 alpha suppression. Vasc. Pharmacol 2007, 47, 313–318. [Google Scholar]
- Zheng, L; Ling, P; Wang, Z; Niu, R; Hu, C; Zhang, T; Lin, X. A novel polypeptide from shark cartilage with potent anti-angiogenic activity. Cancer Biol. Ther 2007, 6, 775–780. [Google Scholar]
- Morgan, JB; Mahdi, F; Liu, Y; Coothankandaswamy, V; Jekabsons, MB; Johnson, TA; Sashidhara, KV; Crews, P; Nagle, DG; Zhou, YD. The marine sponge metabolite mycothiazole: A novel prototype mitochondrial complex I inhibitor. Bioorg. Med. Chem 2010, 18, 5988–5994. [Google Scholar]
- McDonald, LA; Ireland, CM. Patellamide E: A new cyclic peptide from the ascidian Lissoclinum patella. J. Nat. Prod 1992, 55, 376–379. [Google Scholar]
- Rashid, MA; Gustafson, KR; Cardellina, JH, II; Boyd, MR; Patellamide, F. A new cytotoxic cyclic peptide from the colonial ascidian Lissoclinum patella. J. Nat. Prod 1995, 58, 594–597. [Google Scholar]
- Fu, X; Do, T; Schmitz, FJ; Andrusevich, V; Engel, MH. New cyclic peptides from the ascidian Lissoclinum patella. J. Nat. Prod 1998, 61, 1547–1551. [Google Scholar]
- Fu, X; Su, J; Zeng, L. Prepatellamide A, a new cyclic peptide from the ascidian Lissoclinum patella. Sci. China Ser. B Chem 2000, 43, 643–648. [Google Scholar]
- Taylor, SW; Craig, AG; Fischer, WH; Park, M; Lehrer, RI. Styelin D, an extensively modified antimicrobial peptide from ascidian hemocytes. J. Biol. Chem 2000, 275, 38417–38426. [Google Scholar]
- Swersey, JC; Ireland, CM; Cornell, LM; Peterson, RW. Eusynstyelamide, a highly modified dimer peptide from the ascidian Eusynstyela misakiensis. J. Nat. Prod 1994, 57, 842–845. [Google Scholar]
- McDonald, LA; Christopher Swersey, J; Ireland, CM; Carroll, AR; Coll, JC; Bowden, BF; Fairchild, CR; Cornell, L. Botryllamides A–D, new brominated tyrosine derivatives from styelid ascidians of the genus Botryllus. Tetrahedron 1995, 51, 5237–5244. [Google Scholar]
- Degnan, BM; Hawkins, CJ; Lavin, MF; McCaffrey, EJ; Parry, DL; van den Brenk, AL; Watters, DJ. New cyclic peptides with cytotoxic activity from the ascidian Lissoclinum patella. J. Med. Chem 1989, 32, 1349–1354. [Google Scholar]
- Hawkins, CJ; Lavin, MF; Marshall, KA; van den Brenk, AL; Watters, DJ. Structure-activity relationships of the lissoclinamides: Cytotoxic cyclic peptides from the ascidian Lissoclinum patella. J. Med. Chem 1990, 33, 1634–1638. [Google Scholar]
- Donia, MS; Wang, B; Dunbar, DC; Desai, PV; Patny, A; Avery, M; Hamann, MT. Mollamides B and C, Cyclic hexapeptides from the indonesian tunicate Didemnum molle. J. Nat. Prod 2008, 71, 941–945. [Google Scholar]
- Carroll, AR; Bowden, BF; Coll, JC; Hockless, DCR; Skelton, BW; White, AH. Studies of Australian Ascidians. IV. Mollamide, a Cytotoxic Cyclic Heptapeptide from the Compound Ascidian Diplosoma virens. Aust. J. Chem 1994, 47, 61–69. [Google Scholar]
- Sonnenschein, RN; Farias, JJ; Tenney, K; Mooberry, SL; Lobkovsky, E; Clardy, J; Crews, P. A further study of the cytotoxic constituents of a milnamide-producing sponge. Org. Lett 2004, 6, 779–782. [Google Scholar]
- Coleman, JE; van Soest, R; Andersen, RJ. New geodiamolides from the sponge Cymbastela sp. collected in Papua New Guinea. J. Nat. Prod 1999, 62, 1137–1141. [Google Scholar]
- Fusetani, N; Sugawara, T; Matsunaga, S; Hirota, H. Orbiculamide A: A novel cytotoxic cyclic peptide from a marine sponge Theonella sp. J. Am. Chem. Soc 1991, 113, 7811–7812. [Google Scholar]
- Araki, T; Matsunaga, S; Nakao, Y; Furihata, K; West, L; Faulkner, DJ; Fusetani, N. Koshikamide B, a cytotoxic peptide lactone from a marine sponge Theonella sp. J. Org. Chem 2008, 73, 7889–7894. [Google Scholar]
- Pettit, GR; Cichacz, Z; Barkoczy, J; Dorsaz, AC; Herald, DL; Williams, MD; Doubek, DL; Schmidt, JM; Tackett, LP; Brune, DC; et al. Isolation and structure of the marine sponge cell growth inhibitory cyclic peptide phakellistatin 1. J. Nat. Prod 1993, 56, 260–267. [Google Scholar]
- Pettit, GR; Tan, R; Ichihara, Y; Williams, MD; Doubek, DL; Tackett, LP; Schmidt, JM; Cerny, RL; Boyd, MR; Hooper, JN. Antineoplastic agents, 325. Isolation and structure of the human cancer cell growth inhibitory cyclic octapeptides phakellistatin 10 and 11 from Phakellia sp. J. Nat. Prod 1995, 58, 961–965. [Google Scholar]
- Pettit, GR; Tan, R. Antineoplastic agents 390. Isolation and structure of phakellistatin 12 from a Chuuk archipelago marine sponge. Bioorg. Med. Chem. Lett 2003, 13, 685–688. [Google Scholar]
- Pettit, GR; Tan, R. Isolation and structure of phakellistatin 14 from the Western Pacific marine sponge Phakellia sp. J. Nat. Prod 2005, 68, 60–63. [Google Scholar]
- Davis, RA; Mangalindan, GC; Bojo, ZP; Antemano, RR; Rodriguez, NO; Concepcion, GP; Samson, SC; de Guzman, D; Cruz, LJ; Tasdemir, D; Harper, MK; Feng, X; Carter, GT; Ireland, CM. Microcionamides A and B, Bioactive Peptides from the Philippine Sponge Clathria (Thalysias) abietina. J. Org. Chem 2004, 69, 4170–4176. [Google Scholar]
- Li, H; Matsunaga, S; Fusetani, N. Halicylindramides D and E, antifungal peptides from the marine sponge Halichondria cylindrata. J. Nat. Prod 1996, 59, 163–166. [Google Scholar]
- Rashid, MA; Gustafson, KR; Boswell, JL; Boyd, MR. Haligramides A and B, two new cytotoxic hexapeptides from the marine sponge Haliclona nigra. J. Nat. Prod 2000, 63, 956–959. [Google Scholar]
- Bai, R; Durso, NA; Sackett, DL; Hamel, E. Interactions of the sponge-derived antimitotic tripeptide hemiasterlin with tubulin: Comparison with dolastatin 10 and cryptophycin 1. Biochemistry 1999, 38, 14302–14310. [Google Scholar]
- Crews, P; Farias, JJ; Emrich, R; Keifer, PA. Milnamide A, an unusual cytotoxic tripeptide from the marine sponge Auletta cf. constricta. J. Org. Chem 1994, 59, 2932–2934. [Google Scholar]
- Laird, DW; LaBarbera, DV; Feng, X; Bugni, TS; Harper, MK; Ireland, CM. Halogenated cyclic peptides isolated from the sponge Corticium sp. J. Nat. Prod 2007, 70, 741–746. [Google Scholar]
- Ratnayake, AS; Bugni, TS; Feng, X; Harper, MK; Skalicky, JJ; Mohammed, KA; Andjelic, CD; Barrows, LR; Ireland, CM. Theopapuamide, a cyclic depsipeptide from a Papua New Guinea lithistid sponge Theonella swinhoei. J. Nat. Prod 2006, 69, 1582–1586. [Google Scholar]
- Bishara, A; Rudi, A; Aknin, M; Neumann, D; Ben-Califa, N; Kashman, Y. Taumycins A and B, two bioactive lipodepsipeptides from the Madagascar sponge Fascaplysinopsis sp. Org. Lett 2008, 10, 4307–4309. [Google Scholar]
- Fusetani, N; Warabi, K; Nogata, Y; Nakao, Y; Matsunaga, S; van Soest, RRM. Koshikamide A1, a new cytotoxic linear peptide isolated from a marine sponge, Theonella sp. Tetrahedron Lett 1999, 40, 4687–4690. [Google Scholar]
- Araki, T; Matsunaga, S; Fusetani, N. Koshikamide A2, a cytotoxic linear undecapeptide isolated from a marine sponge of Theonella sp. Biosci. Biotechnol. Biochem 2005, 69, 1318–1322. [Google Scholar]
- Boot, CM; Tenney, K; Valeriote, FA; Crews, P. Highly N-methylated linear peptides produced by an atypical sponge-derived Acremonium sp. J. Nat. Prod 2006, 69, 83–92. [Google Scholar]
- Wesson, KJ; Hamann, MT. Keenamide A, a bioactive cyclic peptide from the marine mollusk Pleurobranchus forskalii. J. Nat. Prod 1996, 59, 629–631. [Google Scholar]
- Kimura, J; Takada, Y; Inayoshi, T; Nakao, Y; Goetz, G; Yoshida, WY; Scheuer, PJ. Kulokekahilide-1, a cytotoxic depsipeptide from the cephalaspidean mollusk Philinopsis speciosa. J. Org. Chem 2002, 67, 1760–1767. [Google Scholar]
- Nakao, Y; Yoshida, WY; Takada, Y; Kimura, J; Yang, L; Mooberry, SL; Scheuer, PJ. Kulokekahilide-2, a cytotoxic depsipeptide from a cephalaspidean mollusk Philinopsis speciosa. J. Nat. Prod 2004, 67, 1332–1340. [Google Scholar]
- Yu, Z; Lang, G; Kajahn, I; Schmaljohann, R; Imhoff, JF. Scopularides A and B, cyclodepsipeptides from a marine sponge-derived fungus, Scopulariopsis brevicaulis. J. Nat. Prod 2008, 71, 1052–1054. [Google Scholar]
- Linington, RG; Edwards, DJ; Shuman, CF; McPhail, KL; Matainaho, T; Gerwick, WH. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine Cyanobacterium Symploca sp. J. Nat. Prod 2008, 71, 22–27. [Google Scholar]
- Stolze, SC; Meltzer, M; Ehrmann, M; Kaiser, M. Solid phase total synthesis of the 3-amino-6-hydroxy-2-piperidone (Ahp) cyclodepsipeptide and protease inhibitor Symplocamide A. Chem. Commun 2010, 46, 8857–8859. [Google Scholar]
- Gutierrez, M; Suyama, TL; Engene, N; Wingerd, JS; Matainaho, T; Gerwick, WH. Apratoxin D, a potent cytotoxic cyclodepsipeptide from papua new guinea collections of the marine cyanobacteria Lyngbya majuscula and Lyngbya sordida. J. Nat. Prod 2008, 71, 1099– 1103. [Google Scholar]
- Liu, L; Rein, KS. New peptides isolated from Lyngbya species: A review. Mar. Drugs 2010, 8, 1817–1837. [Google Scholar]
- Andrianasolo, EH; Goeger, D; Gerwick, WH. Mitsoamide: A cytotoxic linear lipopeptide from the Madagascar marine cyanobacterium Geitlerinema sp. Pure Appl. Chem 2007, 79, 593–602. [Google Scholar]
- Maki, A; Diwakaran, H; Redman, B; al-Asfar, S; Pettit, GR; Mohammad, RM; al-Katib, A. The bcl-2 and p53 oncoproteins can be modulated by bryostatin 1 and dolastatins in human diffuse large cell lymphoma. Anticancer Drugs 1995, 6, 392–397. [Google Scholar]
- Taraboletti, G; Poli, M; Dossi, R; Manenti, L; Borsotti, P; Faircloth, GT; Broggini, M; D’Incalci, M; Ribatti, D; Giavazzi, R. Antiangiogenic activity of aplidine, a new agent of marine origin. Br. J. Cancer 2004, 90, 2418–2424. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Name of peptides | Sources | Class/types | Mode of action and references |
|---|---|---|---|
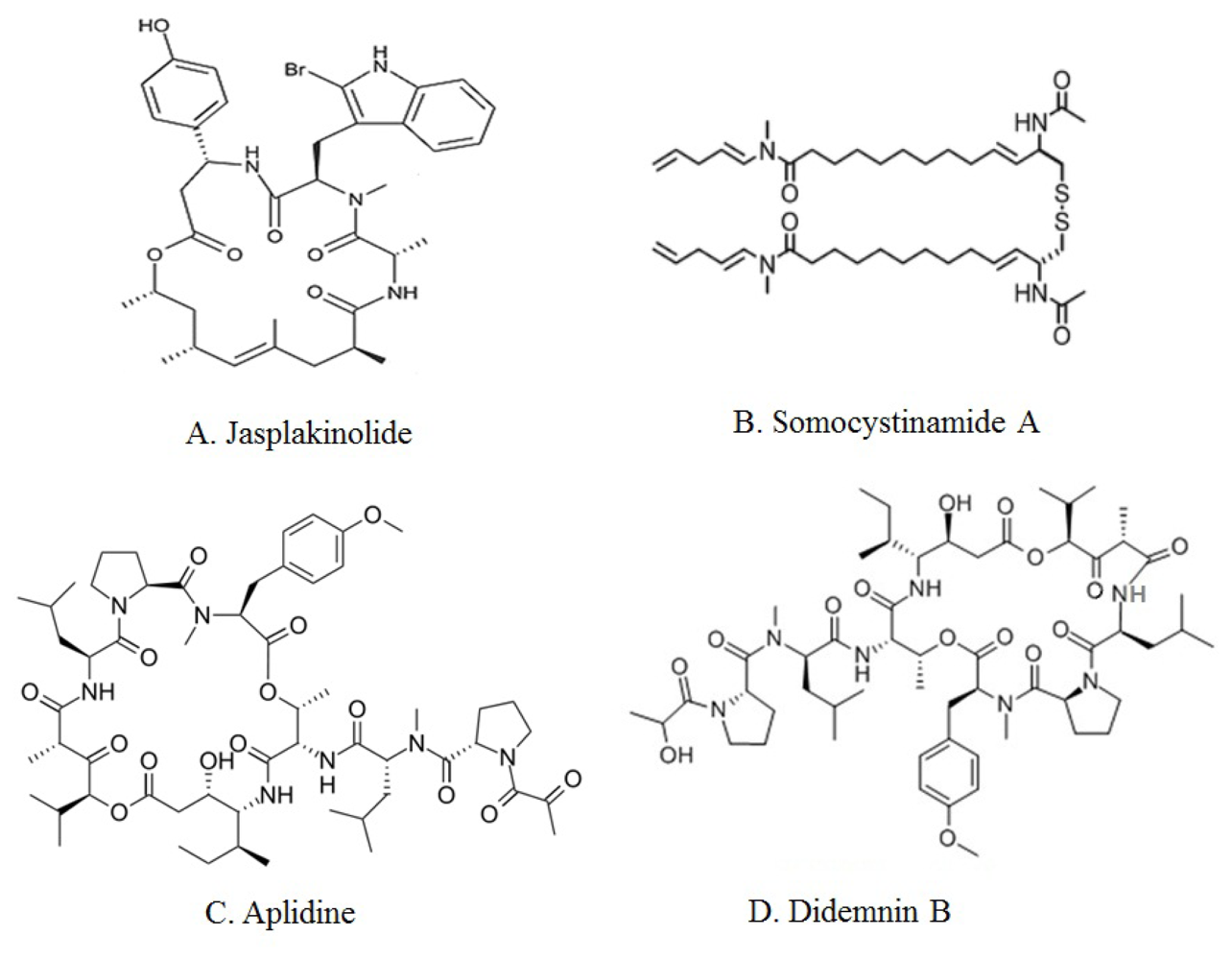
| Jaspamide (Jasplakinolide) | Marine sponge, Jaspis johnstoni | Cyclic depsipeptide | Caspase-3 activation, decreasing in Bcl-2 protein expression [28–30] |
| Somocystinamide A (ScA) | Lyngbya majuscula/Schizothrix sp. assemblage of marine cyanobacteria | Lipopeptide | Caspase-8 activation [31,32] |
| C-phycocyanin (C-PC) | Cyanobacteria Agmenellum quadruplicatum, Mastigocladus laminosus, Spirulina platensis | Tetrapyrrole-protein complex | Caspases-dependent apoptosis [34] |
| Aplidine (dehydrodidemnin B, DDB, Aplidin) | Tunicate, Aplidium albicans | Cyclic depsipeptide | JNK and p38 MAPK phosphorylation activation [37–39] |
| Didemnin B | Tunicate, Trididemnum solidum | Cyclic depsipeptide | Apoptosis, but unclear [44–49] |
| Sansalvamide A | Marine fungus | Cyclic depsipeptide | Apoptosis, but unclear [53,54] |
| Cycloxazoline | Marine ascidian, Lissoclinum bistratum | Cyclic depsipeptide | Apoptosis, but unclear [55,56] |
| Virenamides A–C | Didemnid ascidian, Diplosoma virens | Linear tripeptides | Apoptosis, but unclear [57] |
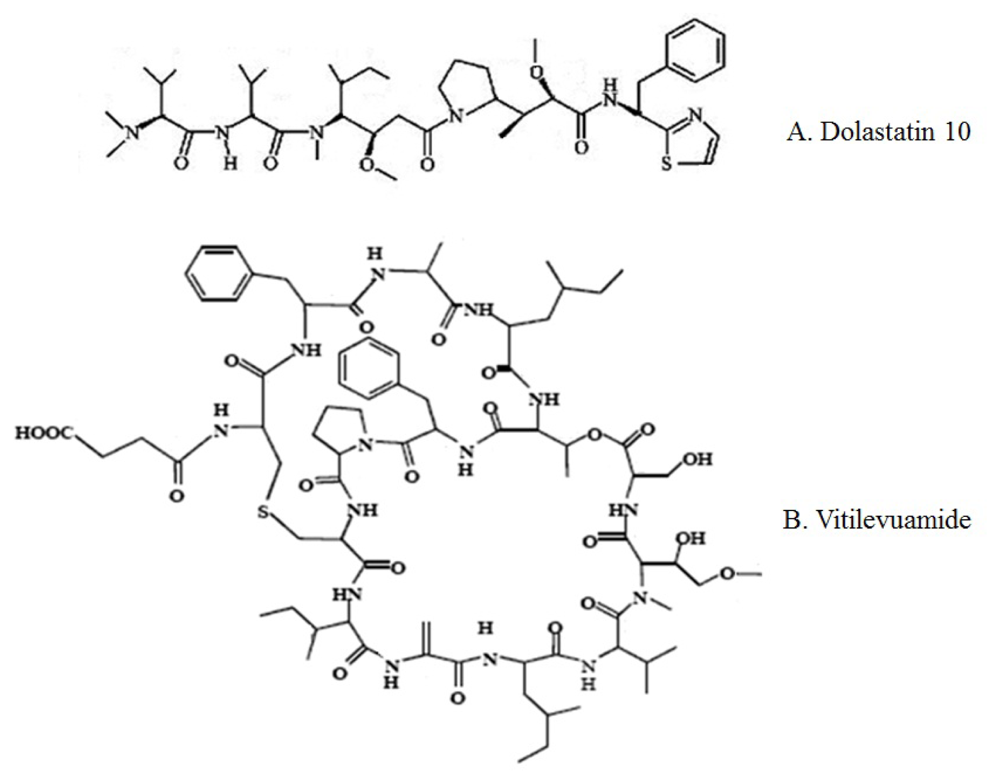
| Dolastatin 10 | Marine mollusk, Dolabella auricularia | Linear peptide | Microtubule assembly Inhibition [60,62] |
| Vitilevuamide | Marine ascidians, Didemnum cuculiferum and Polysyncranton lithostrotum | Bicyclic peptide | Tubulin polymerization inhibition [64] |
| Diazonamide | Marine ascidian, Diazona angulata | Macrocyclic peptide | Tubulin polymerization inhibition [65,66] |
| Scleritodermin A | Lithistid sponge, Scleritoderma nodosum | Cyclic peptide | Tubulin polymerization inhibition [67,68] |
| Hemiasterlin | Marine sponges, Auletta sp. and Siphonochalina sp. | Tripeptide | Tubulin polymerization inhibition [70–72] |
| DMMC | Cyanobacterium Lyngbya majuscula | Cyclic depsipeptide | Tubulin polymerization inhibition [73] |
| MML | Coelomic fluid, Meretrix meretrix | Protein | Tubulin polymerization inhibition [75] |
| Neovastat (AE-941) | Shark cartilage, Squalus acanthias | Extract < 500 kDa | VEGF and HIF2 alpha pathway inhibition [86–88] |
| PG155 | Shark cartilage, Prionace glauca | Polypeptide | VEGF induced angiogenesis inhibition [89] |
| Styelin D | Ascidian, Styela clava | C-terminally amidated peptide | Unknown [95] |
| Lissoclinamides | Aplousobranch ascidian, Lissoclinum patella | Cyclic peptide | Unknown [46,98,99] |
| Geodiamolides A–G | Caribbean sponge, Geodia sp. | Cyclic peptide | Unknown [102,103] |
| Orbiculamide A | Marine sponge, Theonella sp. | Cyclic peptide | Unknown [104] |
| Koshikamide B | Marine sponge, Theonella sp. | Peptide lactone | Unknown [105] |
| Phakellistatins | Marine sponges | Cyclic heptapeptides | Unknown [106–109] |
| Microcionamides A and B | Philippine Sponge, Clathria (Thalysias) abietina | Linear peptides | Unknown [110] |
| Keenamide A | Notaspidean mollusk, Pleurobranchus forskalii | Cyclic hexapeptide | Unknown [121] |
| Scopularides A and B | Fungus Scopulariopsis brevicaulis | Cyclodepsipeptide | Unknown [124] |
| Symplocamide A | Marine cyanobacteria Symploca sp. | Cyclodepsipeptide | Unknown [125,126] |
| Apratoxin D | Marine cyanobacteria Lyngbya majuscula and Lyngbya sordida | Macrocycle peptide | Unknown [127,128] |
| Mitsoamide | Marine cyanobacteria Geitlerinema sp. | Linear peptide | Unknown [129] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zheng, L.-H.; Wang, Y.-J.; Sheng, J.; Wang, F.; Zheng, Y.; Lin, X.-K.; Sun, M. Antitumor Peptides from Marine Organisms. Mar. Drugs 2011, 9, 1840-1859. https://doi.org/10.3390/md9101840
Zheng L-H, Wang Y-J, Sheng J, Wang F, Zheng Y, Lin X-K, Sun M. Antitumor Peptides from Marine Organisms. Marine Drugs. 2011; 9(10):1840-1859. https://doi.org/10.3390/md9101840
Chicago/Turabian StyleZheng, Lan-Hong, Yue-Jun Wang, Jun Sheng, Fang Wang, Yuan Zheng, Xiu-Kun Lin, and Mi Sun. 2011. "Antitumor Peptides from Marine Organisms" Marine Drugs 9, no. 10: 1840-1859. https://doi.org/10.3390/md9101840
APA StyleZheng, L.-H., Wang, Y.-J., Sheng, J., Wang, F., Zheng, Y., Lin, X.-K., & Sun, M. (2011). Antitumor Peptides from Marine Organisms. Marine Drugs, 9(10), 1840-1859. https://doi.org/10.3390/md9101840