Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities



Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Analyses
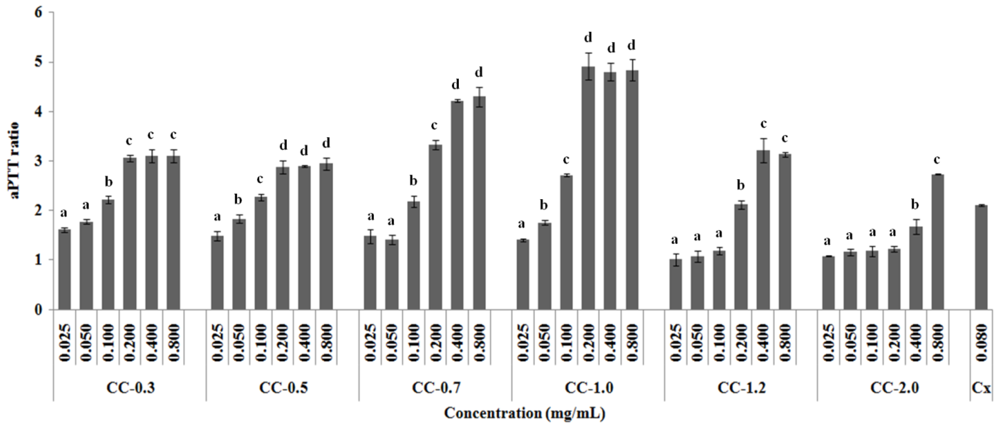
2.2. Anticoagulant Activity
2.3. Antioxidant Activity
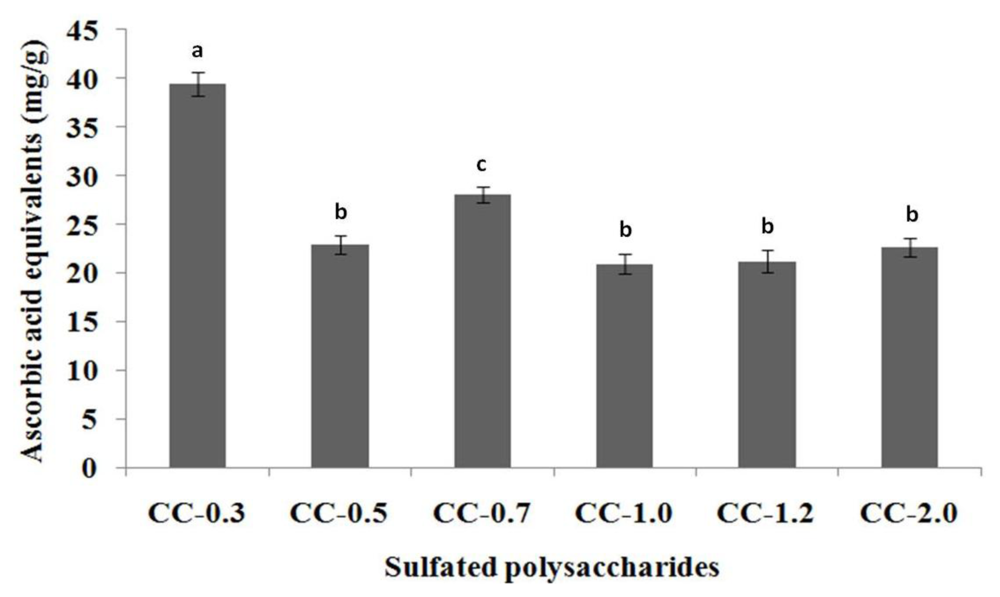
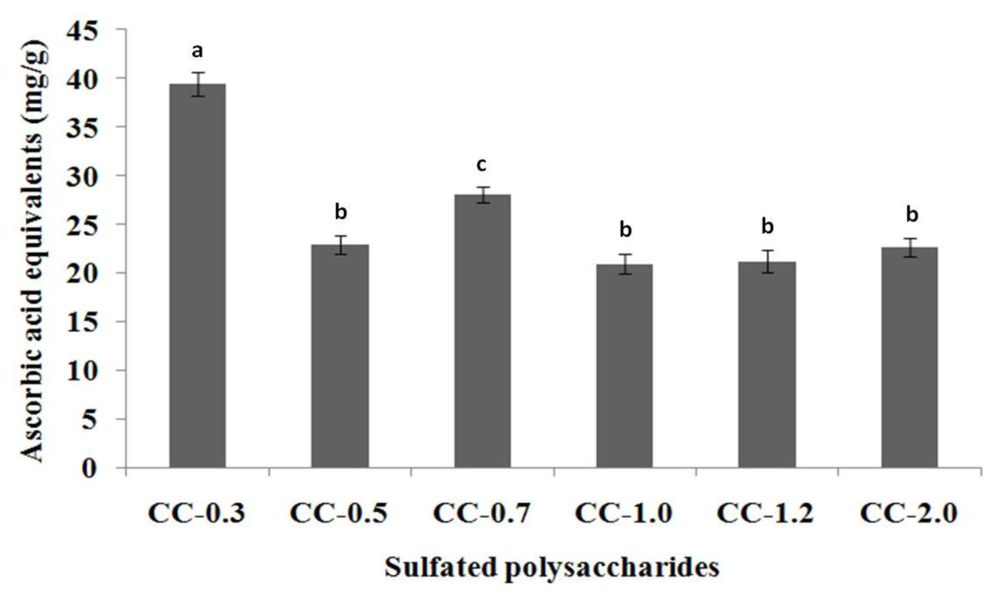
2.3.1. Total Antioxidant Capacity
2.3.2. Hydroxyl and Superoxide Radical Scavenging
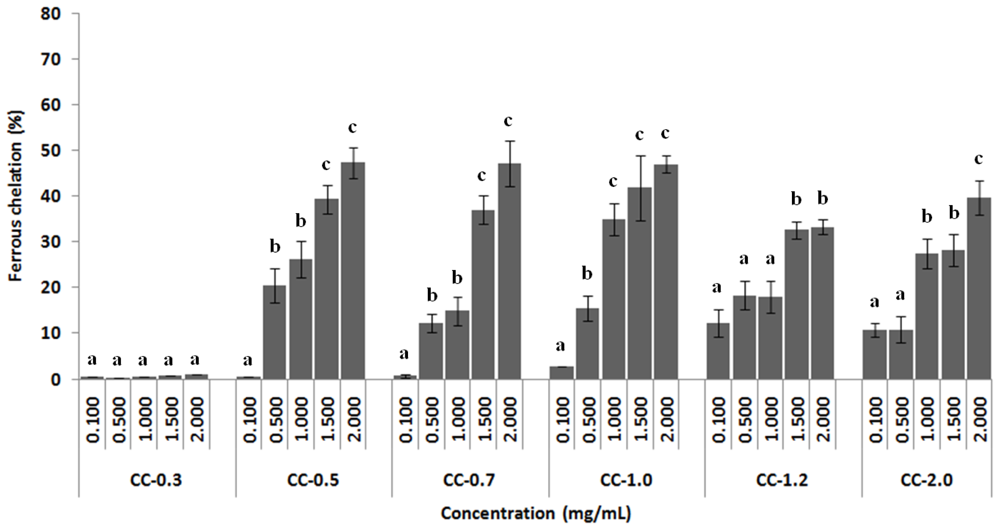
2.3.3. Chelating Effect on Ferrous Ions
3. Experimental Section
3.1. Materials
3.2. Extraction of Sulfated Polysaccharide Fractions
3.3. Agarose Gel Electrophoresis
3.4. Chemical Analysis and Monosaccharide Composition
3.5. Fourier Transformed Infrared Spectroscopy (FT-IR)
3.6. Anticoagulant Activity
3.7. Antioxidant Activity
3.7.1. Determination of Total Antioxidant Capacity
3.7.2. Hydroxyl Radical Scavenging Activity Assay
3.7.3. Superoxide Radical Scavenging Activity Assay
3.7.4. Ferric Chelating
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Cardozo, KHM; Guaratini, T; Barros, MP; Falcão, VR; Tonon, AP; Lopes, NP; Campos, S; Torres, MA; Souza, AO; Colepicolo, P; Pinto, E. Metabolites from algae with economical impact. Comp Biochem Physiol C Toxicol Pharmacol 2007, 146, 60–78. [Google Scholar]
- Costa, LS; Fidelis, GP; Cordeiro, SL; Oliveira, RM; Sabry, DA; Câmara, RBG; Nobre, LTDB; Costa, MSSP; Almeida-Lima, J; Farias, EHC; Leite, EL; Rocha, HAO. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed Pharmacother 2010, 64, 21–28. [Google Scholar]
- Fenoradosoa, TA; Delattre, C; Laroche, C; Wadouachi, A; Dulong, V; Picton, L; Andriamadio, P; Michaud, P. Highly sulphated galactan from Halymenia durvillei (Halymeniales, Rhodophyta), a red seaweed of Madagascar marine coasts. Int J Biol Macromol 2009, 45, 140–145. [Google Scholar]
- Farias, EHC; Pomin, VH; Valente, AP; Nader, HB; Rocha, HAO; Mourão, PAS. A preponderantly 4-sulfated, 3-linked galactan from the green alga Codium isthmocladium. Glycobiology 2008, 18, 250–259. [Google Scholar]
- Li, B; Lu, F; Wei, X; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Holtkamp, AD; Kelly, S; Ulber, R; Lang, S. Fucoidans and fucoidanases—focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl Microbiol Biotechnol 2009, 82, 1–11. [Google Scholar]
- Rocha, HAO; Moraes, FA; Trindade, ES; Franco, CR; Torquato, RJ; Veiga, SS; Valente, AP; Mourão, PA; Leite, EL; Nader, HB; Dietrich, CP. Structural and hemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schröederi. An ideal antithrombotic agent? J Biol Chem 2005, 280, 41278–41288. [Google Scholar]
- World Health Organization (WHO), The World Health Report 2008. Primary Health Care: Now More than Ever; World Health Organization: Geneva, Switzerland, 2008; pp. 1–148.
- Weitz, DS; Weitz, JI. Update on heparin: what do we need to know. J Thromb Thrombolysis 2010, 29, 199–207. [Google Scholar]
- Nader, HB; Lopes, CC; Rocha, HAO; Santos, EA; Dietrich, CP. Heparins and heparinoids: occurrence, structure, and mechanism of antithrombotic and hemorrhagic effect activities. Curr Pharm Des 2004, 10, 951–966. [Google Scholar]
- Freinbichler, W; Bianchi, L; Colivicchi, MA; Ballini, C; Tipton, KF; Linert, W; Corte, LD. The detection of hydroxyl radicals in vivo. J Inorg Biochem 2008, 102, 1329–1333. [Google Scholar]
- Barnham, KJ; Masters, CL; Bush, AI. Neurodegenerative diseases and oxidative stress. Nat Rev Drug Discov 2004, 3, 205–214. [Google Scholar]
- Hu, T; Liu, L; Chen, Y; Wu, J; Wang, S. Antioxidant activity of sulfated polysaccharide fractions extracted from Undaria pinnatifida in vitro. Int J Biol Macromol 2010, 46, 193–198. [Google Scholar]
- Zhang, Z; Wang, F; Wang, X; Liu, X; Hou, Y; Zhang, Q. Extraction of the polysaccharides from five algae and their potential antioxidant activity in vitro. Carbohydr Polym 2010, 82, 118–121. [Google Scholar]
- Wang, J; Zhang, Q; Zhang, Z; Song, H; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int J Biol Macromol 2010, 46, 6–12. [Google Scholar]
- Duarte, MER; Cardoso, MA; Noseda, MD; Cerezo, AS. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr Polym 2001, 333, 281–293. [Google Scholar]
- Leite, EL; Medeiros, MGL; Rocha, HAO; Farias, GGM; Silva, LF; Chavante, SF; Abreu, LD; Dietrich, CP; Nader, HB. Structure and pharmacological activities of a sulfated xylofucoglucuronan from the alga Spatoglossum schröederi. Plant Sci 1998, 132, 215–228. [Google Scholar]
- Dietrich, CP; Farias, GGM; Abreu, LR; Leite, EL; Silva, LF; Nader, HB. A new approach for the characterization of polysaccharides from algae: presence of four main acidic polysaccharides in three species of the class Phaeophycea. Plant Sci 1995, 108, 143–153. [Google Scholar]
- Albuquerque, IRL; Queiroz, KCS; Alves, LG; Santos, EA; Leite, EL; Rocha, HAO. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz J Med Biol Res 2004, 37, 167–171. [Google Scholar]
- Cumashi, A; Ushakova, NA; Preobrazhenskaya, ME; D’Incecco, A; Picooli, A; Totani, L; Tinari, N; Morozevich, GE; Berman, AE; Bilan, MI; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Zhang, Z; Zhang, Q; Wang, J; Zhang, H; Niu, X; Li, P. Preparation of the different derivatives of the low-molecular-weight porphyran from Porphyra haitanensis and their antioxidant activities in vitro. Int J Biol Macromol 2009, 45, 22–26. [Google Scholar]
- Zhang, HJ; Mao, WJ; Fang, F; Li, HY; Sun, HH; Chen, Y; Qi, XH. Chemical characteristics and anticoagulant activities of a sulfated polysaccharide and its fragments from Monostroma latissimum. Carbohydr Polym 2008, 71, 428–434. [Google Scholar]
- Sekkal, M; Legrand, P. A spectroscopic investigation of the carrageenans and agar in the 1500–100 cm−1 spectral range. Spectrochim Acta A Mol Biomol Spectrosc 1993, 49, 209–221. [Google Scholar]
- Lahaye, M; Rochas, C; Yaphe, W. A new procedure for determining the heterogeneity of agar polymer in the cell walls of Gracilaria spp. (Gracilariaceae, Rhodophyta). Can J Bot 1986, 64, 579–585. [Google Scholar]
- Silva, TM; Alves, LG; de Queiroz, KC; Santos, MG; Marques, CT; Chavante, SF; Rocha, HA; Leite, EL. Partial characterization and anticoagulant activity of a heterofucan from the brown seaweed Padina gymnospora. Braz J Med Biol Res 2005, 38, 523–533. [Google Scholar]
- Mayer, AMS; Rodríguez, AD; Berlinck, GS; Hamann, MT. Marine pharmacology in 2005–6 Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim Biophys Acta 2009, 1790, 283–308. [Google Scholar]
- Chevolot, L; Foucault, A; Chaubet, F; Kervarec, N; Sinquin, C; Fisher, A; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: relationships with anticoagulant activity. Carbohydr Res 1999, 319, 154–165. [Google Scholar]
- Fonseca, RJC; Santos, GRC; Mourão, PAS. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding. Practical and conceptual implications. Thromb Haemost 2009, 102, 829–836. [Google Scholar]
- Burritt, DJ; Larkindale, J; Hurd, CL. Antioxidant metabolism in the intertidal red seaweed Stictosiphonia arbuscula following desiccation. Planta 2002, 215, 829–838. [Google Scholar]
- Aguilera, J; Bischof, K; Karsten, U; Hanelt, D; Wiencke, C. Seasonal variation in ecophysiological patterns in macroalgae from an Arctic fjord. II. Pigment accumulation and biochemical defence systems against high light stress. Mar Biol 2002, 140, 12. [Google Scholar]
- Chandini, SK; Ganesan, P; Bhaskar, N. In vitro antioxidant activities of three selected brown seaweeds of India. Food Chem 2008, 107, 707–713. [Google Scholar]
- Qi, H; Zhang, Q; Zhaoa, T; Chenc, R; Zhang, H; Niu, X; Li, Z. Antioxidant activity of different sulfate content derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta) in vitro. Int J Biol Macromol 2005, 37, 195–199. [Google Scholar]
- Valko, M; Leibfritz, D; Moncol, J; Cronin, MTD; Mazur, M; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 2007, 39, 44–84. [Google Scholar]
- Wang, J; Zhang, Q; Zhang, Z; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int J Biol Macromol 2008, 42, 127–132. [Google Scholar]
- Niki, E. Antioxidants and atherosclerosis. Biochem Soc Trans 2004, 32, 156–159. [Google Scholar]
- Dubois, M; Gilles, KA; Hamilton, JK; Rebers, PA; Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem 1956, 28, 250–256. [Google Scholar]
- Dodgson, KS; Price, RG. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem J 1962, 84, 106–110. [Google Scholar]
- Spector, J. Refinement of the coomassie blue method of protein quantification. A simple and linear spectrophotometric assay of 0.5 to 50 μg of protein. Anal Biochem 1978, 86, 142–143. [Google Scholar]
- Somogyi, M. Notes on sugar determination. J Biol Chem 1952, 195, 19–23. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Sulfated Polysaccharides | Yield a (%) | Total sugar (%) | Sulfate (%) | S/TSU ratio (%/%) | Protein (%) | Molar Ratio | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fuc | Gluc Acid | Gal | Xyl | Man | Gluc | ||||||
| CC-0.3 | 14.8 | 12.2 | 2.8 | 0.2 | 0.6 | 1.0 | 2.0 | 0.5 | 0.5 | 0.5 | 0.5 |
| CC-0.5 | 12.1 | 33.4 | 3.9 | 0.1 | 0.5 | 1.0 | 2.0 | - | 0.5 | - | - |
| CC-0.7 | 21.3 | 33.2 | 19.2 | 0.6 | 0.4 | 1.0 | 1.5 | - | 0.5 | - | n.d. |
| CC-1.0 | 17.7 | 41.8 | 16.5 | 0.4 | 0.3 | 1.0 | 1.0 | 0.5 | 0.5 | - | n.d. |
| CC-1.2 | 10.6 | 21.8 | 7.8 | 0.4 | 0.2 | 1.0 | 1.0 | 2.5 | 0.5 | - | n.d. |
| CC-2.0 | 23.5 | 19.9 | 20.1 | 1.0 | 0.2 | 1.0 | 1.0 | 2.0 | 0.5 | 0.5 | n.d. |
| Sulfated Polysaccharides | IR (KBr) (cm−1) |
|---|---|
| CC-0.3 | 3441, 1646, 1247, 1037, 820 |
| CC-0.5 | 3428, 1650, 1233, 1071, 850 |
| CC-0.7 | 3434, 1642, 1247, 1064, 833 |
| CC-1.0 | 3423, 1644, 1231, 1065, 841 |
| CC-1.2 | 3431, 1641, 1239, 1042, 827 |
| CC-2.0 | 3443, 1649, 1251, 1056, 839 |
| Sulfated polysaccharides | Concentration (mg/mL) | Scavenging (%) | |
|---|---|---|---|
| OH• | O2− | ||
| CC-0.3 | 0.010 | 0.0 ± 0.0 a | 3.9 ± 1.4 a |
| 0.025 | 0.0 ± 0.0 a | 18.1 ± 1.3 b | |
| 0.050 | 0.9 ± 0.1 b | 18.0 ± 0.9 b | |
| 0.100 | 1.0 ± 0.2 b | 18.0 ± 1.8 b | |
| CC-0.5 | 0.010 | 0.0 ± 0.0 a | 3.4 ± 0.6 a |
| 0.025 | 0.0 ± 0.0 a | 8.3 ± 1.2 b | |
| 0.050 | 0.1 ± 0.1 a | 13.3 ± 0.9 c | |
| 0.100 | 0.5 ± 0.1 b | 13.0 ± 1.8 c | |
| CC-0.7 | 0.010 | 0.0 ± 0.0 a | 3.1 ± 1.3 a |
| 0.025 | 0.0 ± 0.0 a | 18.1 ± 1.3 b | |
| 0.050 | 0.9 ± 0.2 b | 18.2 ± 6.1 b | |
| 0.100 | 0.9 ± 0.0 b | 18.4 ± 1.2 b | |
| CC-1.0 | 0.010 | 0.0 ± 0.0 a | 0.8 ± 0.5 a |
| 0.025 | 0.0 ± 0.0 a | 0.7 ± 1.4 a | |
| 0.050 | 2.9 ± 0.2 b | 0.8 ± 1.7 a | |
| 0.100 | 3.0 ± 0.1 b | 0.3 ± 2.1 a | |
| CC-1.2 | 0.010 | 0.0 ± 0.0 a | 9.2 ± 1.0 a |
| 0.025 | 0.0 ± 0.0 a | 14.1 ± 1.9 b | |
| 0.050 | 2.9 ± 0.1 b | 24.2 ± 3.7 c | |
| 0.100 | 3.0 ± 0.1 b | 43.1 ± 2.1 d | |
| CC-2.0 | 0.010 | 0.0 ± 0.0 a | 3.4 ± 0.7 a |
| 0.025 | 0.0 ± 0.0 a | 6.0 ± 1.4 b | |
| 0.050 | 2.4 ± 0.2 b | 13.4 ± 0.8 c | |
| 0.100 | 2.9 ± 0.1 b | 13.1 ± 1.7 c | |
| Gallic acid | 0.100 | 43.6 ± 2.4 | 41.8 ± 4.7 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barros Gomes Camara, R.; Silva Costa, L.; Pereira Fidelis, G.; Duarte Barreto Nobre, L.T.; Dantas-Santos, N.; Lima Cordeiro, S.; Santana Santos Pereira Costa, M.; Guimaraes Alves, L.; Oliveira Rocha, H.A. Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities. Mar. Drugs 2011, 9, 124-138. https://doi.org/10.3390/md9010124
Barros Gomes Camara R, Silva Costa L, Pereira Fidelis G, Duarte Barreto Nobre LT, Dantas-Santos N, Lima Cordeiro S, Santana Santos Pereira Costa M, Guimaraes Alves L, Oliveira Rocha HA. Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities. Marine Drugs. 2011; 9(1):124-138. https://doi.org/10.3390/md9010124
Chicago/Turabian StyleBarros Gomes Camara, Rafael, Leandro Silva Costa, Gabriel Pereira Fidelis, Leonardo Thiago Duarte Barreto Nobre, Nednaldo Dantas-Santos, Sara Lima Cordeiro, Mariana Santana Santos Pereira Costa, Luciana Guimaraes Alves, and Hugo Alexandre Oliveira Rocha. 2011. "Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities" Marine Drugs 9, no. 1: 124-138. https://doi.org/10.3390/md9010124
APA StyleBarros Gomes Camara, R., Silva Costa, L., Pereira Fidelis, G., Duarte Barreto Nobre, L. T., Dantas-Santos, N., Lima Cordeiro, S., Santana Santos Pereira Costa, M., Guimaraes Alves, L., & Oliveira Rocha, H. A. (2011). Heterofucans from the Brown Seaweed Canistrocarpus cervicornis with Anticoagulant and Antioxidant Activities. Marine Drugs, 9(1), 124-138. https://doi.org/10.3390/md9010124