Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes

Abstract
:1. Introduction
2. Results
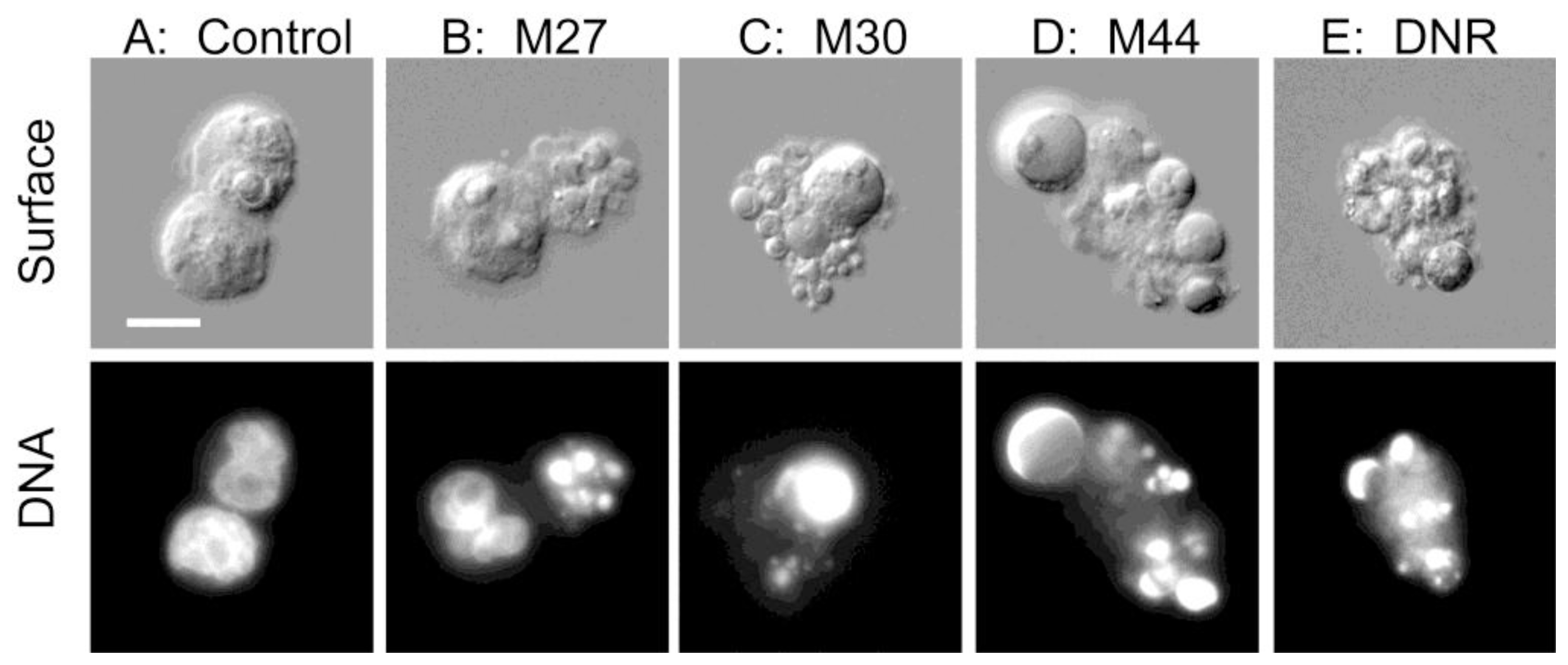
2.1. Screening of benthic marine cyanobacteria for leukemia cell death inducers
2.2. Modulation of blood platelet activity by cyanobacterial extracts
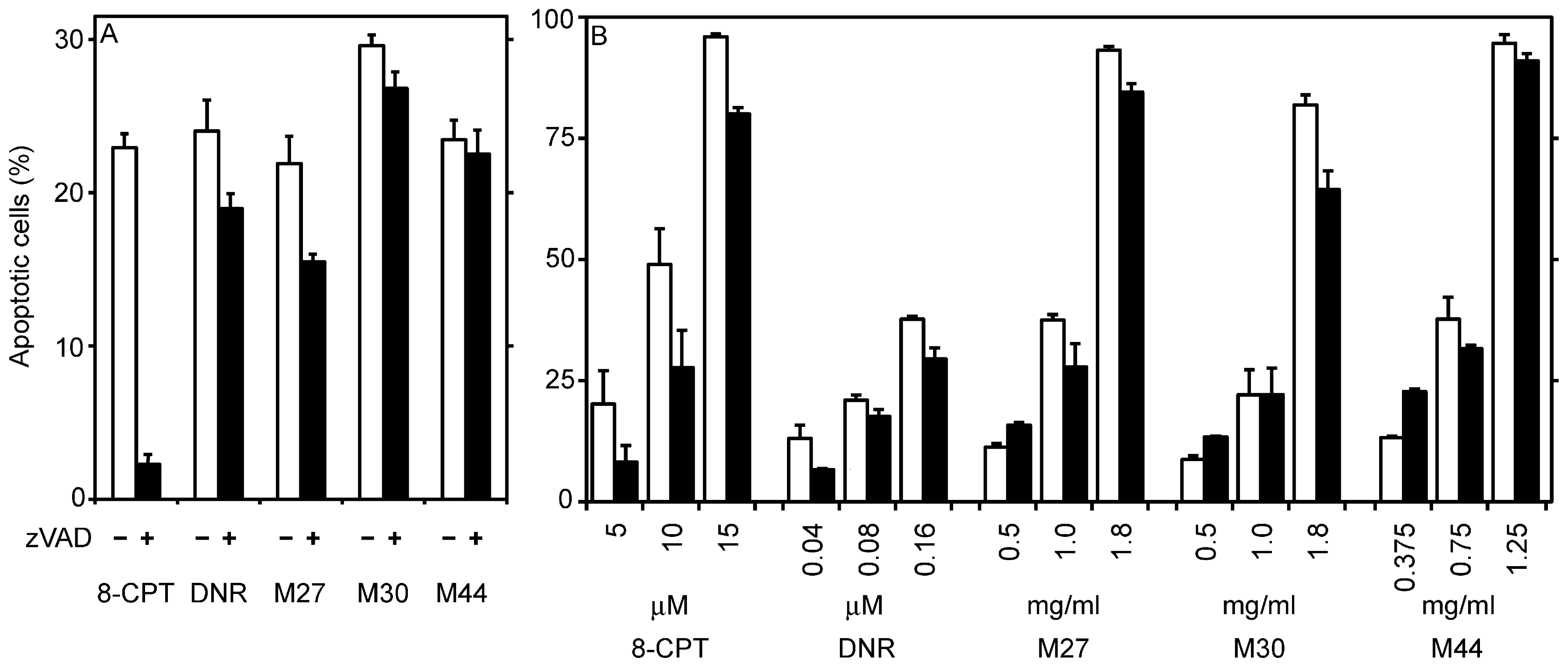
2.3. Drug-relevant features of the apoptogens in cyanobacteria M27, M30 and M44
3. Discussion
4. Conclusions
5. Experimental Section
5.1. Materials
5.2. Isolation and cultivation of cyanobacterial strains
5.3. Sequential extraction of cyanobacterial biomass
5.4. Maintenance of cell lines, experimental conditions and determination of cell death
5.5. Isolation, incubation and flow cytometry analysis of human blood platelets
Supplementary material
Table S1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code number | Genus |
|---|---|
| 1 | Nostoc sp. |
| 2 | Anabaena sp. |
| 3 | Anabaena sp |
| 4 | Anabaena sp |
| 5 | Calothrix sp |
| 6 | Nostoc sp. |
| 7 | Nostoc sp. |
| 8 | Anabaena sp. |
| 9 | Anabaena sp. |
| 11 | Anabaena sp. |
| 12 | Anabaena sp. |
| 13 | Anabaena sp. |
| 14 | Anabaena sp. |
| 15 | Calothrix sp |
| 16 | Calothrix sp |
| 18 | Calothrix sp |
| 19 | Calothrix sp |
| 20 | Anabaena sp. |
| 21 | Calothrix sp |
| 22 | Nostoc sp. |
| 23 | Anabaena sp. |
| 25 | Anabaena sp |
| 26 | Anabaena sp |
| 27 | Anabaena sp |
| 28 | Nostoc sp |
| 29 | Anabaena sp |
| 30 | Anabaena sp. |
| 31 | Anabaena sp |
| 33 | Anabaena sp |
| 34 | Cyanothece sp. |
| 35 | Unidentified |
| 36 | Nostoc sp. |
| 37 | Anabaena sp. |
| 38 | Cyanothece sp. |
| 39 | Anabaena sp |
| 41 | Nostoc sp. |
| 44 | Anabaena sp |
| 45 | Anabaena sp |
| 46 | Anabaena sp |
| 47 | Calothrix sp |
| 48 | Anabaena sp |
Bioguided Partial Purification of the Apoptogenic Constituent in the Aqueous M44-Extract
Solid phase extraction
Reversed phase chromatography

Table S2
| Bioactive fraction | Recovery (%) |
|---|---|
| Water extract supernatant | 100 |
| Water-methanol phase | 96 ± 4.2 |
| SPE flow through fraction | 88 ± 2.5 |
| RP-HPLC fraction (2.2–3.8min) | 58 ± 2.2 |
Acknowledgements
- Samples Availability: Available from the authors.
References
- Burja, AM; Banaigs, B; Abou-Mansour, E; Burgess, JG; Wright, PC. Marine cyanobacteria-a prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar]
- Tan, LT. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar]
- Simmons, TL; Andrianasolo, E; McPhail, K; Flatt, P; Gerwick, WH. Marine natural products as anticancer drugs. Mol. Cancer Ther 2005, 4, 333–342. [Google Scholar]
- Mayer, AM; Glaser, KB; Cuevas, C; Jacobs, RS; Kem, W; Little, RD; McIntosh, JM; Newman, DJ; Potts, BC; Shuster, DE. The odyssey of marine pharmaceuticals: a current pipeline perspective. Trends Pharmacol. Sci 2010, 31, 255–265. [Google Scholar]
- Newman, DJ; Cragg, GM. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod 2004, 67, 1216–1238. [Google Scholar]
- Liu, L; Rein, KS. New peptides isolated from Lyngbya species: a review. Mar. Drugs 2010, 8, 1817–1837. [Google Scholar]
- Jemal, A; Siegel, R; Ward, E; Hao, Y; Xu, J; Thun, MJ. Cancer statistics, 2009. CA Cancer J. Clin 2009, 59, 225–249. [Google Scholar]
- Joel, SP; Rohatiner, A. Henderson, ED, Lister, TA, Greaves, MF, Eds.; Pharmacology of antileukemic drugs. In Leukemia, 7th ed; Saunders: Philadelphia, PA, USA, 2002; pp. 394–440. [Google Scholar]
- Lowenberg, B; Burnett, AK. Degos, L, Linch, DC, Lowenberg, B, Eds.; Acute myeloid leukemia in adults. In Textbook of Malignant Hematology, 2nd ed; Taylor & Francis: New York, NY, USA, 2005; pp. 633–656. [Google Scholar]
- Stapnes, C; Gjertsen, BT; Reikvam, H; Bruserud, Ø. Targeted therapy in acute myeloid leukaemia: current status and future directions. Expert Opin. Investig. Drugs 2009, 18, 433–455. [Google Scholar]
- Huang, TS; Myklebust, LM; Kjarland, E; Gjertsen, BT; Pendino, F; Bruserud, O; D⊘skeland, SO; Lillehaug, JR. LEDGF/p75 has increased expression in blasts from chemotherapy-resistant human acute myelogenic leukemia patients and protects leukemia cells from apoptosis in vitro. Mol. Cancer 2007, 6, 31. [Google Scholar]
- Ahuja, HG; Hong, J; Aplan, PD; Tcheurekdjian, L; Forman, SJ; Slovak, ML. t(9;11)(p22;p15) in acute myeloid leukemia results in a fusion between NUP98 and the gene encoding transcriptional coactivators p52 and p75-lens epithelium-derived growth factor (LEDGF). Cancer Res 2000, 60, 6227–6229. [Google Scholar]
- Daugaard, M; Kirkegaard-Sorensen, T; Ostenfeld, MS; Aaboe, M; Hoyer-Hansen, M; Orntoft, TF; Rohde, M; Jaattela, M. Lens epithelium-derived growth factor is an Hsp70–2 regulated guardian of lysosomal stability in human cancer. Cancer Res 2007, 67, 2559–2567. [Google Scholar]
- Campos, L; Rouault, JP; Sabido, O; Oriol, P; Roubi, N; Vasselon, C; Archimbaud, E; Magaud, JP; Guyotat, D. High expression of bcl-2 protein in acute myeloid leukemia cells is associated with poor response to chemotherapy. Blood 1993, 81, 3091–3096. [Google Scholar]
- Reed, JC. Bcl-2-family proteins and hematologic malignancies: history and future prospects. Blood 2008, 111, 3322–3330. [Google Scholar]
- Minotti, G; Menna, P; Salvatorelli, E; Cairo, G; Gianni, L. Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev 2004, 56, 185–229. [Google Scholar]
- Herfindal, L; Oftedal, L; Selheim, F; Wahlsten, M; Sivonen, K; D⊘skeland, SO. A high proportion of Baltic Sea benthic cyanobacterial isolates contain apoptogens able to induce rapid death of isolated rat hepatocytes. Toxicon 2005, 46, 252–260. [Google Scholar]
- Selheim, F; Herfindal, L; Martins, R; Vasconçelos, V; D⊘skeland, SO. Neuro-apoptogenic and blood platelet targeting toxins in benthic marine cyanobacteria from the Portuguese coast. Aquat. Toxicol 2005, 74, 294–306. [Google Scholar]
- Martins, RF; Ramos, MF; Herfindal, L; Sousa, JA; Skaerven, K; Vasconçelos, VM. Antimicrobial and cytotoxic assessment of marine cyanobacteria-Synechocystis and Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar]
- Oftedal, L; Skjaerven, KH; Coyne, RT; Edvardsen, B; Rohrlack, T; Skulberg, OM; D⊘skeland, SO; Herfindal, L. The apoptosis-inducing activity towards leukemia and lymphoma cells in a cyanobacterial culture collection is not associated with mouse bioassay toxicity. J. Ind. Microbiol. Biotechnol 2010. [Google Scholar] [CrossRef]
- Surakka, A; Sihvonen, LM; Lehtimäki, JM; Wahlsten, M; Vuorela, P; Sivonen, K. Benthic cyanobacteria from the Baltic Sea contain cytotoxic Anabaena, Nodularia, and Nostoc strains and an apoptosis-inducing Phormidium strain. Environ. Toxicol 2005, 20, 285–292. [Google Scholar]
- Lacaze, N; Gombaud-Saintonge, G; Lanotte, M. Conditions controlling long-term proliferation of Brown Norway rat promyelocytic leukemia in vitro: primary growth stimulation by microenvironment and establishment of an autonomous Brown Norway 'leukemic stem cell line'. Leuk. Res 1983, 7, 145–154. [Google Scholar]
- McCormack, E; Bruserud, Ø; Gjertsen, BT. Animal models of acute myelogenous leukaemia - development, application and future perspectives. Leukemia 2005, 19, 687–706. [Google Scholar]
- Barbui, T; Finazzi, G; Falanga, A. Henderson, ES, Lister, TA, Greaves, MF, Eds.; Management of bleeding and thrombosis in acute leukemia and chronic myeloproliferative disorders. In Leukemia, 7th ed; Saunders: Philadelphia, PA, USA, 2002; pp. 363–382. [Google Scholar]
- Degos, L. Degos, L, Finch, DC, Lovenberg, B, Eds.; Acute promyelocytic leukemia. In Textbook of Malignant Hematology; Taylor & Francis: London, UK, 2005; pp. 657–668. [Google Scholar]
- Foss, B; Bruserud, Ø. Platelet functions and clinical effects in acute myelogenous leukemia. Thromb. Haemost 2008, 99, 27–37. [Google Scholar]
- Gausdal, G; Gjertsen, BT; McCormack, E; Van Damme, P; Hovland, R; Krakstad, C; Bruserud, Ø; Gevaert, K; Vandekerckhove, J; D⊘skeland, SO. Abolition of stress-induced protein synthesis sensitizes leukemia cells to anthracycline-induced death. Blood 2008, 111, 2866–2877. [Google Scholar]
- Fulda, S; Galluzzi, L; Kroemer, G. Targeting mitochondria for cancer therapy. Nat. Rev. Drug Discov 2010, 9, 447–464. [Google Scholar]
- Seite, P; Ruchaud, S; Hillion, J; Gendron, MC; Bruland, O; Segal-Bendirdjian, E; D⊘skeland, SO; Lillehaug, JR; Lanotte, M. Ectopic expression of Bcl-2 switches over nuclear signalling for cAMP-induced apoptosis to granulocytic differentiation. Cell Death Differ 2000, 7, 1081–1089. [Google Scholar]
- Kroemer, G; Galluzzi, L; Vandenabeele, P; Abrams, J; Alnemri, ES; Baehrecke, EH; Blagosklonny, MV; El-Deiry, WS; Golstein, P; Green, DR; Hengartner, M; Knight, RA; Kumar, S; Lipton, SA; Malorni, W; Nunez, G; Peter, ME; Tschopp, J; Yuan, J; Piacentini, M; Zhivotovsky, B; Melino, G. Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ 2009, 16, 3–11. [Google Scholar]
- Hanahan, D; Weinberg, RA. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar]
- Lipinski, CA; Lombardo, F; Dominy, BW; Feeney, PJ. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev 1997, 23, 3–25. [Google Scholar]
- Vieth, M; Siegel, MG; Higgs, RE; Watson, IA; Robertson, DH; Savin, KA; Durst, GL; Hipskind, PA. Characteristic physical properties and structural fragments of marketed oral drugs. J. Med. Chem 2004, 47, 224–232. [Google Scholar]
- Marston, A; Hostettmann, K. Developments in the application of counter-current chromatography to plant analysis. J. Chromatogr. A 2006, 1112, 181–194. [Google Scholar]
- Sutherland, IA. Recent progress on the industrial scale-up of counter-current chromatography. J. Chromatogr. A 2007, 1151, 6–13. [Google Scholar]
- Castenholz, RW. Garrity, GM, Ed.; Cyanobacteria. In Bergeys Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2001; pp. 473–599. [Google Scholar]
- Prestegard, SK; Oftedal, L; Coyne, RT; Nygaard, G; Skjaerven, KH; Knutsen, G; D⊘skeland, SO; Herfindal, L. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar]
- Tysnes, OB; Aarbakke, GM; Verhoeven, AJ; Holmsen, H. Thin-layer chromatography of polyphosphoinositides from platelet extracts: interference by an unknown phospholipid. Thromb. Res 1985, 40, 329–338. [Google Scholar]
- Selheim, F; Fukami, MH; Holmsen, H; Vassbotn, FS. Platelet-derived-growth-factor-induced signalling in human platelets: phosphoinositide-3-kinase-dependent inhibition of platelet activation. Biochem. J 2000, 350(Pt. 2), 469–475. [Google Scholar]
- Selheim, F; Holmsen, H; Vassbotn, FS. Platelet-derived growth factor inhibits platelet activation in heparinized whole blood. Thromb. Res 1999, 95, 185–196. [Google Scholar]






© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.O.; Herfindal, L. Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes. Mar. Drugs 2010, 8, 2659-2672. https://doi.org/10.3390/md8102659
Oftedal L, Selheim F, Wahlsten M, Sivonen K, Døskeland SO, Herfindal L. Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes. Marine Drugs. 2010; 8(10):2659-2672. https://doi.org/10.3390/md8102659
Chicago/Turabian StyleOftedal, Linn, Frode Selheim, Matti Wahlsten, Kaarina Sivonen, Stein Ove Døskeland, and Lars Herfindal. 2010. "Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes" Marine Drugs 8, no. 10: 2659-2672. https://doi.org/10.3390/md8102659
APA StyleOftedal, L., Selheim, F., Wahlsten, M., Sivonen, K., Døskeland, S. O., & Herfindal, L. (2010). Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes. Marine Drugs, 8(10), 2659-2672. https://doi.org/10.3390/md8102659