Marine Toxins That Target Voltage-gated Sodium Channels

Abstract
:1. Overview
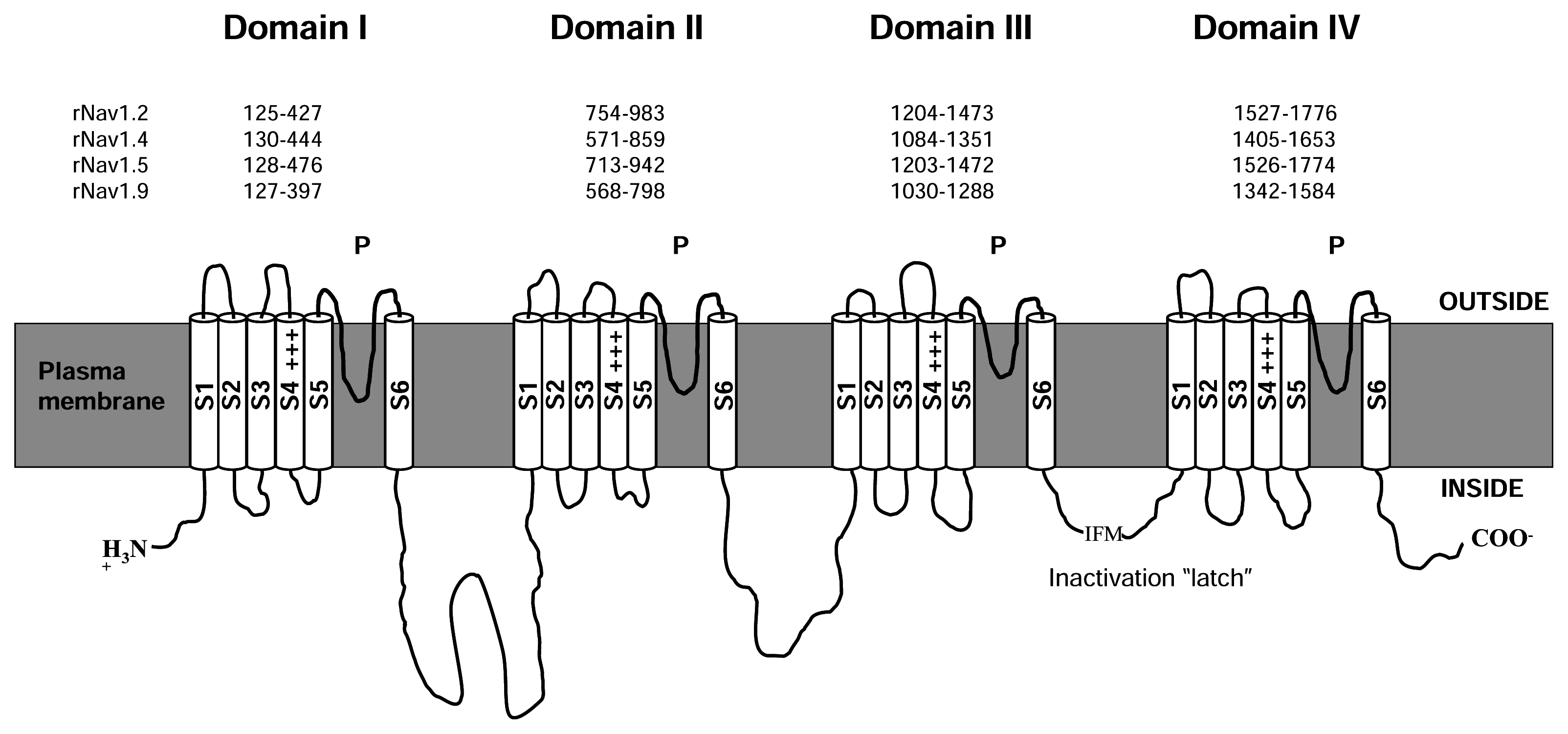
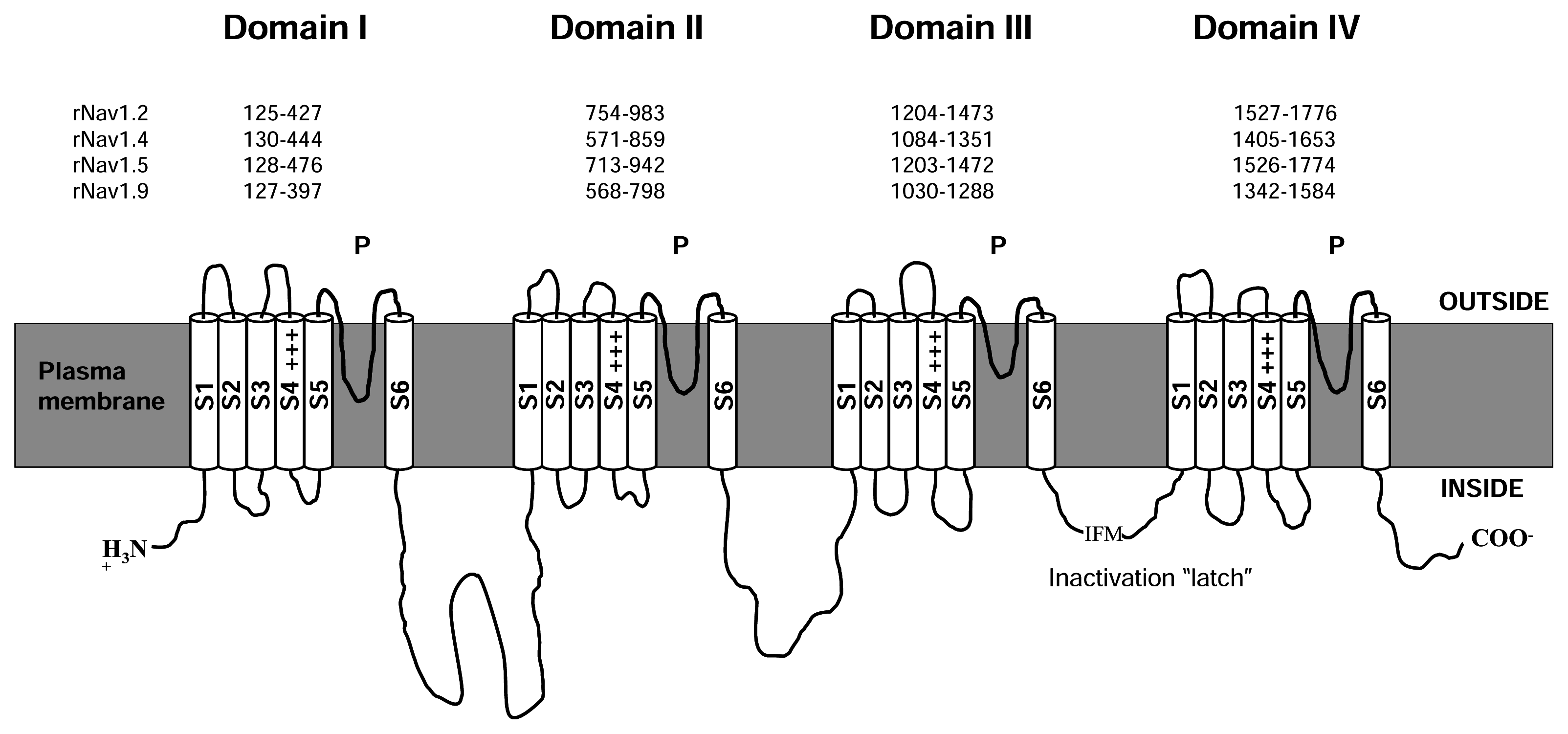
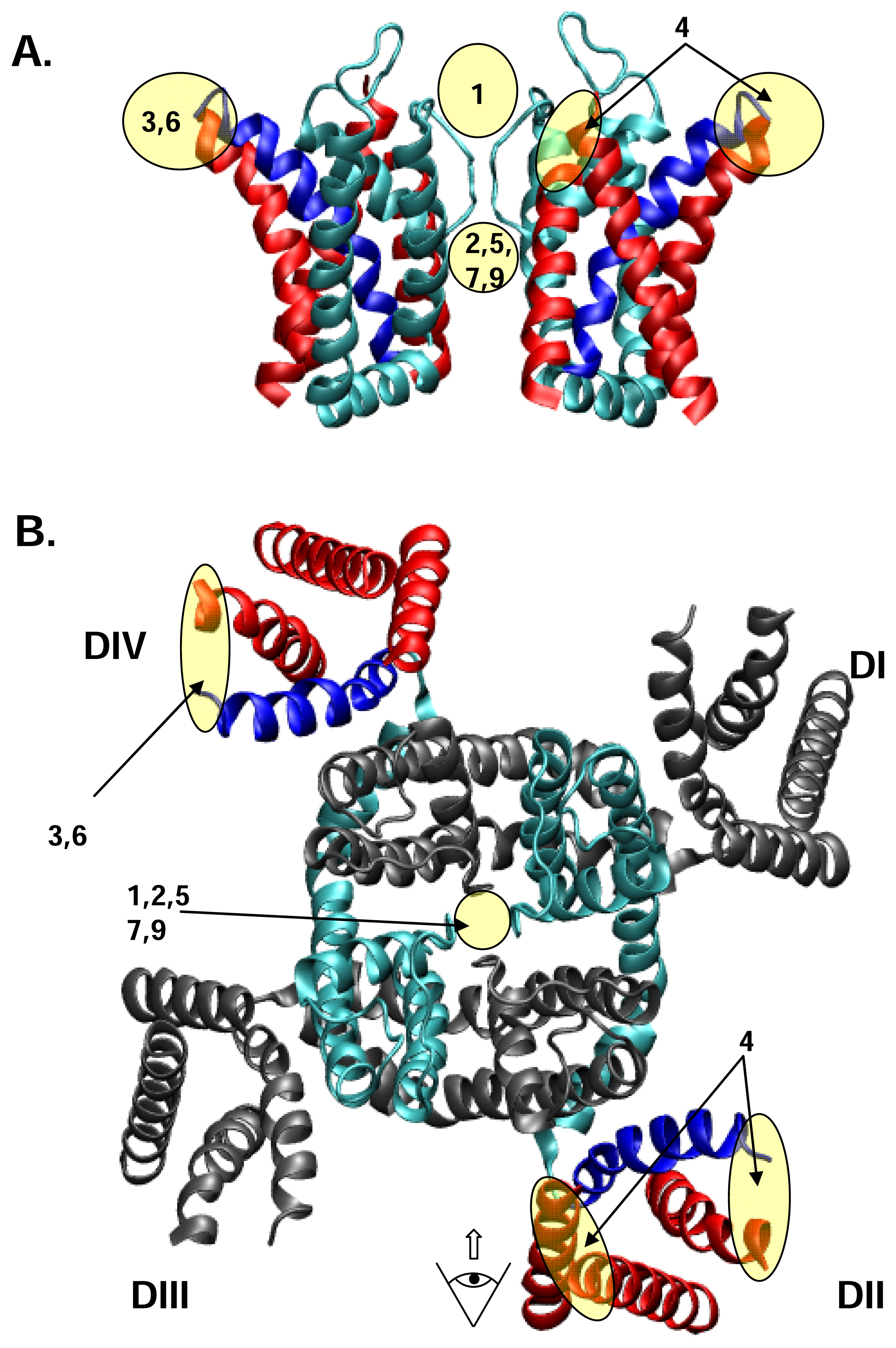
2. Structure of NaV channels
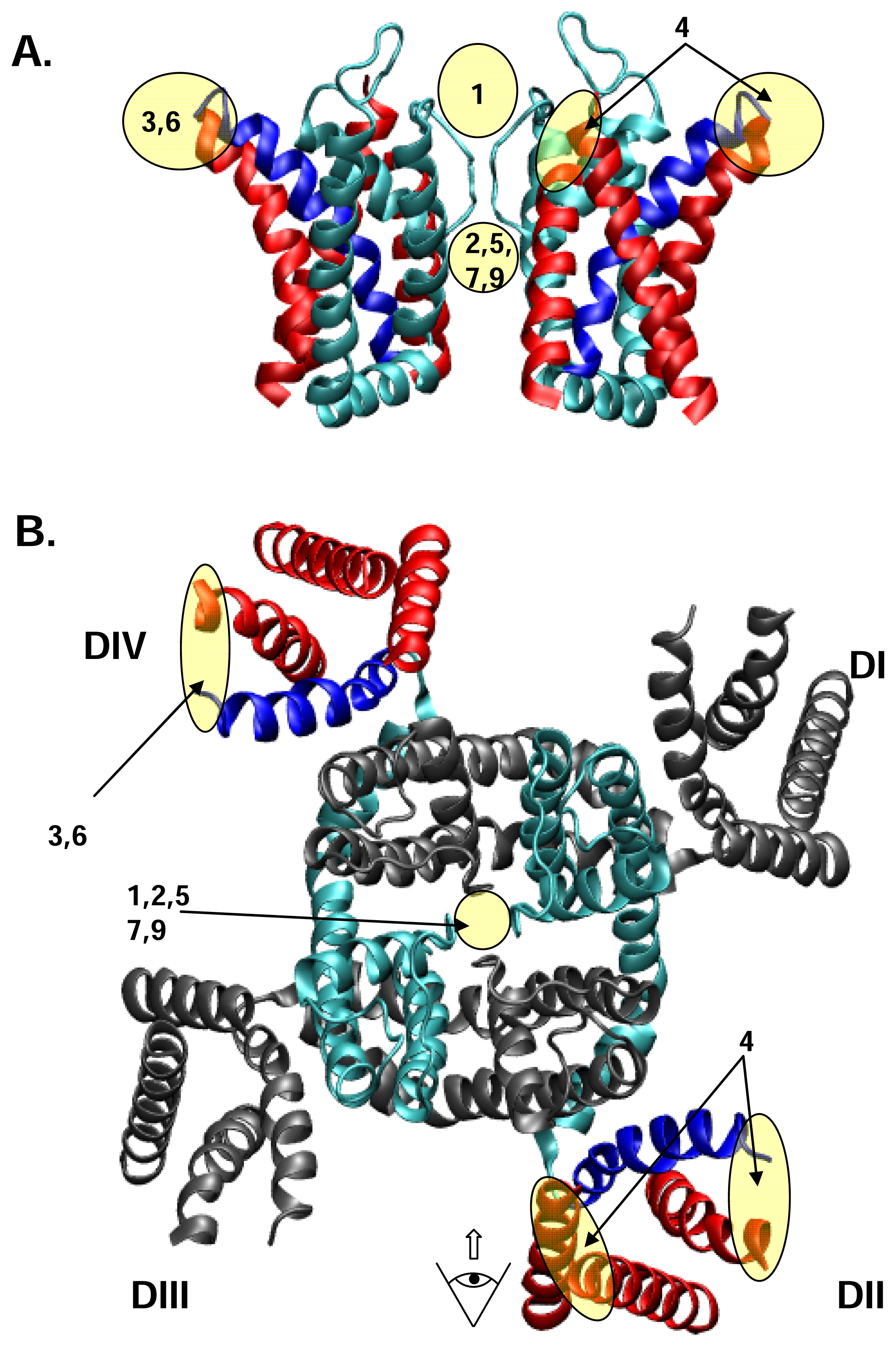
3. Drug and toxin receptors on NaV channels
4. NaV channel blockers and other inhibitors
4.1. Receptor site 1
4.2. Non-peptide NaV channel pore blockers and inhibitors
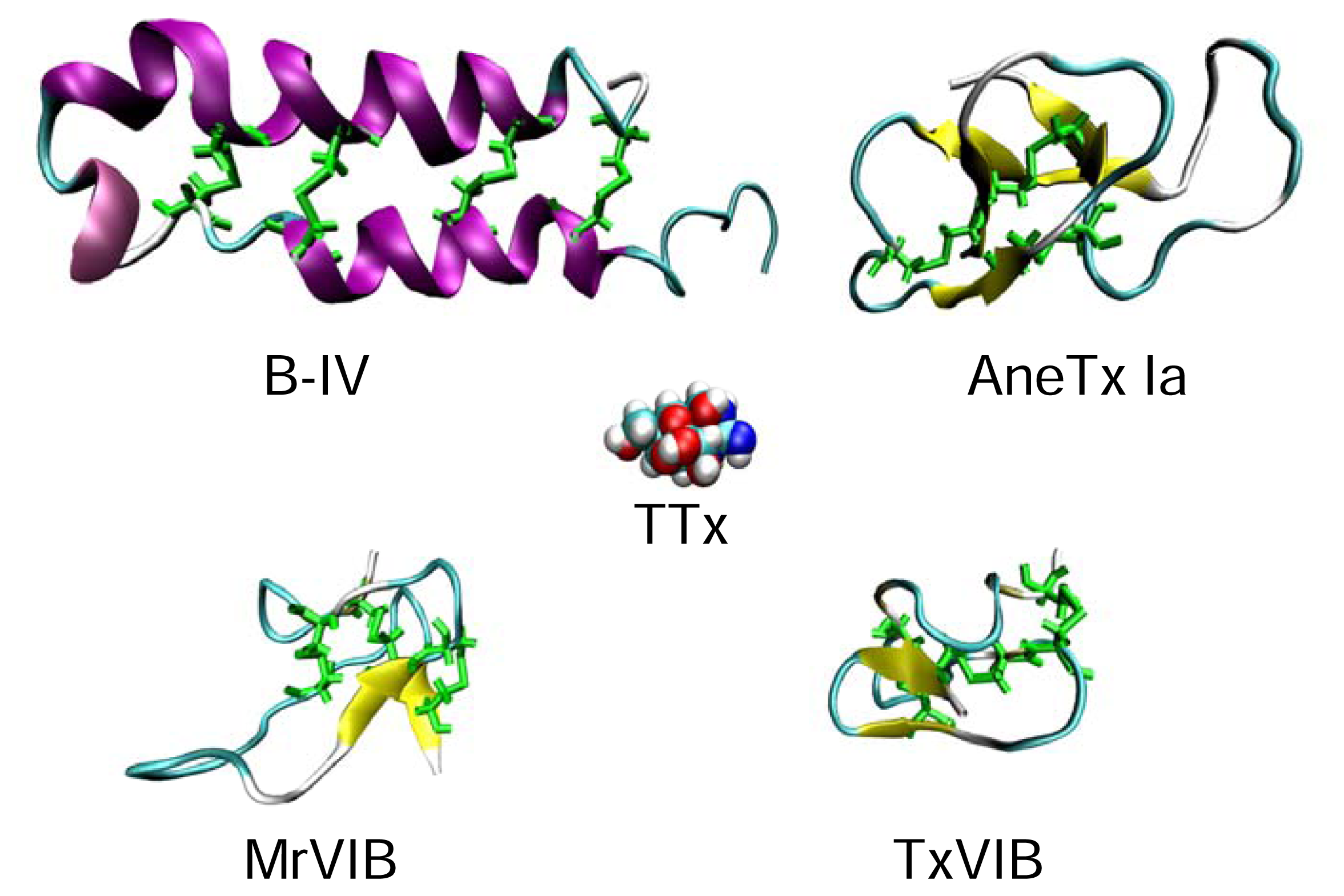
4.2.1. Tetrodotoxin
4.2.2. Saxitoxin
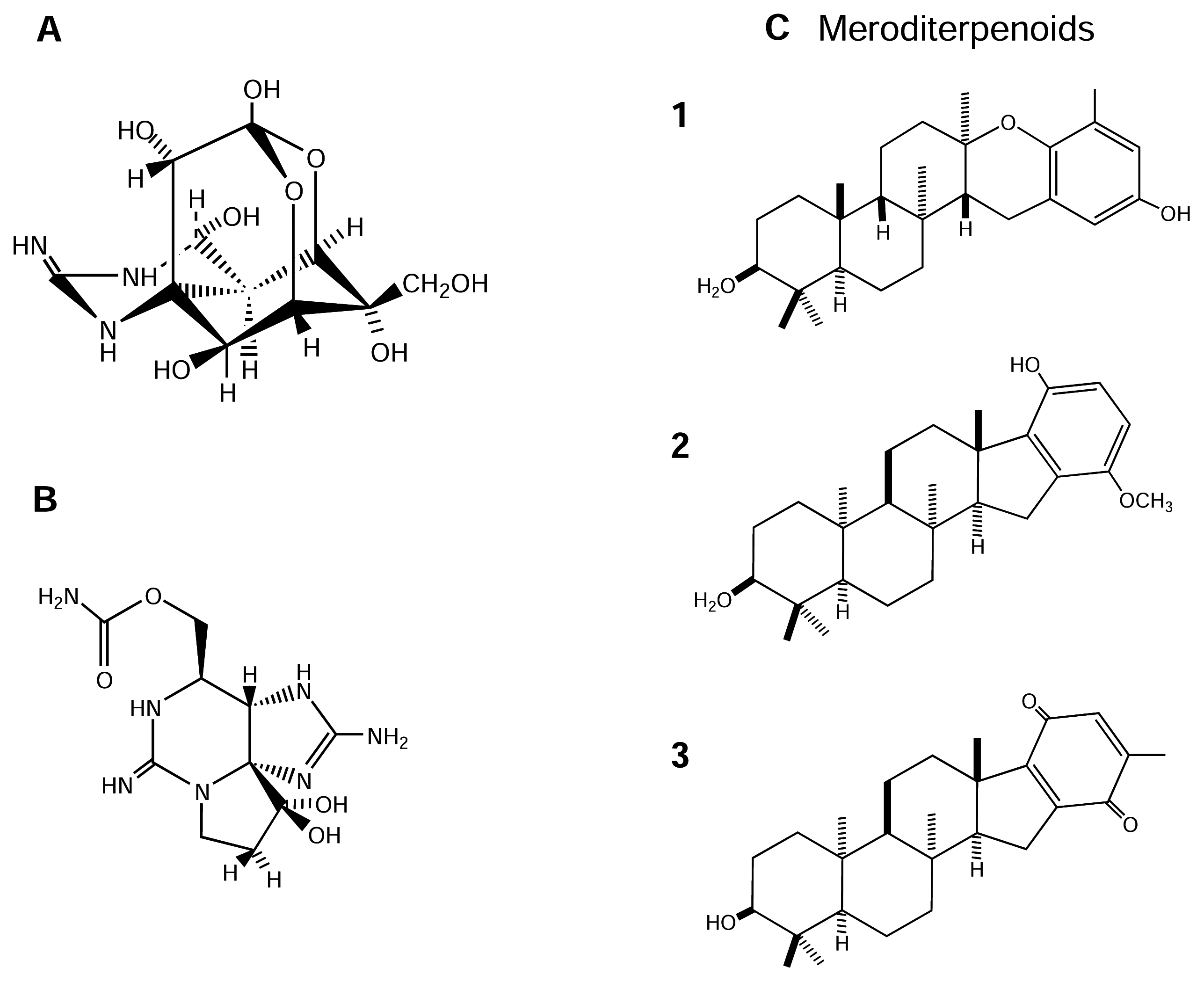
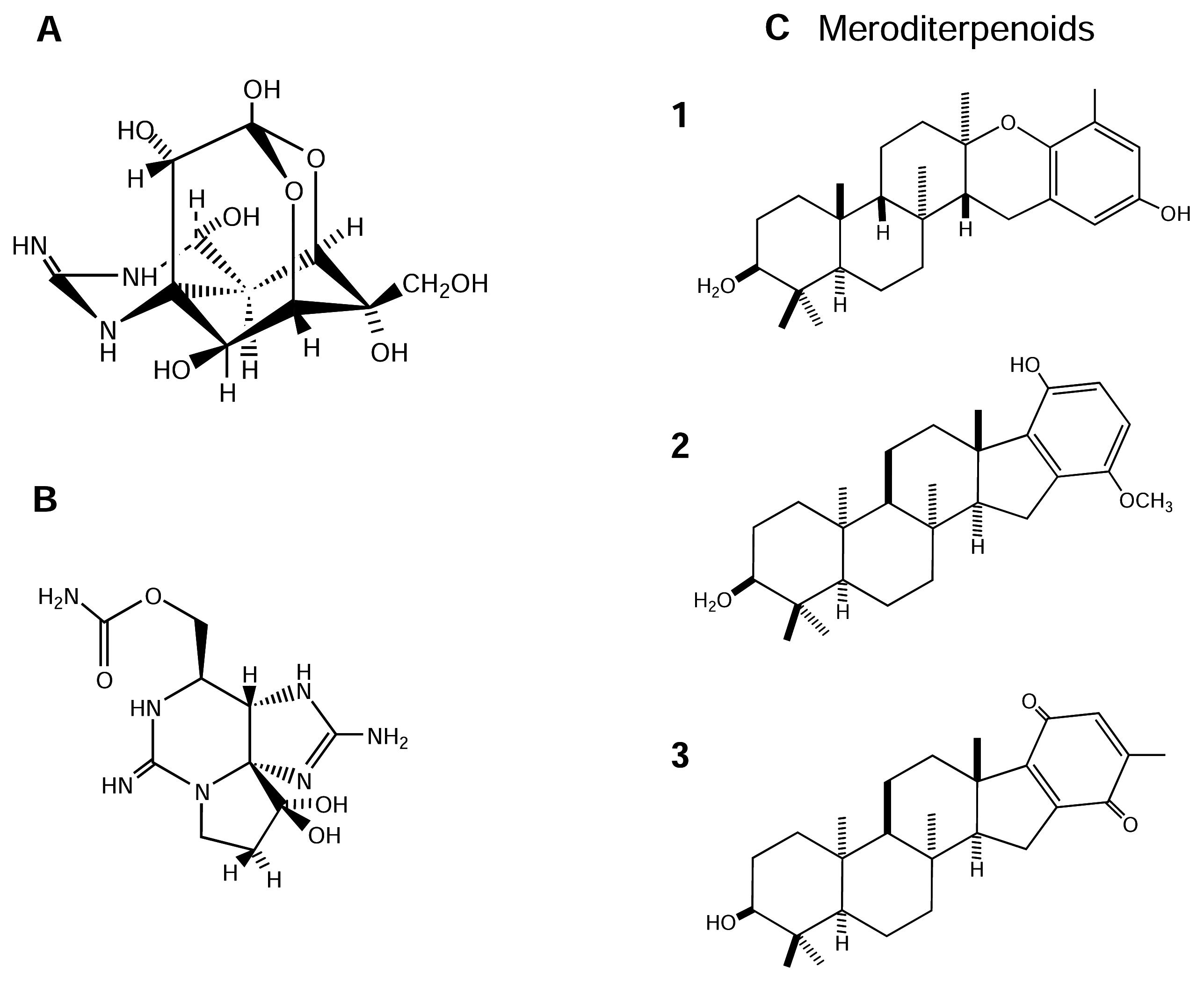
4.2.3. Meroditerpenoids
4.3. Peptide NaV channel pore blockers and other inhibitors
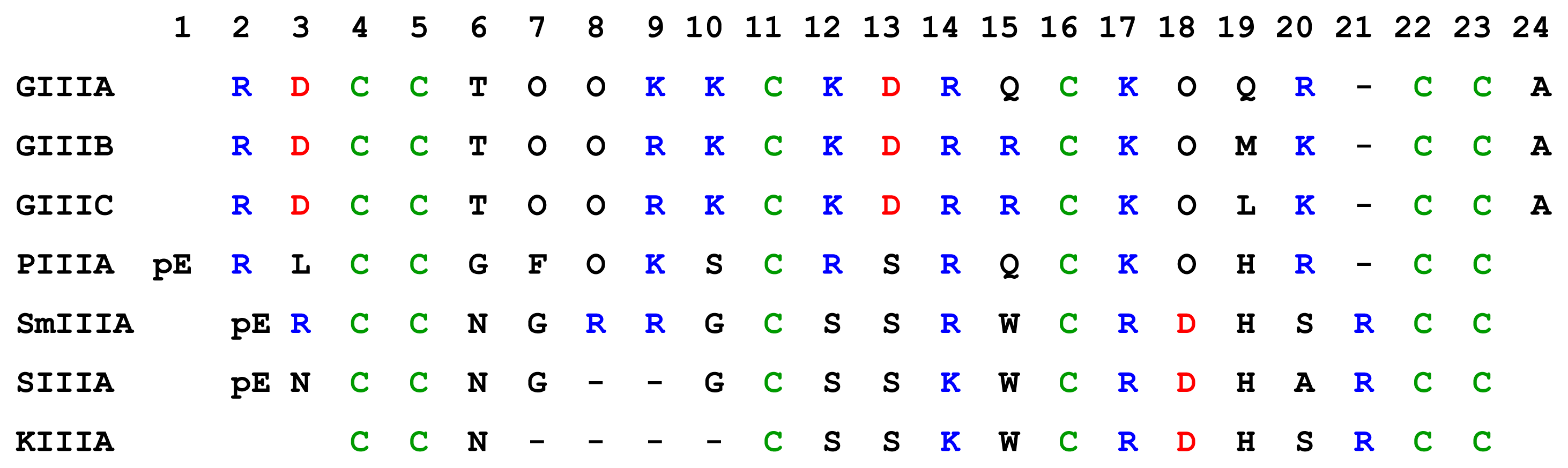
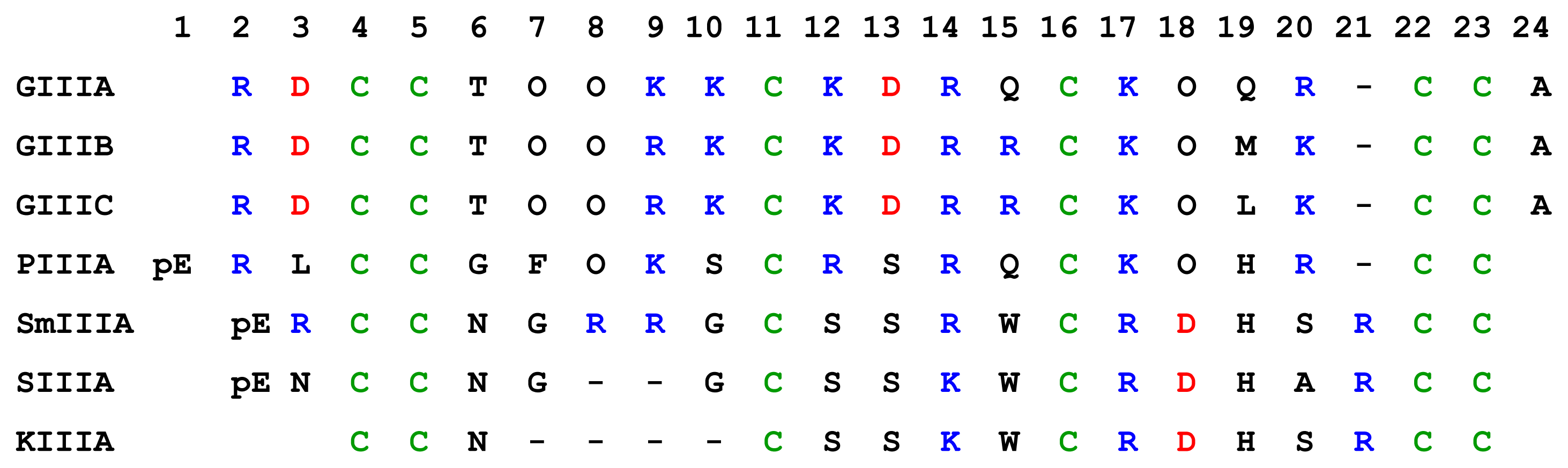
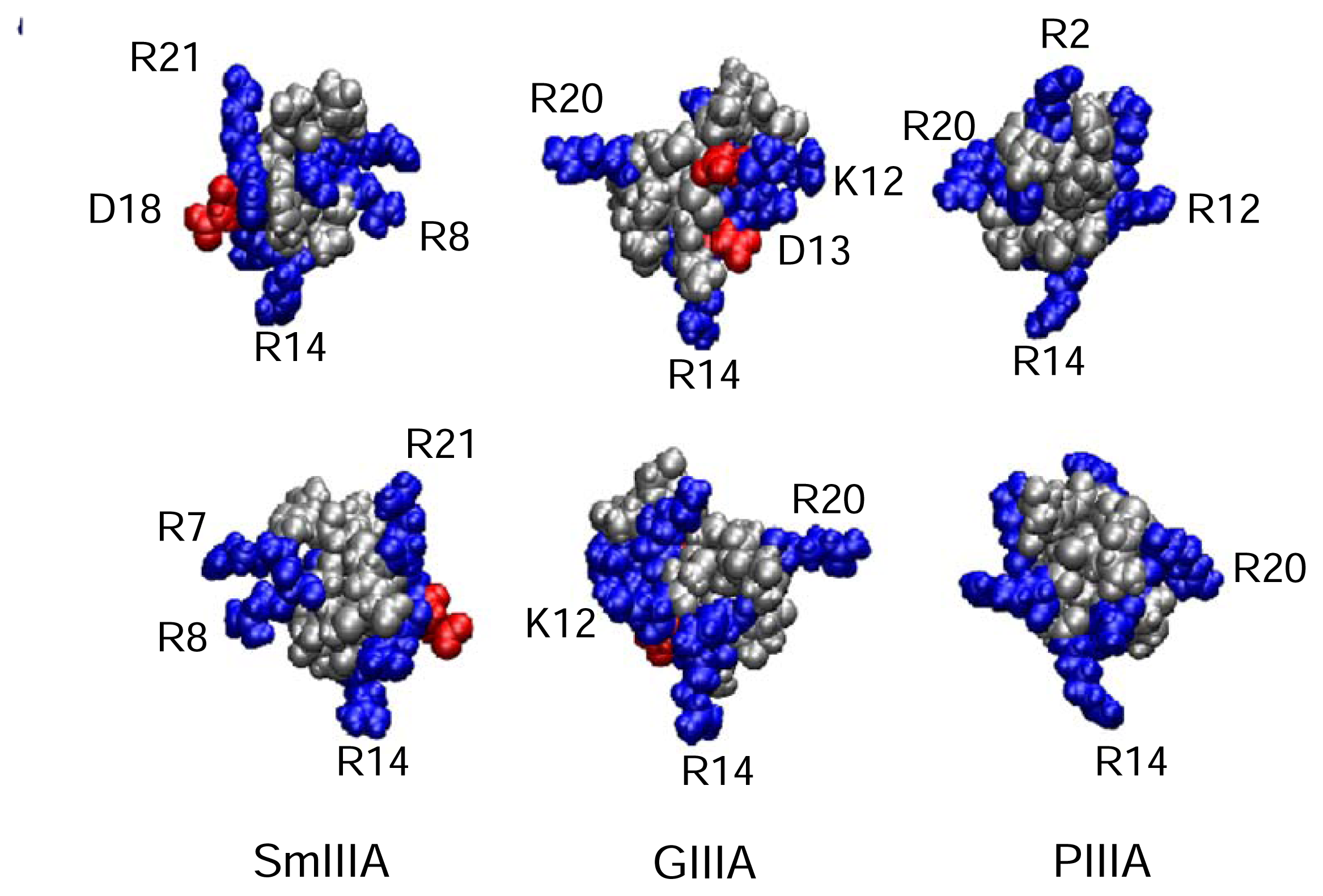
4.3.1. μ-Conotoxins
4.3.2. μO-Conotoxins
4.3.3. Unclassified conotoxins, GS and TVIIA
4.4. Lipopeptide Nav channel inhibitors
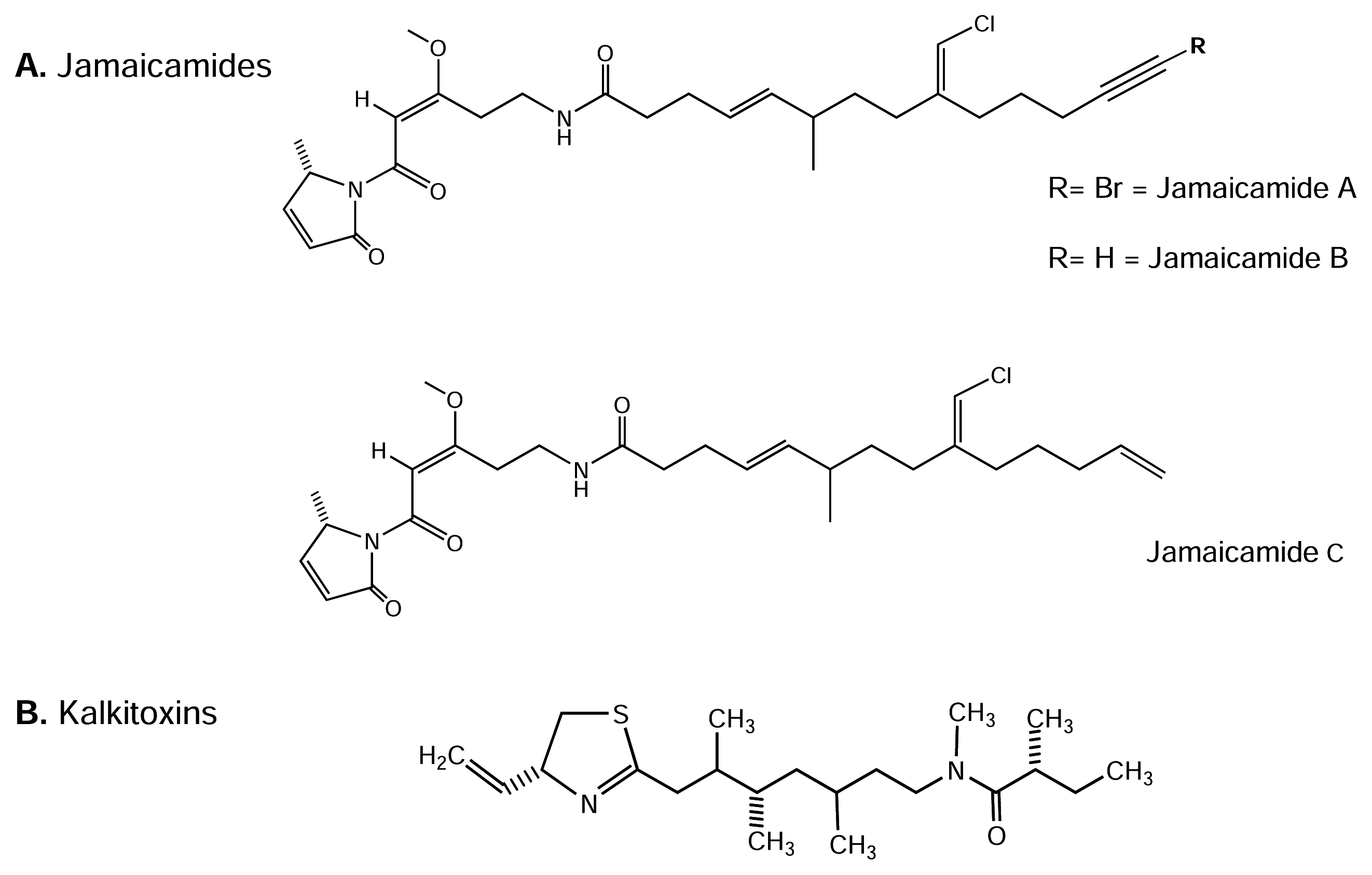
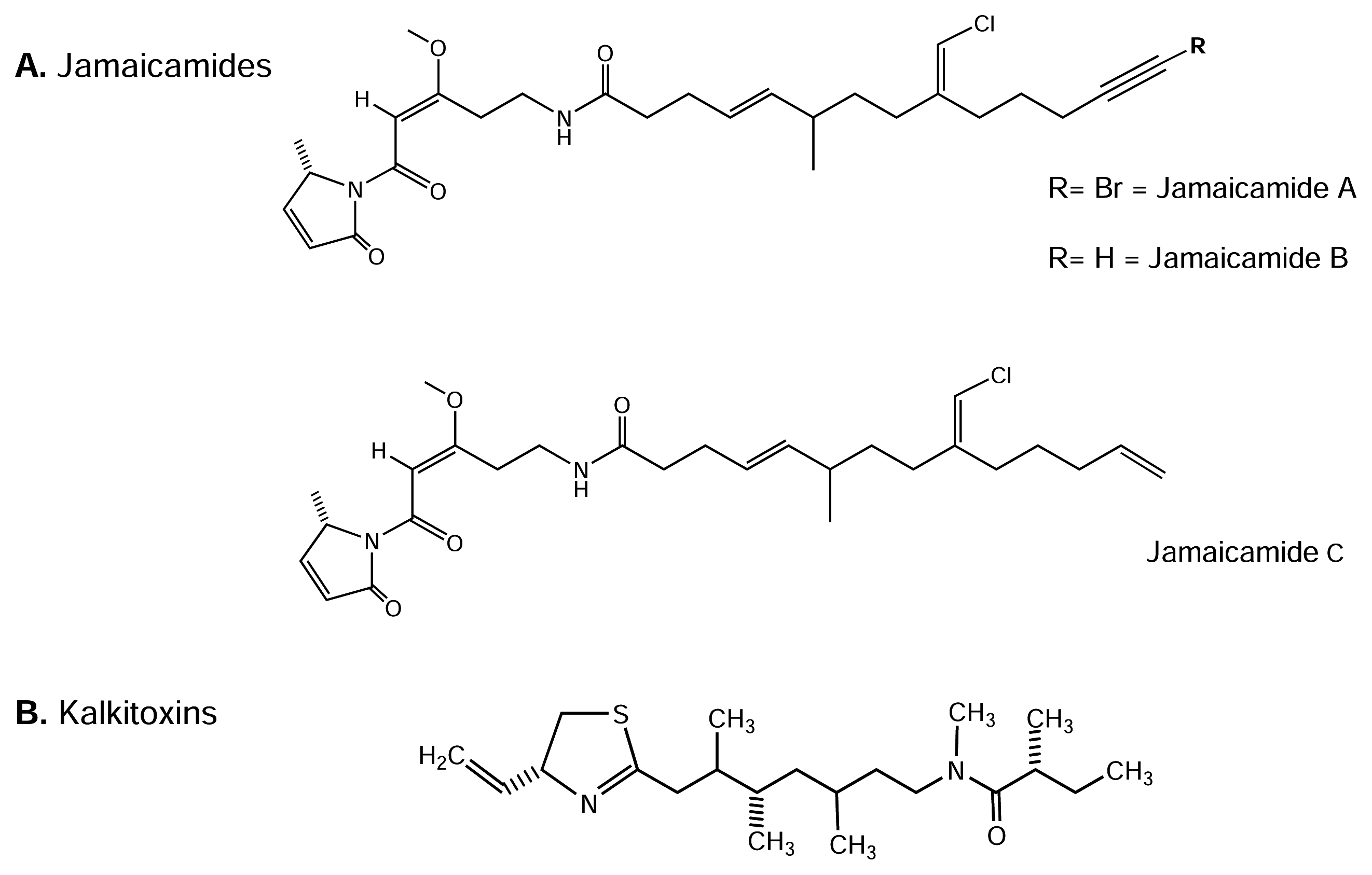
4.4.1. Jamaicamides
4.4.2. Kalkitoxin
5. Gating modifiers
5.1. Non-peptide NaV channel modulators
5.1.1. Alkaloid inhibitors of Na+ channels
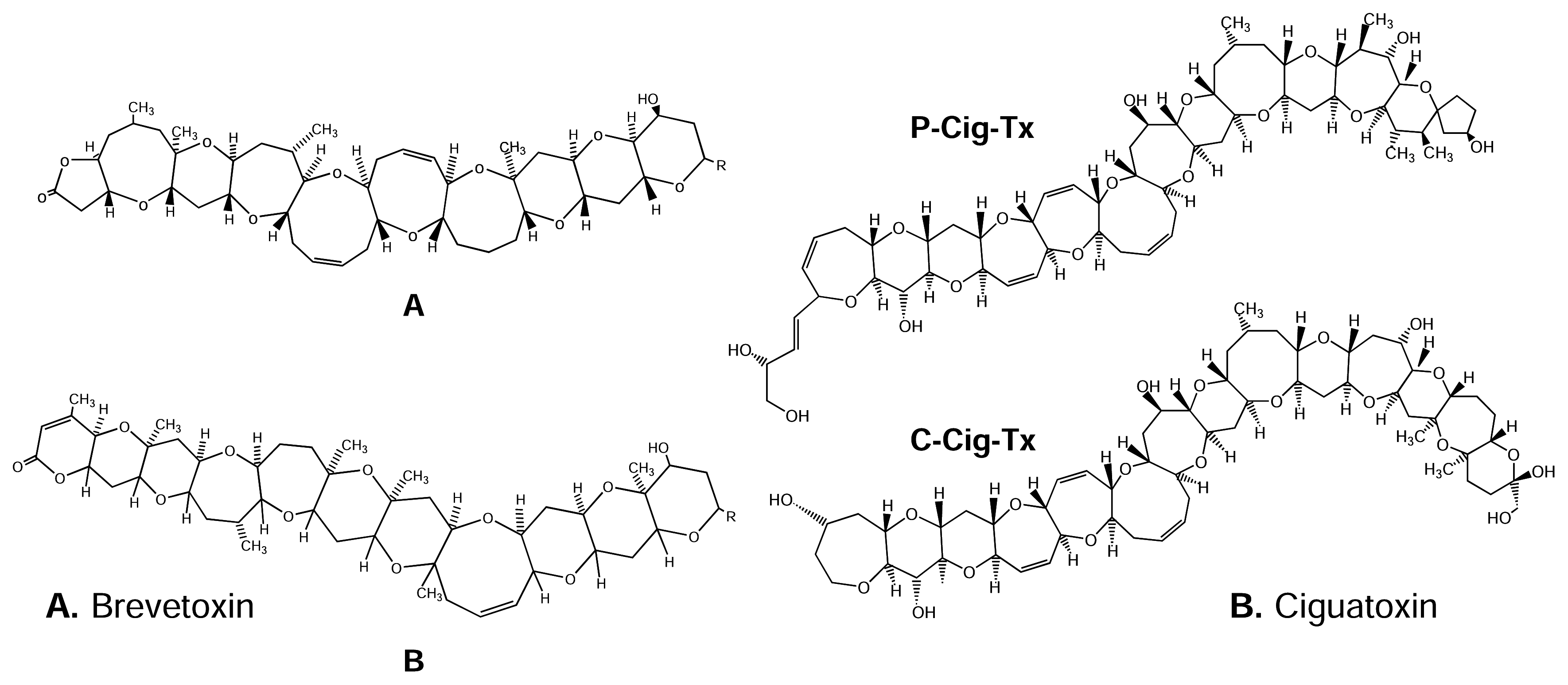
5.1.2. Brevetoxins
5.1.3. Ciguatoxins
5.1.4. Ostreotoxin-3
5.2. Peptide NaV channel gating modifiers
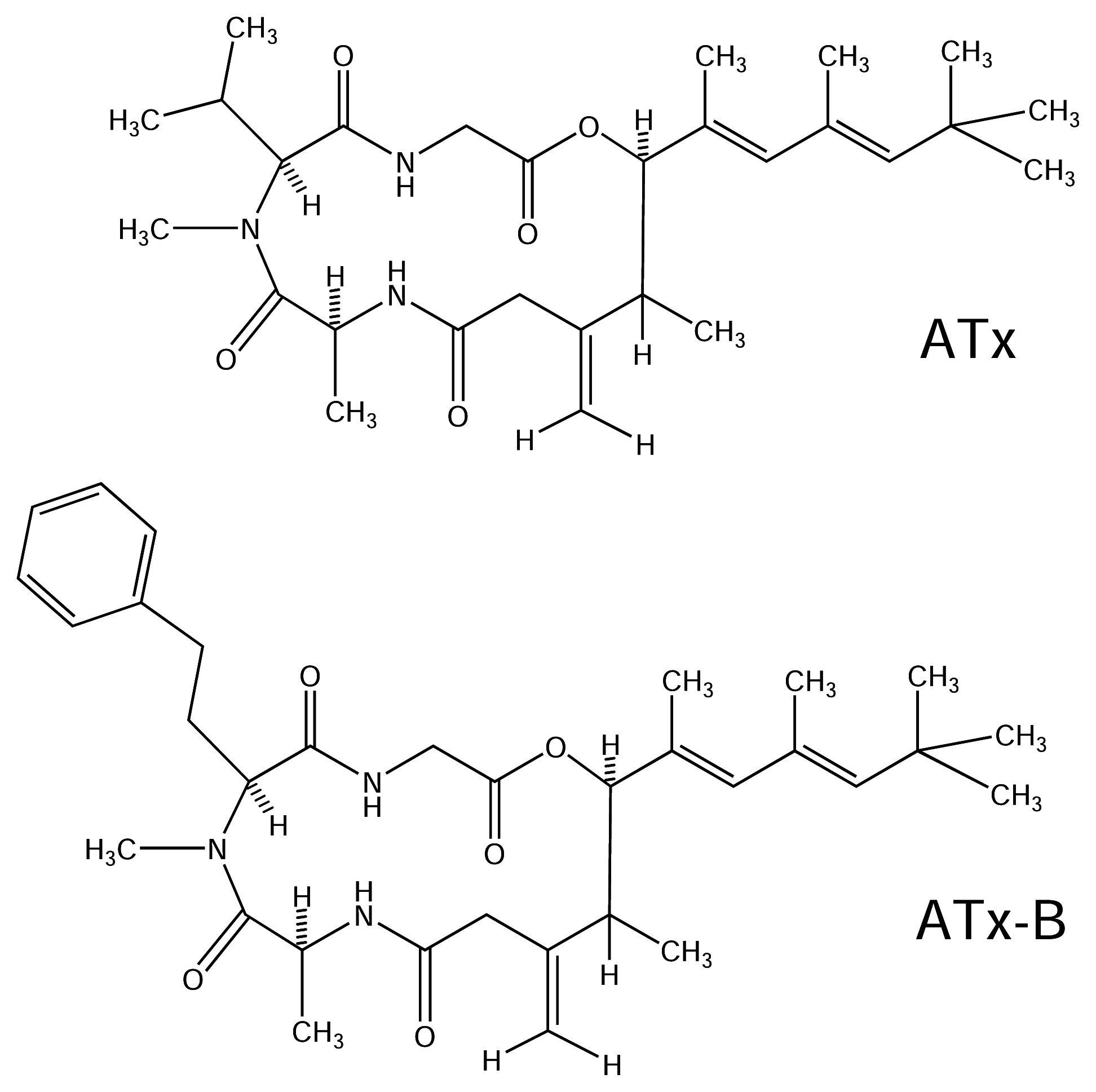
5.2.1. Antillatoxin
5.3. Peptide gating modifiers
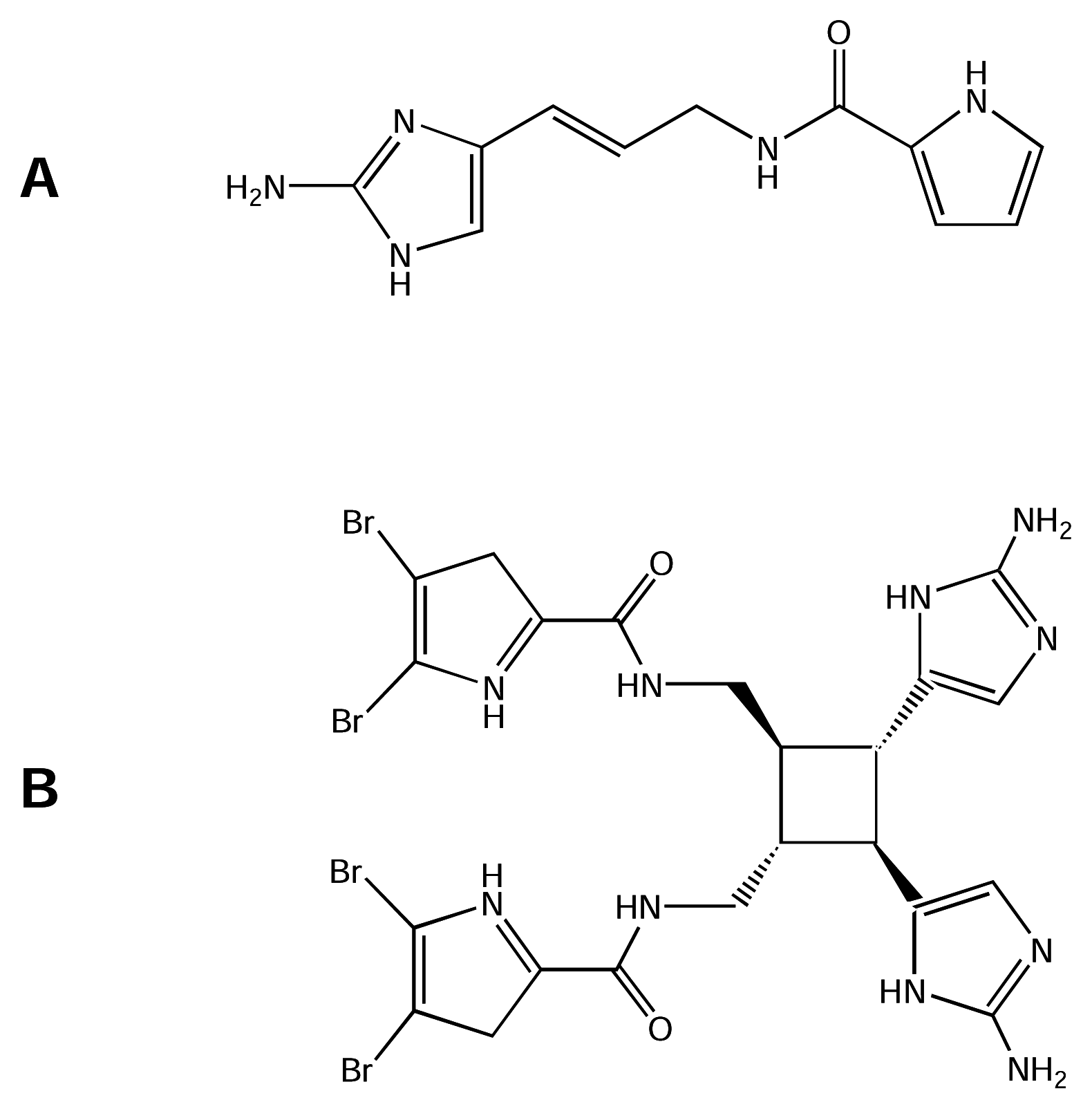
5.3.1. Sea anemone toxins
5.3.2. Goniopora Toxin
5.3.3. B-toxins
5.3.4. δ-Conotoxins
5.3.5. Conus striatus toxin
6. Concluding remarks
Acknowledgments
Abbreviations
| AneTx | anemone toxin (often abbreviated ATX in other papers) |
| ATx | antillatoxin |
| B toxins | ribbon worm toxins |
| CaV, NaV, and KV | voltage-gated ion channels selective for calcium, sodium or potassium, respectively |
| NMDA | N-methyl-D-aspartate |
| CigTx | ciguatoxin |
| CLA | clathrodin |
| CNS | central nervous system |
| PNS | peripheral nervous system |
| DBS | dibromosceptin |
| EC50 | concentration producing 50% of the maximal effect |
| GPT | Goniopora (coral) toxin |
| KTx | kalkitoxin |
| IC50 | concentration producing 50% inhibition |
| NSP | neurotoxic shellfish poisoning |
| LD50 | lethal dose/concentration for 50% of population |
| Kd | dissociation contant |
| OTx | ostreotoxin |
| PbTx-n | brevetoxins where n = 1–10 |
| PSP/PST | paralytic shellfish poisoning/toxins |
| STx | saxitoxin |
| TTx | tetrodotoxin |
| TTx-s | TTx sensitive |
| TTx-r | TTx resistant |
- Samples Availability: Available from the authors.
References
- Alonso, D.; Khalil, Z.; Satkunanthan, N.; Livett, B.G. Drugs from the sea: conotoxins as drug leads for neuropathic pain and other neurological conditions. [Review] [37 refs]. Mini-Rev. in Med. Chem 2003, 3, 785–787. [Google Scholar]
- Baden, D.G. Brevetoxins: unique polyether dinoflagellate toxins. [Review] [56 refs]. FASEB J 1989, 3, 1807–1817. [Google Scholar]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and derivative brevetoxins: historical background, multiplicity, and effects. [Review] [42 refs]. Environ. Health Persp 2005, 113, 621–625. [Google Scholar]
- Baden, D.G.; Mende, T.J. Toxicity of two toxins from the Florida red tide marine dinoflagellate, Ptychodiscus brevis. Toxicon 1982, 20, 457–461. [Google Scholar]
- Barbier, J.; Lamthanh, H.; Le Gall, F.; Favreau, P.; Benoit, E.; Chen, H.; Gilles, N.; Ilan, N.; Heinemann, S.H.; Gordon, D.; Menez, A.; Molgo, J. A delta-conotoxin from Conus ermineus venom inhibits inactivation in vertebrate neuronal Na+ channels but not in skeletal and cardiac muscles. J. Biol. Chem 2004, 279, 4680–4685. [Google Scholar]
- Barnham, K.J.; Dyke, T.R.; Kem, W.R.; Norton, R.S. Structure of neurotoxin B-IV from the marine worm Cerebratulus lacteus: a helical hairpin cross-linked by disulphide bonding. J. Mol. Biol 1997, 268, 886–902. [Google Scholar]
- Benoit, E.; Legrand, A.M.; Dubois, J.M. Effects of ciguatoxin on current and voltage clamped frog myelinated nerve fibre. Toxicon 1986, 24, 357–364. [Google Scholar]
- Beress, L.; Wunderer, G.; Wachter, E. Amino acid sequence of toxin III from Anemonia sulcata. Hoppe Seylers Z. Physiol. Chem 1977, 358, 985–988. [Google Scholar]
- Berman, F.W.; Gerwick, W.H.; Murray, T.F. Antillatoxin and kalkitoxin, ichthyotoxins from the tropical cyanobacterium Lyngbya majuscula, induce distinct temporal patterns of NMDA receptor-mediated neurotoxicity. Toxicon 1999, 37, 1645–1648. [Google Scholar]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res 2000, 28, 235–242. [Google Scholar]
- Bickmeyer, U.; Drechsler, C.; Kock, M.; Assmann, M. Brominated pyrrole alkaloids from marine Agelas sponges reduce depolarization-induced cellular calcium elevation. Toxicon 2004, 44, 45–51. [Google Scholar]
- Bidard, J.N.; Vijverberg, H.P.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem 1984, 259, 8353–8357. [Google Scholar]
- Blumenthal, K.M.; Keim, P.S.; Heinrikson, R.L.; Kem, W.R. Structure and action of heteronemertine polypeptide toxins. Amino acid sequence of Cerebratulus lacteus toxin B-II and revised structure of toxin B-IV. J. Biol. Chem 1981, 256, 9063–9067. [Google Scholar]
- Blumenthal, K.M.; Kem, W.R. Structure and action of heteronemertine polypeptide toxins. Primary structure of Cerebratulus lacteus toxin B-IV. J. Biol. Chem 1976, 251, 6025–6029. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep 2004, 21, 1–49. [Google Scholar]
- Bottein Dechraoui, M.Y.; Ramsdell, J.S. Type B brevetoxins show tissue selectivity for voltage-gated sodium channels: comparison of brain, skeletal muscle and cardiac sodium channels. Toxicon 2003, 41, 919–927. [Google Scholar]
- Bruhn, T.; Schaller, C.; Schulze, C.; Sanchez-Rodriguez, J.; Dannmeier, C.; Ravens, U.; Heubach, J.F.; Eckhardt, K.; Schmidtmayer, J.; Schmidt, H.; Aneiros, A.; Wachter, E.; Beress, L. Isolation and characterisation of five neurotoxic and cardiotoxic polypeptides from the sea anemone Anthopleura elegantissima. Toxicon 2001, 39, 693–702. [Google Scholar]
- Bulaj, G.; DeLaCruz, R.; zimi-Zonooz, A.; West, P.; Watkins, M.; Yoshikami, D.; Olivera, B.M. Delta-conotoxin structure/function through a cladistic analysis. Biochemistry 2001, 40, 13201–13208. [Google Scholar]
- Bulaj, G.; West, P.J.; Garrett, J.E.; Marsh, M.; Zhang, M.M.; Norton, R.S.; Smith, B.J.; Yoshikami, D.; Olivera, B.M. Novel conotoxins from Conus striatus and Conus kinoshitai selectively block TTX-resistant sodium channels. Biochemistry 2005, 44, 7259–7265. [Google Scholar]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria - a profile source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Seaweed dermatitis: structure of lyngbyatoxin A. Science 1979, 204, 193–195. [Google Scholar]
- Catterall, W.A. Structure and function of voltage-sensitive ion channels. Science 1988, 242, 50–61. [Google Scholar]
- Catterall, W.A. Molecular properties of a superfamily of plasma-membrane cation channels. Curr. Opin. Cell Biol 1994, 6, 607–615. [Google Scholar]
- Catterall, W.A. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron 2000, 26, 13–25. [Google Scholar]
- Catterall, W.A. Molecular mechanisms of gating and drug block of sodium channels. Novartis Found. Symp 2002, 241, 206–218. [Google Scholar]
- Catterall, W.A.; Gainer, M. Interaction of brevetoxin A with a new receptor site on the sodium channel. Toxicon 1985, 23, 497–504. [Google Scholar]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. [Review] [111 refs]. Biochimie 2000, 82, 883–892. [Google Scholar]
- Chahine, M.; Sirois, J.; Marcotte, P.; Chen, L.-Q.; Kallen, R.G. Extrapore residues of the S5–S6 loop of domain 2 of the voltage-gated skeletal muscle sodium channel (rSkM1) contribute to the μ-conotoxin GIIIA binding site. Biophys. J 1998, 75, 236–246. [Google Scholar]
- Choudhary, G.; Shang, L.; Li, X.; Dudley, S.C., Jr. Energetic localization of saxitoxin in its channel binding site. Biophys. J. 2002, 83, 912–919. [Google Scholar]
- Choudhary, G.; Yotsu-Yamashita, M.; Shang, L.; Yasumoto, T.; Dudley, S.C., Jr. Interactions of the C-11 hydroxyl of tetrodotoxin with the sodium channel outer vestibule. Biophys. J. 2003, 84, 287–294. [Google Scholar]
- Córdova, J.L.; Escudero, C.; Bustamante, J. Bloom inside the bloom: intracellular bacteria multiplication within toxic dinoflagellates. Revista de Biologia Marina y Oceanografia 2003, 38, 57–67. [Google Scholar]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; Hillyard, D.R.; Rivier, J.; McIntosh, J.M.; Cruz, L.J.; Olivera, B.M. Definition of the M-conotoxin superfamily: characterization of novel peptides from molluscivorous Conus venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar]
- Cruz, L.J.; Gray, W.R.; Olivera, B.M.; Zeikus, R.D.; Kerr, L.; Yoshikami, D.; Moczydlowski, E. Conus geographus toxins that discriminate between neuronal and muscle sodium channels. J. Biol. Chem 1985, 260, 9280–9288. [Google Scholar]
- Daly, N.L.; Ekberg, J.A.; Thomas, L.; Adams, D.J.; Lewis, R.J.; Craik, D.J. Structures of muO-conotoxins from Conus marmoreus. Inhibitors of tetrodotoxin (TTX)-sensitive and TTXresistant sodium channels in mammalian sensory neurons. J. Biol. Chem 2004, 279, 25774–25782. [Google Scholar]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, function and pharmacology of voltagegated sodium channels. [Review] [288 refs]. Naunyn-Schmiedebergs Arch. Pharmacol 2000, 362, 453–479. [Google Scholar]
- Dravid, S.M.; Baden, D.G.; Murray, T.F. Brevetoxin augments NMDA receptor signaling in murine neocortical neurons. Brain Res 2005, 1031, 30–38. [Google Scholar]
- Dudley, S.C., Jr; Chang, N.; Hall, J.; Lipkind, G.; Fozzard, H.A.; French, R.J. μ-Conotoxin interactions with the voltage-gated Na+ channel predict a clockwise arrangement of domains. J. Gen. Physiol. 2000, 116, 679–689. [Google Scholar]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and biosynthesis of the jamaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscula.[see comment]. Chem. & Biol 2004, 11, 817–833. [Google Scholar]
- Ellis, S. Introduction to symposium - brevetoxins: chemistry and pharmacology of ‘red tide’ toxins from Ptychodiscus brevis (formerly Gymnodinium breve). Toxicon 1985, 23, 469–472. [Google Scholar]
- Endean, R.; Gyr, P.; Surridge, J. The pharmacological actions on guinea-pig ileum of crude venoms from the marine gastropods Conus striatus and Conus magus. Toxicon 1977, 15, 327–337. [Google Scholar]
- Endean, R.; Gyr, P.; Surridge, J. The effects of crude venoms of Conus magus and Conus striatus on the contractile response and electrical activity of guinea-pig cardiac musculature. Toxicon 1979, 17, 381–395. [Google Scholar]
- Endean, R.; Surridge, J.; Gyr, P. Some effects of crude venom from the cones Conus striatus and Conus magus on isolated guinea-pig atria. Toxicon 1977, 15, 369–374. [Google Scholar]
- Endean, R.; Williams, H.; Gyr, P.; Surridge, J. Some effects on muscle and nerve of crude venom from the gastropod Conus striatus. Toxicon 1976, 14, 267–274. [Google Scholar]
- Escalona de Motta, G.; Rentas, A.L.; Mercado, J.A.; Tosteson, T.R.; Gonzalez, I. Identification of a sodium channel toxin from a Caribbean benthic dinoflagellate. Toxicon 1996, 34, 308. [Google Scholar]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-specific toxins from the venom of Conus textile neovicarius. Eur. J. Biochem 1991, 202, 589–595. [Google Scholar]
- Fainzilber, M.; Kofman, O.; Zlotkin, E.; Gordon, D. A new neurotoxin receptor site on sodium channels is identified by a conotoxin that affects sodium channel inactivation in molluscs and acts as an antagonist in rat brain. J. Biol. Chem 1994, 269, 2574–2580. [Google Scholar]
- Fainzilber, M.; Lodder, J.C.; Kits, K.S.; Kofman, O.; Vinnitsky, I.; Van Rietschoten, J.; Zlotkin, E.; Gordon, D. A new conotoxin affecting sodium current inactivation interacts with the delta-conotoxin receptor site. J. Biol. Chem 1995, 270, 1123–1129. [Google Scholar]
- Fainzilber, M.; van der, S.R.; Lodder, J.C.; Li, K.W.; Geraerts, W.P.; Kits, K.S. New sodium channel-blocking conotoxins also affect calcium currents in Lymnaea neurons. Biochemistry 1995, 34, 5364–5371. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep 1999, 16, 155–198. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep 2002, 19, 1–48. [Google Scholar]
- Freitas, J.C.; Rangel, M.; Oliveira, J.S.; Zaharenko, A.J.; Rozas, E. An online of marine toxicology studies in the Brazilian coast. Comm. Toxicol 2003, 9, 137–159. [Google Scholar]
- French, R.J.; Terlau, H. Sodium channel toxins--receptor targeting and therapeutic potential. [Review] [85 refs]. Curr. Med. Chem 2004, 11, 3053–3064. [Google Scholar]
- Fu, L.Y.; Li, Y.; Cheng, L.; Zhou, H.Y.; Yao, W.X.; Xia, G.J.; Jiang, M.X. Effect of sea anemone toxin anthopleurin-Q on sodium current in guinea pig ventricular myocytes. Acta Pharmacol. Sinica 2001, 22, 1107–1112. [Google Scholar]
- Fujiwara, M.; Muramatsu, I.; Hidaka, H.; Ikushima, S.; Ashida, K. Effects of Goniopora toxin, a polypeptide isolated from coral, on electromechanical properties of rabbit myocardium. J. Pharmacol. Exp. Ther 1979, 210, 153–157. [Google Scholar]
- Gallacher, S.; Flynn, K.J.; Franco, J.M.; Brueggemann, E.E.; Hines, H.B. Evidence for production of paralytic shellfish toxins by bacteria associated with Alexandrium spp. (Dinophyta) in culture. Appl. Env. Microbiol 1997, 63, 239–245. [Google Scholar]
- Gerwick, W.H.; Fenical, W.; Norris, J.N. Chemical variation in the tropical seaweed Stypopodium zonale (dictyotaceae). Phytochemistry 1985, 24, 1279–1283. [Google Scholar]
- Gonoi, T.; Ashida, K.; Feller, D.; Schmidt, J.; Fujiwara, M.; Catterall, W.A. Mechanism of action of a polypeptide neurotoxin from the coral Goniopora on sodium channels in mouse neuroblastoma cells. Mol. Pharmacol 1986, 29, 347–354. [Google Scholar]
- Gonoi, T.; Hille, B. Gating of Na channels. Inactivation modifiers discriminate among models. J. Gen. Physiol 1987, 89, 253–274. [Google Scholar]
- Gonoi, T.; Ohizumi, Y.; Kobayashi, J.; Nakamura, H.; Catterall, W.A. Actions of a polypeptide toxin from the marine snail Conus striatus on voltage-sensitive sodium channels. Molecular Pharmacology 1987, 32, 691–698. [Google Scholar]
- Hashimoto, Y.; Ashida, K. Screening of toxic corals and isolation of a toxic polypeptide from Goniopora spp. Pub. the Seto Marine Biol. Lab 1973, 20, 703–711. [Google Scholar]
- Hasson, A.; Shon, K.J.; Olivera, B.M.; Spira, M.E. Alterations of voltage-activated sodium current by a novel conotoxin from the venom of Conus gloriamaris. J. Neurophysiol 1995, 73, 1295–1301. [Google Scholar]
- Hill, J.M.; Alewood, P.F.; Craik, D.J. Conotoxin TVIIA, a novel peptide from the venom of Conus tulipa 2. Three-dimensional solution structure. Eur. J. Biochem 2000, 267, 4649–4657. [Google Scholar]
- Hille, B. The permeability of the sodium channel to metal cations in myelinated nerve. J. Gen. Physiol 1972, 59, 637–658. [Google Scholar]
- Hille, B. Ion channels of excitable membranes; Sunderland; Sinauer Associates, Inc, 2001. [Google Scholar]
- Hillyard, D.R.; Olivera, B.M.; Woodward, S.; Corpuz, G.P.; Gray, W.R.; Ramilo, C.A.; Cruz, L.J. A molluscivorous Conus toxin: conserved frameworks in conotoxins. Biochemistry 1989, 28, 358–361. [Google Scholar]
- Honma, T.; Iso, T.; Ishida, M.; Nagashima, Y.; Shiomi, K. Occurrence of type 3 sodium channel peptide toxins in two species of sea anemones (Dofleinia armata and Entacmaea ramsayi). Toxicon 2003, 41, 637–639. [Google Scholar]
- Howell, M.L.; Blumenthal, K.M. Mutagenesis of Cerebratulus lacteus neurotoxin B-IV identifies NH2-terminal sequences important for biological activity. J. Biol. Chem 1991, 266, 12884–12888. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: visual molecular dynamics. J. Mol. Graphics 1996, 14, 33–38. [Google Scholar]
- Ishida, H.; Nozawa, A.; Totoribe, K.; Muramatsu, N.; Nukaya, H.; Tsuji, K.; Yamaguchi, K.; Yasumoto, T.; Kaspar, H.; Berkett, N.; Kosuge, T. Brevetoxin B1, a new polyether marine toxin from the New Zealand shellfish, Austrovenus stutchburyi. Tetrahedron Letters 1995, 36, 725–728. [Google Scholar]
- Isom, L.L. Sodium channel β subunits: anything but auxiliary. Neuroscientist 2001, 7(1), 42–54. [Google Scholar]
- Isom, L.L.; De Jongh, K.S.; Catterall, W.A. Auxiliary subunits of voltage-gated ion channels. Neuron 1994, 12, 1183–1194. [Google Scholar]
- Jeglitsch, G.; Rein, K.; Baden, D.G.; Adams, D.J. Brevetoxin-3 (PbTx-3) and its derivatives modulate single tetrodotoxin-sensitive sodium channels in rat sensory neurons. J. Pharmacol. Exp. Ther 1998, 284, 516–525. [Google Scholar]
- Keizer, D.W.; West, P.J.; Lee, E.F.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structural basis for tetrodotoxin-resistant sodium channel binding by mu-conotoxin SmIIIA. J. Biol. Chem 2003, 278, 46805–46813. [Google Scholar]
- Kem, W.R. Purification and characterization of a new family of polypeptide neurotoxins from the heteronemertine Cerebratulus lacteus (Leidy). J. Biol. Chem 1976, 251, 4184–4192. [Google Scholar]
- Kem, W.R.; Parten, B.; Pennington, M.W.; Price, D.A.; Dunn, B.M. Isolation, characterization, and amino acid sequence of a polypeptide neurotoxin occurring in the sea anemone Stichodactyla helianthus. Biochemistry 1989, 28, 3483–3489. [Google Scholar]
- Kobayashi, J.; Nakamura, H.; Hirata, Y.; Ohizumi, Y. Isolation of a cardiotonic glycoprotein, striatoxin, from the venom of the marine snail Conus Striatus. Biochem. Biophys. Res. Commun 1982, 105, 1389–1395. [Google Scholar]
- Kohno, T.; Sasaki, T.; Kobayashi, K.; Fainzilber, M.; Sato, K. Three-dimensional solution structure of the sodium channel agonist/antagonist delta-conotoxin TxVIA. J. Biol. Chem 2002, 277, 36387–36391. [Google Scholar]
- Kulagina, N.V.; O’shaughnessy, T.J.; Ma, W.; Ramsdell, J.S.; Pancrazio, J.J. Pharmacological effects of the marine toxins, brevetoxin and saxitoxin, on murine frontal cortex neuronal networks. Toxicon 2004, 44, 669–676. [Google Scholar]
- Lehane, L.; Lewis, R.J. Ciguatera: recent advances but the risk remains. Int. J. Food Microbiol 2000, 61, 91–125. [Google Scholar]
- Leipold, E.; Hansel, A.; Olivera, B.M.; Terlau, H.; Heinemann, S.H. Molecular interaction of delta-conotoxins with voltage-gated sodium channels. FEBS Lett 2005, 579, 3881–3884. [Google Scholar]
- LePage, K.T.; Goeger, D.; Yokokawa, F.; Asano, T.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. The neurotoxic lipopeptide kalkitoxin interacts with voltage-sensitive sodium channels in cerebellar granule neurons. Toxicol. Lett 2005, 158, 133–139. [Google Scholar]
- Lewis, R.J. Ciguatoxins are potent ichthyotoxins. Toxicon 1992, 30, 207–211. [Google Scholar]
- Lewis, R.J. The changing face of ciguatera. [Review] [64 refs]. Toxicon 2001, 39, 97–106. [Google Scholar]
- Li, R.A.; Ennis, I.L.; French, R.J.; Dudley, S.C., Jr.; Tomaselli, G.F.; Marbán, E. Clockwise domain arrangement of the sodium channel revealed by μ−conotoxin (GIIIA) docking orientation. J. Biol. Chem. 2001, 276(14), 11072–11077. [Google Scholar]
- Li, R.A.; Ennis, I.L.; Xue, T.; Nguyen, H.M.; Tomaselli, G.F.; Goldin, A.L.; Marban, E. Molecular basis of isoform-specific μ−conotoxin block of cardiac, skeletal muscle, and brain Na+ channels. J. Biol. Chem 2003, 278, 8717–8724. [Google Scholar]
- Li, W.I.; Berman, F.W.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. Antillatoxin is a marine cyanobacterial toxin that potently activates voltage-gated sodium channels. Proc. Natl. Acad. Sci. U. S. A 2001, 98, 7599–7604. [Google Scholar]
- Lindel, T.; Hoffman, H.; Hochgurtel, M.; Pawlik, J.R. Structure-activity relationship of inhibition of fish feeding by sponge-derived and synthetic pyrrole-imidazole alkaloids. J. Chem. Ecol. 2000, 26. [Google Scholar]
- Llewellyn, L.; Negri, A.; Quilliam, M. High affinity for the rat brain sodium channel of newly discovered hydroxybenzoate saxitoxin analogues from the dinoflagellate Gymnodinium catenatum. Toxicon 2004, 43, 101–104. [Google Scholar]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett 1987, 219, 355–359. [Google Scholar]
- Long, S.B.; Campbell, E.B.; MacKinnon, R. Crystal structure of a mammalian voltagedependent Shaker family K+ channel. Science 2005, 309, 897–903. [Google Scholar]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int 1995, 78, 521–527. [Google Scholar]
- Martins, C.A.; Alvito, P.; Tavares, M.J.; Pereira, P.; Doucette, G.; Franca, S. Reevaluation of production of paralytic shellfish toxin by bacteria associated with dinoflagellates of the Portuguese coast. Appl. Environ. Microbiol 2003, 69, 5693–5698. [Google Scholar]
- Mattei, C.; Dechraoui, M.Y.; Molgo, J.; Meunier, F.A.; Legrand, A.M.; Benoit, E. Neurotoxins targetting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J. Neurosci. Res 1999, 55, 666–673. [Google Scholar]
- McFarren, E.F.; Silva, F.J.; Tanabe, H.; Wilson, W.B.; Campbell, J.E.; Lewis, K.H. The occurrence of a ciguatera-like poison in oysters, clams, and Gymnodinium breve cultures. Toxicon 1965, 3, 111–123. [Google Scholar]
- McIntosh, J.M.; Hasson, A.; Spira, M.E.; Gray, W.R.; Li, W.; Marsh, M.; Hillyard, D.R.; Olivera, B.M. A new family of conotoxins that blocks voltage-gated sodium channels. J. Biol. Chem 1995, 270, 16796–16802. [Google Scholar]
- Morales, J.J.; Rodriguez, A.D. The structure of clathrodin, a novel alkaloid isolated from the Caribbean Sea sponge Agelas clathrodes. J. Nat. Prod 1991, 54, 629–631. [Google Scholar]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Osillatoriaceae: isolation of Debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar]
- Nielsen, K.J.; Watson, M.; Adams, D.J.; Hammarstrom, A.K.; Gage, P.W.; Hill, J.M.; Craik, D.J.; Thomas, L.; Adams, D.; Alewood, P.F.; Lewis, R.J. Solution structure of mu-conotoxin PIIIA, a preferential inhibitor of persistent tetrodotoxin-sensitive sodium channels. J. Biol. Chem 2002, 277, 27247–27255. [Google Scholar]
- Nishida, S.; Fujita, S.; Warashina, A.; Satake, M.; Tamiya, N. Amino acid sequence of a sea anemone toxin from Parasicyonis actinostoloides. Eur. J. Biochem 1985, 150, 171–173. [Google Scholar]
- Noda, M.; Suzuki, H.; Numa, S.; Stühmer, W. A single point mutation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II. FEBS Lett 1989, 259, 213–216. [Google Scholar]
- Nogle, L.M.; Okino, T.; Gerwick, W.H. Antillatoxin B, a neurotoxic lipopeptide from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod 2001, 64, 983–985. [Google Scholar]
- Norton, R.S. Structure and structure-function relationships of sea anemone proteins that interact with the sodium channel. [Review] [127 refs]. Toxicon 1991, 29, 1051–1084. [Google Scholar]
- Norton, R.S.; Pallaghy, P.K. The cystine knot structure of ion channel toxins and related polypeptides. Toxicon 1998, 36, 1573–1583. [Google Scholar]
- Ogata, N.; Ohishi, Y. Molecular diversity of structure and function of the voltage-gated Na+ channels. [Review] [82 refs]. Jap. J. Pharmacol 2002, 88, 365–377. [Google Scholar]
- Olivera, B.M. E.E. Just Lecture, 1996. Conus venom peptides, receptor and ion channel targets, and drug design: 50 million years of neuropharmacology. Mol. Biol. Cell 1997, 8, 2101–2109. [Google Scholar]
- Olivera, B.M. Conus venom peptides: reflections from the biology of clades and species. Annu. Rev. Ecol. Syst 2002, 33, 25–47. [Google Scholar]
- Orjala, J.; Nagle, D.G.; Hsu, V.L.; Gerwick, W.H. Antillatoxin: an exceptionally ichthyotoxic cyclic lipopeptide from the tropical cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc 1995, 117, 8281–8282. [Google Scholar]
- Pavlov, E; Bladen, C; Diao, C; French, RJ. Bacterial Na channels: progenitors, progeny, or parallel evolution? In Bacterial ion channels and eukaryotic homologues; Kubalski, A., Martinac, B., Eds.; American Society for Microbiology, 2005; pp. 191–207. [Google Scholar]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psych 2001, 70, 4–8. [Google Scholar]
- Penzotti, J.L.; Fozzard, H.A.; Lipkind, G.M.; Dudley, S.C., Jr. Differences in saxitoxin and tetrodotoxin binding revealed by mutagenesis of the Na+ channel outer vestibule. Biophys. J. 1998, 75, 2647–2657. [Google Scholar]
- Penzotti, J.L.; Lipkind, G.; Fozzard, H.A.; Dudley, S.C., Jr. Specific neosaxitoxin interactions with the Na+ channel outer vestibule determined by mutant cycle analysis. Biophys. J. 2001, 80, 698–706. [Google Scholar]
- Pierce, R.H. Red tide (Ptychodiscus brevis) toxin aerosols: a review. Toxicon 1986, 24, 955–965. [Google Scholar]
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes. Mol. Pharmacol 1986, 30, 129–135. [Google Scholar]
- Pomati, F.; Rossetti, C.; Calamari, D.; Neilan, B.A. Effects of saxitoxin (STx) and Veratridine on Bacterial Na+-K+ fluxes: a Prokaryote-based STx bioassay. Appl. Environ. Microbiol 2003, 69, 7371–7376. [Google Scholar]
- Reimer, N.S.; Yasunobu, C.L.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin-B. J. Biol. Chem 1985, 260, 8690–8693. [Google Scholar]
- Rein, K.S.; Baden, D.G.; Gawley, R.E. Conformational analysis of the sodium channel modulator, brevetoxin A, comparison with brevetoxin B conformations, and a hypothesis about the common pharmacophore of the “site 5” toxins. J. Org. Chem 1994, 59, 2101–2106. [Google Scholar]
- Rein, K.S.; Lynn, B.; Gawley, R.E.; Baden, D.G. Brevetoxin B: chemical modifications, synaptosome binding, toxicity, and an unexpected conformational effect. J. Org. Chem 1994, 59, 2107–2113. [Google Scholar]
- Ren, D.; Navarro, B.; Xu, H.; Yue, L.; Shi, Q.; Clapham, D.E. A prokaryotic voltage-gated sodium channel. Science 2001, 294, 2372–2375. [Google Scholar]
- Rentas, A.L.; Rosa, R.; Rodriguez, A.D.; De Motta, G.E. Effect of alkaloid toxins from tropical marine sponges on membrane sodium currents. Toxicon 1995, 33, 491–497. [Google Scholar]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. U. S. A 2006, 103, 3176–3179. [Google Scholar]
- Rogers, J.C.; Qu, Y.; Tanada, T.N.; Scheuer, T.; Catterall, W.A. Molecular determinants of high affinity binding of alpha-scorpion toxin and sea anemone toxin in the S3–S4 extracellular loop in domain IV of the Na+ channel alpha subunit. J. Biol. Chem 1996, 271, 15950–15962. [Google Scholar]
- Rohl, C.A.; Boeckman, F.A.; Baker, C.; Scheuer, T.; Catterall, W.A.; Klevit, R.E. Solution structure of the sodium channel inactivation gate. Biochemistry 1999, 38, 855–861. [Google Scholar]
- Sabry, O.M.; Andrews, S.; McPhail, K.L.; Goeger, D.E.; Yokochi, A.; LePage, K.T.; Murray, T.F.; Gerwick, W.H. Neurotoxic meroditerpenoids from the tropical marine brown alga Stypopodium flabelliforme. J. Nat. Prod 2005, 68, 1022–1030. [Google Scholar]
- Safo, P.; Rosenbaum, T.; Shcherbatko, A.; Choi, D.Y.; Han, E.; Toledo-Aral, J.J.; Olivera, B.M.; Brehm, P.; Mandel, G. Distinction among neuronal subtypes of voltage-activated sodium channels by mu-conotoxin PIIIA. J. Neurosci 2000, 20, 76–80. [Google Scholar]
- Scheffler, J.J.; Tsugita, A.; Linden, G.; Schweitz, H.; Lazdunski, M. The amino acid sequence of toxin V from Anemonia sulcata. Biochem. Biophys. Res. Commun 1982, 107, 272–278. [Google Scholar]
- Schlief, T.; Schonherr, R.; Imoto, K.; Heinemann, S.H. Pore properties of rat brain II sodium channels mutated in the selectivity filter domain. Eur. Biophys. J 1996, 25, 75–91. [Google Scholar]
- Seibert, A.L.; Liu, J.; Hanck, D.A.; Blumenthal, K.M. Arg-14 loop of site 3 anemone toxins: effects of glycine replacement on toxin affinity. Biochemistry 2003, 42, 14515–14521. [Google Scholar]
- Seoh, S.A.; Sigg, D.; Papazian, D.M.; Bezanilla, F. Voltage-sensing residues in the S2 and S4 segments of the Shaker K+ channel. Neuron 1996, 16, 1159–1167. [Google Scholar]
- Sharkey, R.G.; Jover, E.; Couraud, F.; Baden, D.G.; Catterall, W.A. Allosteric modulation of neurotoxin binding to voltage-sensitive sodium channels by Ptychodiscus brevis toxin 2. Mol. Pharmacol 1987, 31, 273–278. [Google Scholar]
- Shichor, I.; Fainzilber, M.; Pelhate, M.; Malecot, C.O.; Zlotkin, E.; Gordon, D. Interactions of delta-conotoxins with alkaloid neurotoxins reveal differences between the silent and effective binding sites on voltage-sensitive sodium channels. J. Neurochem 1996, 67, 2451–2460. [Google Scholar]
- Shimizu, Y. Microalgal metabolites. Chem. Rev 1993, 93, 1685–1698. [Google Scholar]
- Shon, K.J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, characterization, synthesis, and cloning of the lockjaw peptide from Conus purpurascens venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar]
- Shon, K.J.; Hasson, A.; Spira, M.E.; Cruz, L.J.; Gray, W.R.; Olivera, B.M. Delta-conotoxin GmVIA, a novel peptide from the venom of Conus gloriamaris. Biochemistry 1994, 33, 11420–11425. [Google Scholar]
- Shon, K.-J.; Olivera, B.M.; Watkins, M.; Jacobsen, R.B.; Gray, W.R.; Floresca, C.Z.; Cruz, L.J.; Hillyard, D.R.; Brink, A.; Terlau, H.; Yoshikami, D. μ-conotoxin PIIIA, a new peptide for discriminating among tetrodotoxin-sensitive Na channel subtypes. J. Neurosci. 1998, 18(12), 4473–4481. [Google Scholar]
- Smith, E.A.; Grant, F.; Ferguson, C.M.; Gallacher, S. Biotransformations of paralytic shellfish toxins by bacteria isolated from bivalve molluscs. Appl. Env. Microbiol 2001, 67, 2345–2353. [Google Scholar]
- Snyder, R.V.; Guerrero, M.A.; Sinigalliano, C.D.; Winshell, J.; Perez, R.; Lopez, J.V.; Rein, K.S. Localization of polyketide synthase encoding genes to the toxic dinoflagellate Karenia brevis. Phytochemistry 2005, 66, 1767–1780. [Google Scholar]
- Stephan, M.M.; Potts, J.F.; Agnew, W.S. The μI skeletal muscle sodium channel: mutation E403Q eliminates sensitivity to tetrodotoxin but not to μ-conotoxins GIIIA and GIIIB. J. Membrane Biol 1994, 137, 1–8. [Google Scholar]
- Strachan, L.C.; Lewis, R.J.; Nicholson, G.M. Differential actions of pacific ciguatoxin-1 on sodium channel subtypes in mammalian sensory neurons. J Pharmacol. Exp. Ther 1999, 288, 379–388. [Google Scholar]
- Suarez-Isla, B; Guzman-Mendez, L. Mareas rojas y toxinas marinas. Floraciones de algas nocivas; Santiago de Chile; Editorial Universitaria, 1998. [Google Scholar]
- Sudarslal, S.; Majumdar, S.; Ramasamy, P.; Dhawan, R.; Pal, P.P.; Ramaswami, M.; Lala, A.K.; Sikdar, S.K.; Sarma, S.P.; Krishnan, K.S.; Balaram, P. Sodium channel modulating activity in a delta-conotoxin from an Indian marine snail. FEBS Lett 2003, 553, 209–212. [Google Scholar]
- Sunahara, S.; Muramoto, K.; Tenma, K.; Kamiya, H. Amino acid sequence of two sea anemone toxins from Anthopleura fuscoviridis. Toxicon 1987, 25, 211–219. [Google Scholar]
- Tanaka, M.; Hainu, M.; Yasunobu, K.T.; Norton, T.R. Amino acid sequence of the Anthopleura xanthogrammica heart stimulant, anthopleurin A. Biochemistry 1977, 16, 204–208. [Google Scholar]
- Terlau, H.; Heinemann, S.H.; Stühmer, W.; Pusch, M.; Conti, F.; Imoto, K.; Numa, S. Mapping the site of block by tetrodotoxin and saxitoxin of sodium channel II. FEBS Lett 1991, 293, 93–96. [Google Scholar]
- Terlau, H.; Olivera, B.M. Conus venoms: a rich source of novel ion channel-targeted peptides. [Review] [196 refs]. Physiol. Rev 2004, 84, 41–68. [Google Scholar]
- Terlau, H.; Shon, K.J.; Grilley, M.; Stocker, M.; Stuhmer, W.; Olivera, B.M. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar]
- Terlau, H.; Stocker, M.; Shon, K.-J.; McIntosh, J.M.; Olivera, B.M. μO-conotoxin MrVIA inhibits mammalian sodium channels, but not through site I. J. Neurophysiol. 1996, 76(3), 1423–1429. [Google Scholar]
- Todt, H.; Dudley, S.C., Jr; Kyle, J.W.; French, R.J.; Fozzard, H.A. Ultra-slow inactivation in mu1 Na+ channels is produced by a structural rearrangement of the outer vestibule. Biophys. J. 1999, 76, 1335–1345. [Google Scholar]
- Vasquez, M.; Gruttner, C.; Moeller, B.; Moore, E.R. Limited selection of sodium channel blocking toxin-producing bacteria from paralytic shellfish toxin-contaminated mussels (Aulacomya ater). Res. Microbiol 2002, 153, 333–338. [Google Scholar]
- Vilin, Y.Y.; Ruben, P.C. Slow inactivation in voltage-gated sodium channels: molecular substrates and contributions to channelopathies. Cell Biochem. Biophys 2001, 35, 171–190. [Google Scholar]
- Volpon, L.; Lamthanh, H.; Barbier, J.; Gilles, N.; Molgo, J.; Menez, A.; Lancelin, J.M. NMR solution structures of delta-conotoxin EVIA from Conus ermineus that selectively acts on vertebrate neuronal Na+ channels. J. Biol. Chem 2004, 279, 21356–21366. [Google Scholar]
- Wang, S.Y.; Wang, G.K. Voltage-gated sodium channels as primary targets of diverse lipid-soluble neurotoxins. [Review] [68 refs]. Cell. Signal 2003, 15, 151–159. [Google Scholar]
- Wemmer, D.E.; Kumar, N.V.; Metrione, R.M.; Lazdunski, M.; Drobny, G.; Kallenbach, N.R. NMR analysis and sequence of toxin II from the sea anemone Radianthus paumotensis. Biochemistry 1986, 25, 6842–6849. [Google Scholar]
- Wen, P.H.; Blumenthal, K.M. Role of electrostatic interactions in defining the potency of neurotoxin B-IV from Cerebratulus lacteus. J. Biol. Chem 1996, 271, 29752–29758. [Google Scholar]
- Wen, P.H.; Blumenthal, K.M. Structure and function of Cerebratulus lacteus neurotoxin B-IV: tryptophan-30 is critical for function while lysines-18, -19, -29, and -33 are not required. Biochemistry 1997, 36, 13435–13440. [Google Scholar]
- Wessels, M.; Konig, G.M.; Wright, A.D. A new tyrosine kinase inhibitor from the marine brown alga Stypopodium zonale. J. Nat. Prod 1999, 62, 927–930. [Google Scholar]
- West, P.J.; Bulaj, G.; Garrett, J.E.; Olivera, B.M.; Yoshikami, D. Mu-conotoxin SmIIIA, a potent inhibitor of tetrodotoxin-resistant sodium channels in amphibian sympathetic and sensory neurons. Biochemistry 2002, 41, 15388–15393. [Google Scholar]
- West, P.J.; Bulaj, G.; Yoshikami, D. Effects of delta-conotoxins PVIA and SVIE on sodium channels in the amphibian sympathetic nervous system. J. Neurophysiol 2005, 94, 3916–3924. [Google Scholar]
- Widmer, H.; Billeter, M.; Wuthrich, K. Three-dimensional structure of the neurotoxin ATX Ia from Anemonia sulcata in aqueous solution determined by nuclear magnetic resonance spectroscopy. Proteins 1989, 6, 357–371. [Google Scholar]
- Wolfe, G.V. The chemical defense ecology of marine unicellular plankton: constraints, mechanisms, and impacts. [Review] [270 refs]. Biol. Bull 2000, 198, 225–244. [Google Scholar]
- Wu, M.; Okino, T.; Nogle, L.M.; Marquez, B.L.; Williamson, R.T.; Sitachitta, N.; Berman, F.W.; Murray, T.F.; McGough, K.; Jacobs, R.; Colsen, K.; Asano, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H. Structure, synthesis, and biological properties of kalkitoxin, a novel neurotoxin from the marine cyanobacterium Lyngbya majuscule. J. Am. Chem. Soc 2000, 122, 1241–1242. [Google Scholar]
- Wunderer, G.; Eulitz, M. Amino-acid sequence of toxin I from Anemonia sulcata. Eur. J. Biochem 1978, 89, 11–17. [Google Scholar]
- Wunderer, G.; Fritz, H.; Wachter, E.; Machleidt, W. Amino-acid sequence of a coelenterate toxin: toxin II from Anemonia sulcata. Eur. J. Biochem 1976, 68, 193–198. [Google Scholar]
- Yanagawa, Y.; Abe, T.; Satake, M.; Odani, S.; Suzuki, J.; Ishikawa, K. A novel sodium channel inhibitor from Conus geographus: purification, structure, and pharmacological properties. Biochemistry 1988, 27, 6256–6262. [Google Scholar]
- Yasumoto, T.; murata, M. Marine toxins. Chem. Rev 1993, 93, 1897–1909. [Google Scholar]
- Yotsu-Yamashita, M.; Kim, Y.H.; Dudley, S.C., Jr; Choudhary, G.; Pfahnl, A.; Oshima, Y.; Daly, J.W. The structure of zetekitoxin AB, a saxitoxin analog from the Panamanian golden frog Atelopus zeteki: a potent sodium-channel blocker. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 4346–4351. [Google Scholar]
- Zingone, A.; Enevoldsen, H.O. The diversity of harmful algal blooms: a challenge for science and management. Ocean & Coastal Management 2000, 43, 725–748. [Google Scholar]
- Zlotkin, E. The insect voltage-gated sodium channel as target of insecticides. [Review] [73 refs]. Ann. Rev. Entomol 1999, 44, 429–455. [Google Scholar]
- Zykova, T.A.; Kozlovskaia, E.P. (Amino acid sequence of a neurotoxin from the anemone Radianthus macrodactylus). Bioorg. Khim 1989, 15, 1301–1306.
- Zykova, T.A.; Kozlovskaia, E.P. (Disulfide bonds in neurotoxin-III from the sea anenome Radianthus macrodactylus). Bioorg. Khim 1989, 15, 904–907.
- Zykova, T.A.; Kozlovskaia, E.P.; Eliakov, G.B. (Amino acid sequence of neurotoxin II from the sea anemone Radianthus macrodactylus). Bioorg. Khim 1988, 14, 878–882.
- Zykova, T.A.; Kozlovskaia, E.P.; Eliakov, G.B. (Amino acid sequence of neurotoxins IV and V from the sea anemone Radianthus macrodactylus). Bioorg. Khim. 1988, 14, 1489–1494.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor Site | Physiological Effect | Putative location | Toxin | Organism |
|---|---|---|---|---|
| 1 | Pore blocker | P loop at DI, DII, DIII, DIV | Tetrodotoxins | Puffer fish (Tetraodontiformes), starfish, molluscs, tunicates, dinoflagellates, symbiotic bacteria |
| Saxitoxins | Shellfishes, Red tide dinoflagellates and symbiotic bacteria | |||
| μ-Conotoxins | Cone snails | |||
| 2 | Persistent activation gating; | DI-S6 | Batrachotoxin | [Dendrobatid “poison dart” frogs] |
| Depolarization of resting potentials | DII:S6 | Veratridine | [In the seed of Schoenocaulon officinale and in rhizome of Veratrum album] | |
| Repetitive firings | DIII:S6 | Aconitine | [In the root of Aconitum plants] | |
| DIV:S6 | Grayanotoxin | [In plants of the family Ericaceae;] | ||
| 3 | Prolonged Na+ channel opening | DIV:S3–S4 | α-Scorpion toxins | [Scorpion spp.] |
| DI:S3–S4 Loop | Sea anemone toxins | Sea anemones spp. | ||
| Inactivation gating modifier | DIV:S5–S6 Loop | δ-Atracotoxins | [Funnel-web spider Hadronyche versuta] | |
| 4 | Agonist | ? | Antillatoxin | Dinoflagellate Lyngbya majuscule |
| Shifts in activation gating; repetitive firings | DII:S3–S4 | Loop β-Scorpion toxins | [Scorpion spp,] | |
| 5 | Shift in activation gating | DI:S6 | Brevetoxins | Red tide dinoflagellate Karenia brevis |
| DIV:S5 | Ciguatoxins | Red tide dinoflagellate Gambierdiscus toxicus and tropical coral fishes | ||
| 6 | Prolonged Na+ channel opening | DIV:S3–S4 | δ-Conotoxins | Cone snails |
| 7 | Inhibitor, not a pore blocker | ? | Kalkitoxin | Dinoflagellate Lyngbya majuscule |
| Persistent activation gating; depolarization of resting potentials; repetitive firings | DI-S6 DII-S6 DIII-S6 | DDT and analogs, Pyrethroids | [Insecticide chemicals] | |
| 8 | Inactivation gating modifier | ? | Goniopora | genus Goniopora |
| Prolonged Na+ channel opening | ? | Conus striatus toxin | Conus striatus | |
| 9 | Inhibition of Na+ permeability | DI-S6 DIII-S6 DIV-S6 | Local anesthetics, Anticonvulsants, Antidepressants | [Chemicals from various sources] |
| N.I. | Inhibitor, not a pore blocker | ? | μO-Conotoxins | Cone snails |
| N.I. | Inactivation gating modifier | ? | B-Toxins | Ribbon worm (Cerebratulus lacteus) |
| N.I. | Inhibitor, not a pore blocker | ? | Meroditerpenoids | Brown algae |
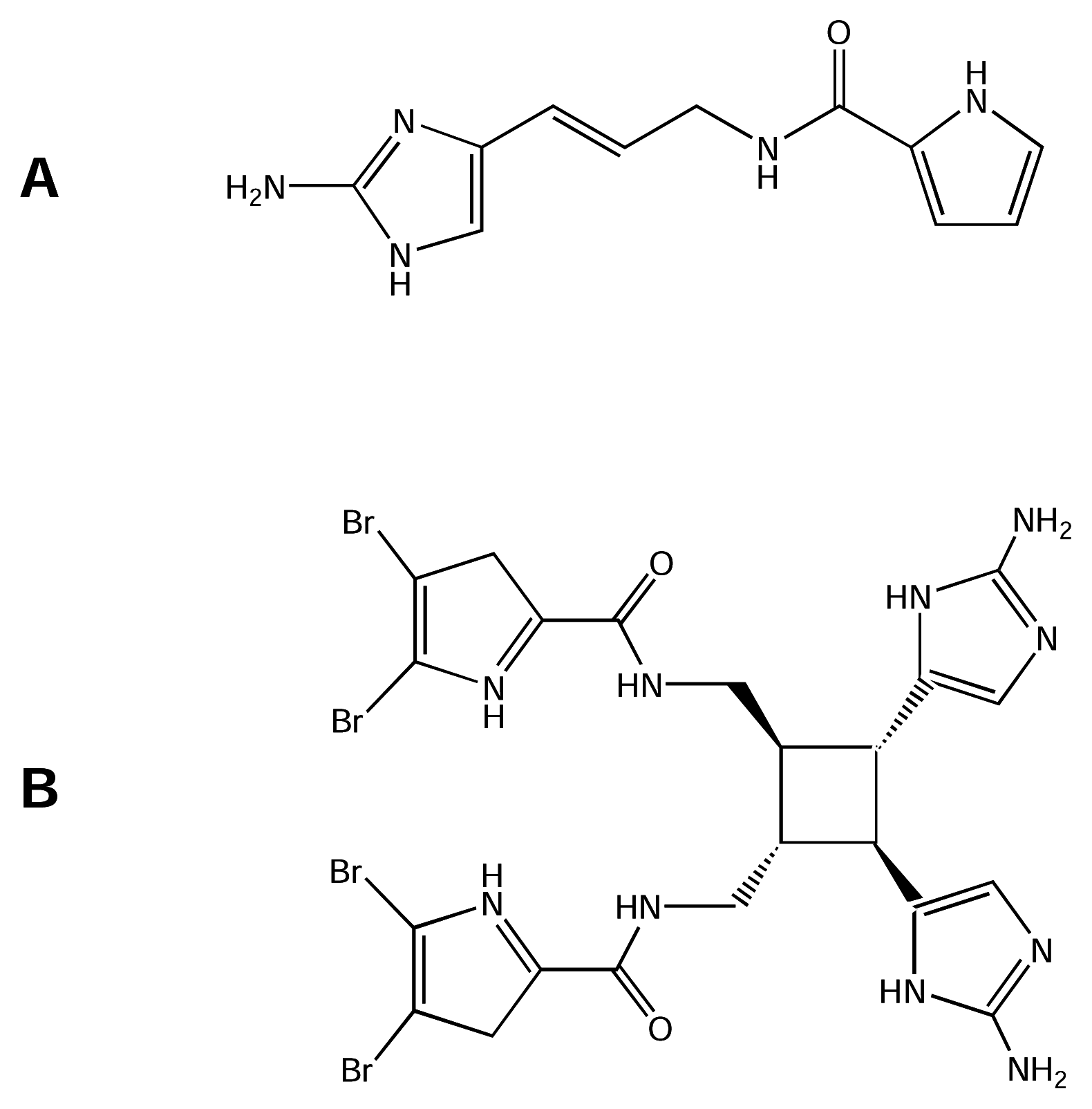
| N.I. | Inactivation gating modifier | ? | Clathrodin | Sponge (genus Agelas) |
| N.I. | Inactivation gating modifier | ? | Dibromosceptrin | Sponge (genus Agelas) |
| N.I. | Antagonist | ? | Jamaicamides A-C | Dinoflagellate Lyngbya majuscule |
| N.I. | Inactivation gating modifier | ? | Ostreotoxin-3 | Dinoflagellate Ostreopsis lenticularis |
| μ-Conotoxins | Conus species | # of residues | Nominal net charge (using R, K=1, H=+0.5, D=−1) | Major tissue or channel subtype(s) targeted | References |
|---|---|---|---|---|---|
| GIIIA | C. geographus | 22 | 6 | Rat muscle | [98] |
| GIIIB | C. geographus | 22 | 7 | Rat muscle, rat brain | [98] |
| GIIIC | C. geographus | 22 | 7 | Rat muscle | [33; 98] |
| PIIIA | C. purpurascens | 22 | 6.5 | Rat muscle, rat brain, human brain | [98; 134] |
| SmIIIA | C. stercusmuscarum | 22 | 5.5 | TTx-r (amphibian) skeletal muscle, sympathetic and dorsal root ganglia (DRG) neurons | [19; 73; 156] |
| SIIIA | C. striatus | 20 | 2.5 | TTx-r (amphibian) sympathetic and DRG neurons | [19] |
| KIIIA | C. kinoshitai | 16 | 3.5 | TTx-r (amphibian) sympathetic and DRG neurons | [19] |
| μO-Conotoxins | Conus species | Number of residues | Subtype Targeted | Reference |
|---|---|---|---|---|
| MrVIA | C. marmoreus | 31 | Inhibits TTx-r NaV channels rat dorsal root ganglia, TTx-s NaV channels, molluscan Ca2+ channels | [34; 95] |
| MrVIB | C. marmoreus | 31 | Inhibits TTx-r NaV channels rat dorsal root ganglia, TTx-s NaV channels, molluscan Ca2+ channels | [34; 95] |
| Unclassified Conotoxins | Conus species | Number of residues | Subtype Targeted | Reference |
|---|---|---|---|---|
| TVIIA | C. tulipa | 30 | Not established | [62] |
| GS | C. geographus | 34 | Muscle Na+ blocker receptor site 1 | [163] |
| δ-Conotoxin | Conus species | Natural prey | No. of residues | Subtype Targeted | Reference |
|---|---|---|---|---|---|
| PVIA | C. purpurascens | Fish | 29 | Vertebrate preparations only (including fish, frog and mammalian NaV1.2; NaV1.4 and NaV1.6). | [132; 157] [124; 145] |
| EVIA | C. ermineus | Fish | 32 | Amphibians and mammalian preparations (rNaV1.2, rNaV1.3 and rNaV1.6 but not muscular rNaV1.4 and cardiac hNaV1.5) | [5; 150] |
| SVIE | C. striatus | Fish | 31 | Diverse (Molluscan and Amphibian sympathic neurons) | [18; 157] |
| Am2766 | C. amadis | Molluscs | 26 | Diverse [molluscan and mammalian preparations (rNaV1.2)]. | [140] |
| TxVIA | C. textile | Molluscs | 27 | Molluscan subtype and rat brain neuronal membranes but not toxic to vertebrate system | [45–47; 65; 77] |
| GmVIA | C. gloriamaris | Molluscs | 29 | Diverse (molluscan subtype and mammalian NaV1.2 and NaV1.4 but not NaV1.6-mediated Na+ currents). | [61; 133] |
| NgVIA | C. nigropunctatus | Molluscs | 31 | Diverse (molluscan and more potent to mammalian preparations) | [47] |
Share and Cite
Al-Sabi, A.; McArthur, J.; Ostroumov, V.; French, R.J. Marine Toxins That Target Voltage-gated Sodium Channels. Mar. Drugs 2006, 4, 157-192. https://doi.org/10.3390/md403157
Al-Sabi A, McArthur J, Ostroumov V, French RJ. Marine Toxins That Target Voltage-gated Sodium Channels. Marine Drugs. 2006; 4(3):157-192. https://doi.org/10.3390/md403157
Chicago/Turabian StyleAl-Sabi, Ahmed, Jeff McArthur, Vitaly Ostroumov, and Robert J. French. 2006. "Marine Toxins That Target Voltage-gated Sodium Channels" Marine Drugs 4, no. 3: 157-192. https://doi.org/10.3390/md403157
APA StyleAl-Sabi, A., McArthur, J., Ostroumov, V., & French, R. J. (2006). Marine Toxins That Target Voltage-gated Sodium Channels. Marine Drugs, 4(3), 157-192. https://doi.org/10.3390/md403157