

Bioactivity of Marine-Derived Peptides and Proteins: A Review



Abstract
1. Introduction
2. Marine-Derived Peptides and Proteins: Sources and Characteristics
3. Production of Peptides from Marine Organisms

{kind=link}
{kind=link}
{kind=link}
| Enzyme (Protease) | References |
|---|---|
| Papain | [71,72,73] |
| Corolase | [74] |
| Thermolysin | [75] |
| Pancreatin | [76] |
| Ficin | [77] |
| Viscozyme | [78,79] |
| Trypsin | [73,80,81] |
| Pepsin | [64,82,83] |
| Chymotrypsin | [38,66] |
| Flavourzyme | [84,85] |
| Neutral protease | [86] |
| Alkaline protease | [14] |
| Subtilisin | [84] |
| Alcalase | [12,79,87,88,89] |
| Nucleicin | [90] |
| Orientase 22 BF | [90] |
| Neutrase | [12,39] |
| Protamex | [63,91,92] |
| Protease N | [63] |
| Corolase PP | [93] |
| Bromelain | [94] |
| Collagenase | [94] |
| Se-abzyme | [58] |
4. Bioactivities of Marine-Derived Protein and Peptides
4.1. Antioxidant Activity
4.2. Antimicrobial Activity
4.3. Anti-Inflammatory Effects
4.4. Antihypertensive Effects
4.5. Anti-Cancer Potential
4.6. Immunomodulatory Effects
4.7. Antihyperlipidemic Properties
4.8. Wound Healing and Skin Health
4.9. Antidiabetic Properties
4.10. Anticoagulation Potential
4.11. Miscellaneous
| Bioactivity | Marine Source | Method of Extraction | Protein/Peptide | Mode of Action | Reference |
|---|---|---|---|---|---|
| Antioxidant | Skipjack tuna (Katsuwonus pelamis) and SkinSkipjack Tuna (Katsuwonus pelamis) Roe |
|
|
| [225,226,227] |
| Siberian sturgeon (Acipenserbaerii) cartilage | Enzymatic hydrolysis | GEYGFE (700.70 Da), PSVSLT (602.67 Da), and IELFPGLP (942.12 Da) | DPPH and hydroxyl free radical scavenging activity. | [228] | |
| Antarctic krill (Euphausia superba) |
|
|
| [102,107] | |
| Seahorse (Hippocampus abdominalis) | Alcalase hydrolysis |
|
| [88,229] | |
| Pinctada fucata |
| Peptides | Exhibiting hydroxyl and superoxide radical-scavenging properties along with cellular antioxidant activity. | [230] | |
| Squid (Loligo formosana) head | Enzyme hydrolysis | Arg-Glu-Gly-Tyr-Phe-Lys | DPPH and ABTS scavenging activity. | [231] | |
| Atlantic sea cucumber (Cucumaria frondosa) | Enzymatic hydrolysis (Alcalase) | TEFHLL | Myeloperoxidase inhibition reduces oxidative stress in vitro. | [109] | |
| Cucumaria frondosa | Enzymatic hydrolysis (Alcalase, flavozyme, Corolase) | Protein hydrolysate | Hydroxyl radical scavenging activity, ferric reducing. | [74] | |
| Stripped weakfish by-products (skin and bone) | Alcalase and protamex hydrolysis | IELIEKPMGIF (1288.71 Da) RADLSRELEEISERL (1814.95 Da) | ABTS and hydroxyl radical scavenging activity. | [91] | |
| Miiuy croaker (Miichthys miiuy) | Enzymatic hydrolysis (papain) | FWKVV (612 Da) | DPPH, hydroxyl, and superoxide anion radical scavenging activity. | [232] | |
| Green algae (Dunaliella salina) | Ultrasound extraction followed by in vitro digestion | ILTKAAIEGK (1042 Da) IIYFQGK (867 Da) NDPSTVK (759 Da) TVRPPQR (852 Da) | DPPH radical scavenging activity. | [64] | |
| Sea cucumber (Acaudina molpadioides) |
|
|
| [39,165] | |
| Abalone (Haliotis discus hannai) | Water hydrolysis | ATPGDEG (752 Da) | DPPH radical, hydroxyl, ROS scavenging activity. | [233] | |
| Red microalgae (Palmaria palmata) | Enzyme hydrolysis (Corolase PP) | Ser-Asp-Ile-Thr-Arg-Pro-Gly-Gly-Asn-Met | Oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) activity. | [93] | |
| Shrimp shell discards | Enzymatic hydrolysis (Alcalase, trypsin, chymotrypsin, and pepsin) |
|
| [89,106] | |
| Sea squirt (Halocynthia roretzi) | Enzymatic hydrolysis (pepsin, Protamex) |
| DPPH and ABTS+ radical scavenging activity, Fe2+ chelating activity, ORAC, and reducing power. | [92,234] | |
| By-product of serra Spanish mackerel (Scomberomorus brasiliensis) | Enzymatic hydrolysis | Protein hydrolysate | DPPH and ABTS radical scavenging activity, ferric reducing ability, iron chelating ability. | [235] | |
| Antimicrobial | Octopus minor | -- | GWLIRGAIHAGKAIHGLIHRRRH | Exhibited antifungal activity against Candida albicans. | [123] |
| Half-Fin Anchovy (Setipinna taty) | -- | FEDQLR, ALERTF, and RHPEYAVSVLLR | Peptides increased H2O2 production and K+ leakage, causing irreversible membrane damage. | [128] | |
| Horseshoe crab (Limulus polyphemus) | -- | RRGCFRVCYRGFCFQRCR | Disrupts plasma membrane integrity, causing non-apoptotic cell death. | [127] | |
| Chlorella vulgaris | Enzymatic hydrolysis (pepsin) | Protein hydrolysate (62 kDa) | Inhibitory effect of 34.1% on E. coli growth. | [236] | |
| Mollusk (Cenchritis muricatus) | Solid-phase synthesis method | SRSELIVHQRLF | Fungistatic effect on Candida albicans. | [237] | |
| Antihypertentsive | By-catch shrimp (Oratosquilla woodmasoni) waste | Enzymatic hydrolysis (thermolysin and pepsin) | Asn-Gly-Val-Ala-Ala (431 Da) | Produces ACE-I inhibition peptide that could be utilized as an antihypertentsive. | [238] |
| Leathery sea squirt (Styela clava) | Enzymatic hydrolysis (pepsin) | LWHTH | Binds to the active site of ACE. | [239] | |
| Pearl oyster (Pinctada fucata) shell and pearl | Enzymatic hydrolysis
|
| ACE inhibitory activity. | [90,221] | |
| Squid (Todarodes pacificus) skin | Enzyamtic hydrolysis (alkaline protease) | FHGLPAK, IIAPPERKY, RGLPAYE, and VPSDVEF | Binds to the ACE active site via hydrogen bonding. | [240] | |
| Mackerel (Scomber japonicas) | Enzyamtic hydrolysis (papain) | PLITT | ACE is inhibited by hydrogen bonding interaction. | [72] | |
| Larimichthys crocea | Enzymatic hydrolysis (papain, trypsin) |
| Peptides bind to active or non-central sites of ACE through hydrogen bonding. | [73,241] | |
| Monkfish (Lophius litulon) swim bladders | Acid and enzyme hydrolysis (Alcalase, neutrase) | SEGPK (516.5 Da), FDGPY (597.6 Da), and SPGPW (542.6 Da) | Angiotensin-I-converting enzyme (ACE) inhibitory activities. | [12] | |
| Gracilariopsis chorda | Enzymatic hydrolysis (thermolysin) | IDHY, LRY, LVVER | ACE inhibitory activity. | [75] | |
| Arthrospira platensis | Enzymatic hydrolysis (papain, ficin, pepsin, or Alcalase) | Protein hydrolysate | ACE inhibitory activity. | [77] | |
| Laminaria digitata | Enzymatic hydrolysis (Viscozyme) |
| ACE inhibitory activity. | [242,243] | |
| Isochrysis zhanjiangensis | Enzymatic hydrolysis (trypsin) |
|
| [81,145] | |
| Blue mussels (Mytilus edulis) | Enzymatic hydrolysis | IK, YEGDP, WF, and SWISS | The inhibitory effect emerges from their strong interaction with the ACE active site via hydrogen bonding, electrostatic forces, and hydrophobic interactions. | [70] | |
| Shortfin scad (Decapterus macrosoma) waste | Alcalase hydrolysis | RGVGPVPAA (<3 kDa) | ACE inhibitory activity. | [244] | |
| Ruditapes philippinarum | Enzymatic hydrolysis (pepsin and pancreatin) | GRVSNCAA and TYLPVH | ACE inhibitory activity. | [107] | |
| Pufferfish (Takifugu flavidus) skin | Enzyme hydrolysis | PPLLFAAL (1 kDa) | Peptides exhibit strong binding affinity to angiotensin-converting enzyme (ACE). | [245] | |
| Chlorella pyrenoidosa | Enzyme hydrolysis (pepsin and trypsin) | FLKPLGSGK, LFVAEAIYK, and QIYTMGK | Peptides have potential interaction with ACE and block the enzyme. | [83] | |
| Atlantic salmon bone | Enzyme (trypsin) hydrolysis | Phe-Cys-Leu-Tyr-Glu- Leu-Ala-Arg | ACE inhibition occurs through hydrogen bond interactions between the peptide and the enzyme. | [246] | |
| Mesopelagic fish (Maurolicus muelleri and Meganyctiphanes norvegica) | Enzymatic hydrolysis | Protein hydrolysate | Acetylcholine esterase inhibition. | [247] | |
| Mazzaella japonica | Thermolysin | VDAHY, CPYDWV, SRIYNVKSNG, DFGVPGHEP, VSEGLD, YRD, SSNDYPI, GGPAT, TIMPHPR, YGDPDHY, and NLGN | ACE inhibition. | [248] | |
| Porphyra dioica | Enzyme hydrolysis | Thr-Tyr-Ile-Ala | ACE inhibitory activity. | [146] | |
| Spirulina (Arthrospira platensis) | Enzyme hydrolysis | IRDLDYY | Inhibits ACE through hydrophobic interactions and specific residue binding. | [5] | |
| Tilapia (Oreochromis niloticus) skin | -- | Leu-Ser-Gly-Tyr-Gly-Pro | Reducing oxidative stress and alleviating endothelial damage. | [182] | |
| Ulva intestinalis | Enzymatic hydrolysis (trypsin) | FGMPLDR (834.41 Da), MELVLR (759.43 Da) | Peptide activity relied on hydrogen bonds and Zn(II) interactions with ACE. | [249] | |
| Cobia (Rachycentron Canadum) skin | Enzymatic hydrolysis (Protamex and protease N) |
| ACE inhibitory activity. | [63] | |
| Deep-water pink shrimp (Parapenaeus longirostris) waste | Enzyme hydrolysis |
| ACE inhibitory activity. | [24,250] | |
| Bangia fuscopurpurea | Enzymatic hydrolysis (pepsin and trypsin) | ALLAGDPSVLEDR (1356 Da) VVGGTGPVDEWGIAGAR (1641 Da) | ACE inhibitory activity. | [251] | |
| Stylotella aurantium | Enzymatic hydrolysis (pepsin) | YR (337 Da) and IR (287 Da) | ACE inhibitory activity. | [252] | |
| Antiinflammation | Red algae (Amansia multifida) | Chemical hydrolysis | Protein (lectin) | Multi-faceted anti-inflammatory response: reducing edema, suppressing pro-inflammatory cytokines, limiting neutrophil migration, and enhancing antioxidant defenses through elevated glutathione levels. | [253] |
| Puffer fish (Lagocephalus guentheri) | Enzymatic hydrolysates (trypsin, Alcalase, and papain) |
|
| [71,254] | |
| Sturgeon (Acipenser schrenckii) cartilage | Enzymatic hydrolysis (papain and pancreatin); ethanol, hot pressure |
|
| [76,255] | |
| Sole fish (Psettodes erumei) | Enzymatic hydrolysis | MTQML (622 Da) | Suppresses NO and NF-κB complex expression while downregulating the NF-κB signaling cascade. | [256] | |
| Pearl oyster (Pinctada martensii) | Enzymatic hydrolysis (neutral protease) | TWP (402.19 Da), TAMY (484.19 Da), FPGA (390.19 Da) | Decreases NO and pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) while enhancing IL-10 production. | [134] | |
| Porphyridium sp. | Enzymatic hydrolysis (Viscozyme and Alcalase) | GVDYVRFF (1002.11 Da), AIPAAPAAPAGPKLY (1407.65 Da), LIHADPPGVGL (1088.25 Da) | Inhibits COX-1 and COX-2. | [78] | |
| Baijiao sea bass (Lateolabrax maculatus) | Ultrasonication aided extraction | AADGPMKGILGY (1192.38 Da), DAPAPPSQLEHIRAA | Suppresses NO production in LPS-induced macrophages. | [137] | |
| Marine crab (Charybdis natator) | Enzymatic hydrolysis (Alcalase, trypsin, and papain) | ESPVL (544 Da) | Inhibits iNOS and COX-2 expression at higher doses in LPS-induced macrophages. | [254] | |
| Herring (Clupea harengus) | Enzymatic hydrolysis | IVPAS (485.56 Da) | Reduces iNOS activation. | [257] | |
| Green mussel (Perna viridis) foot | Enzymatic hydrolysis (Alcalase) | EGLLGDVF (849.43 Da) | Reduces pro-inflammatory cytokines, NO, and COX-2 activation. | [87] | |
| Sea cucumber (Apostichopus japonicas, Acaudina leucoprocta) | Enzymatic hydrolysis | GPSGRP (569.60 Da), GPAGPR (553.60 Da), PQGETGA (658.64 Da), GFDGPEGPR (930.95 Da) | Suppresses TNF-α, IL-1β, and IL-6 by inactivating the MAPK/NF-κB pathway in an LPS-induced liver injury model. | [136] | |
| Crab (Charybdis natator) leg muscle | Enzymatic hydrolysis (trypsin, Alcalase, and papain) | LGLGAAVL (713.5 Da) | Suppression of LPS-mediated induction of COX-2 in RAW264.7 macrophage cells. | [133] | |
| Sardine (S. pilchardus) by-products | Neutral protease | Protein hydrolysate | Hindering the inflammation regulatin in the endothelial cells | [142] | |
| Anti-cancer | Cyclina sinensis | Ultrafiltration and chromatographic method | Ile-Leu-Tyr-Met-Pro (635.71 Da) | Induces anti-proliferative effects by promoting apoptosis by upregulating protein expression, attributed to the hydrophobic properties of the amino acids alanine (A) and leucine (L). | [159] |
| Catfish (Clarias gariepinus) | -- | HSDGIFTDSYSRYRKQMAVKKY-LAAVLGRRYRQRFRNK-NH2 | Exhibited dose-dependent cytotoxic activity and inhibition of cancer cell proliferation. | [258] | |
| Sinonovacula constricta | Enzymatic hydrolysis | Leu-Pro-Gly-Pro (382.46 Da) and Asp-Tyr-Val-Pro (492.53 Da) | Inhibited cell growth and reduced cell populations through the induction of apoptosis. | [259] | |
| Nile tilapia | -- | FIHHIIGGLFSAGKAIHRLIRRRRR | Induced cytotoxic activity, leading to cell death through necrosis. | [157] | |
| Spirulina platensis | Enzymatic hydrolysis | HVLSRAPR | Showed anti-proliferative activity against cancer cells while exhibiting low cytotoxicity toward normal cells. | [260] | |
| Antiallergic | Atlantic salmon (Salmo salar) viscera | Enzymatic (pepsin) hydrolysis | Thr-Pro-Glu-Val-His-Ile-Ala-Val-Aso-Lys-Phe | Inhibition of β-hexosaminidase release during IgE-mediated degranulation of RBL-2H3 cells. | [59] |
| Antidiabetic | Mesopelagic fish (Maurolicus muelleri and Meganyctiphanes norvegica) | Enzymatic hydrolysis | Protein hydrolysate | DPP-IV inhibitory activity. | [247] |
| Sardine pilchardus | Enzymatic hydrolysis (subtilisin, trypsin, and Flavourzyme) | NAPNPR, YACSVR | DPP-IV inhibitory activity. | [84] | |
| Spirulina platensis | -- | VPMPNK, RNPFVFAPTLLTVAAR, and LRSELAAWSR | α-amylase, α-glucosidase, and DPP-IV inhibition activities. | [184] | |
| Porphyra spp. | Enzymatic hydrolysis | GGSK, ELS | Inhibited α-amylase activity. | [261] | |
| Antiviral | Cod Skin | Hot-water hydrolysis | PGEKGPSGEAGTAGPPGTPGPQG (2163 Da) | Neuraminidase blocker to inhibit influenza A virus in MDCK cells. | [217] |
| Anticoagulation | Marine Bacillus velezensis Z01 | -- | Fibrinolytic protein | Ameliorated blood coagulation (aPTT, PT, TT, and FIB). | [204] |
| Marine bacterium (Pseudomonas aeruginosa KU1) | -- | Fibrinolytic protein (50 kDa) |
| [205] | |
| Spirulina (Arthrospira maxima) | -- | Phycocyanin protein | Prevented PAF and thrombin-induced platelet aggregation. | [210] | |
| Seaweed (Porphyra yezoensis) Nori | Enzymatic hydrolysis (pepsin) | NMEKGSSSVVSSRM (1.77 KDa) | Prolongation of APTT. | [196] | |
| Mussel (Mytilus edulis) | Enzymatic hydrolysis | Protein hydrolysates, KNAENELGEVYVR | Inhibited thrombin activity. | [200,262] | |
| Protein hydrolysates, ELEDSLDSER | Hindered thrombin action toward fibrinogen. | ||||
| Oyster (Crassostrea gigas) | Enzymatic hydrolysis (pepsin) | Protein hydrolysates, TARNEANVNIY | Prolongation aPTT and TT. | [35,198] | |
| Anticoagulant heptapeptides, NAESLRK |
| ||||
| Immunoregulator | Stolephorus chinensis | Enzymatic hydrolysis (pepsin, neutral protease, alkaline protease, trypsin, and papain) | YVMRF (715.4 Da) | Immunomodulatory activity. | [14] |
| Giant Croaker (Nibea japonica) | Neutral protease | Protein hydrolysate | Hydrolysate promotes the immune system through cell-mediated immunity. | [86] | |
| Antiobesity | Hammerhead shark (Sphyrna mokarran) skin | Enzymatic hydrolysis (pepsin and papain) | Collagen peptides | Attenuating high-fat diet–alcohol-induced hyperlipidemia. | [101] |
| Ark shell | Protein hydrolysate | LLRLTDL and GYALPCDCL | Modulating adipocyte differentiation. | [169] | |
| Miichthys miiuy muscle | Chemically synthethized | VIAPW and IRWWW | Inhibition of oleic acid (OA)-induced lipid accumulation in HepG2 cells along with a reduction in intracellular TG and TC levels. | [59] | |
| Spirulina platensis | Enzymatic hydrolysis (trypsin, alcalase, pepsin, papain, and Protamex) | NALKCCHSCPA, LNNPSVCDCDCMMKAAR, NPVWKRK, and CANPHELPNK | Inhibiotory effect on T3-L1 preadipocyte proliferation. | [263] | |
| Skate (Raja kenojei) skin | -- | Collagen peptide (1050 Da) | Suppression of fat accumulation and regulation of lipid metabolism. | [173] |
5. Applications of Marine Bioactive Peptides and Proteins
5.1. Food Industry
5.2. Pharmaceuticals
5.3. Cosmeceuticals
5.4. Nutraceuticals
6. Challenges and Future Direction
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Okeke, E.S.; Okagu, I.U.; Chukwudozie, K.; Ezike, T.C.; Ezeorba, T.P.C. Marine-Derived Bioactive Proteins and Peptides: A Review of Current Knowledge on Anticancer Potentials, Clinical Trials, and Future Prospects. Nat. Prod. Commun. 2024, 19, 1–19. [Google Scholar]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Rafieezadeh, D.; Esfandyari, G. Marine Bioactive Peptides with Anticancer Potential, a Narrative Review. Int. J. Biochem. Mol. Biol. 2024, 15, 118–126. [Google Scholar]
- Cunha, S.A.; Pintado, M.E. Bioactive Peptides Derived from Marine Sources: Biological and Functional Properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar]
- Anekthanakul, K.; Senachak, J.; Hongsthong, A.; Charoonratana, T.; Ruengjitchatchawalya, M. Natural ACE Inhibitory Peptides Discovery from Spirulina (Arthrospira platensis) Strain C1. Peptides 2019, 118, 170107. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Xiao, M.; Sudo, N.; Liu, Q. Bioactive Peptides of Marine Organisms: Roles in the Reduction and Control of Cardiovascular Diseases. Food Sci. Nutr. 2024, 12, 5271–5284. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Chang, J.S. Proteins and Bioactive Peptides from Algae: Insights into Antioxidant, Anti-Hypertensive, Anti-Diabetic, and Anti-Cancer Activities. Trends Food Sci. Technol. 2024, 145, 104352. [Google Scholar] [CrossRef]
- Venkatesan, J.; Anil, S.; Kim, S.K.; Shim, M.S. Marine Fish Proteins and Peptides for Cosmeceuticals: A Review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef]
- Macedo, M.W.F.S.; Cunha, N.B.d; Carneiro, J.A.; Costa, R.A.d.; Alencar, S.A.d.; Cardoso, M.H.; Franco, O.L.; Dias, S.C. Marine Organisms as a Rich Source of Biologically Active Peptides. Front. Mar. Sci. 2021, 8, 667764. [Google Scholar]
- Cermeño, M.; Kleekayai, T.; Amigo-Benavent, M.; Harnedy-Rothwell, P.; FitzGerald, R.J. Current Knowledge on the Extraction, Purification, Identification, and Validation of Bioactive Peptides from Seaweed. Electrophoresis 2020, 41, 1694–1717. [Google Scholar]
- Sila, A.; Bougatef, A. Antioxidant Peptides from Marine By-Products: Isolation, Identification and Application in Food Systems—A Review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Hu, Y.D.; Xi, Q.H.; Kong, J.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from the Collagens of Monkfish (Lophius litulon) Swim Bladders: Isolation, Characterization, Molecular Docking Analysis and Activity Evaluation. Mar. Drugs 2023, 21, 516. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, H.; Dinh, T.B.; Choi, S.; De Saeger, J.; Depuydt, S.; Brown, M.T.; Han, T. Commercial Potential of the Cyanobacterium Arthrospira maxima: Physiological and Biochemical Traits and the Purification of Phycocyanin. Biology 2022, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liang, R.; Li, D.; Jiang, C.; Lin, S. Evaluation of Sea Cucumber Peptides-Assisted Memory Activity and Acetylation Modification in Hippocampus of Test Mice Based on Scopolamine-Induced Experimental Animal Model of Memory Disorder. J. Funct. Foods 2020, 68, 68. [Google Scholar] [CrossRef]
- Ishak, N.H.; Sarbon, N.M. A Review of Protein Hydrolysates and Bioactive Peptides Deriving from Wastes Generated by Fish Processing. Food Bioprocess Technol. 2017, 11, 2–16. [Google Scholar] [CrossRef]
- Shahidi, F.; Santhiravel, S. Novel Marine Bioactives: Application in Functional Foods, Nutraceuticals, and Pharmaceuticals. J. Food Bioact. 2022, 19, 4–96. [Google Scholar]
- Portelinha, J.; Heilemann, K.; Jin, J.; Angeles-Boza, A.M. Unraveling the Implications of Multiple Histidine Residues in the Potent Antimicrobial Peptide Gaduscidin-1. J. Inorg. Biochem. 2021, 219, 111391. [Google Scholar] [CrossRef]
- Masso-Silva, J.A.; Diamond, G. Antimicrobial Peptides from Fish. Pharmaceuticals 2014, 7, 265–310. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Li, J.; Zhou, B. Peptides Derived from Rhopilema esculentum Hydrolysate Exhibit Angiotensin Converting Enzyme (ACE) Inhibitory and Antioxidant Abilities. Molecules 2014, 19, 13587–13602. [Google Scholar] [CrossRef]
- Lim, Y.S.; Ok, Y.J.; Hwang, S.Y.; Kwak, J.Y.; Yoon, S. Marine Collagen as a Promising Biomaterial for Biomedical Applications. Mar. Drugs 2019, 17, 467. [Google Scholar] [CrossRef]
- Farooq, S.; Ahmad, M.I.; Zheng, S.; Ali, U.; Li, Y.; Shixiu, C.; Zhang, H. A Review on Marine Collagen: Sources, Extraction Methods, Colloids Properties, and Food Applications. Collagen Leather 2024, 6, 11. [Google Scholar] [CrossRef]
- Zheng, S.L.; Wang, Y.Z.; Zhao, Y.Q.; Chi, C.F.; Zhu, W.Y.; Wang, B. High Fischer Ratio Oligopeptides from Hard-Shelled Mussel: Preparation and Hepatoprotective Effect against Acetaminophen-Induced Liver Injury in Mice. Food Biosci. 2023, 53, 53. [Google Scholar]
- Babita, S.; Krishan, D.S.; Caresma, C. Seafood Bioactives for Health and Wellness. Int. J. Sci. Res. 2020, 9, 1588–1598. [Google Scholar]
- Krichen, F.; Sila, A.; Caron, J.; Kobbi, S.; Nedjar, N.; Miled, N.; Blecker, C.; Besbes, S.; Bougatef, A. Identification and Molecular Docking of Novel ACE Inhibitory Peptides from Protein Hydrolysates of Shrimp Waste. Eng. Life Sci. 2018, 18, 682–691. [Google Scholar]
- Liu, Z.; Li, M.; Yi, Q.; Wang, L.; Song, L. The Neuroendocrine-Immune Regulation in Response to Environmental Stress in Marine Bivalves. Front. Physiol. 2018, 9, 1456. [Google Scholar] [CrossRef]
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and Immunomodulatory Properties and Applications of Marine-Derived Proteins and Peptides. Mar. Drugs 2019, 17, 350. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide Identification in a Salmon Gelatin Hydrolysate with Antihypertensive, Dipeptidyl Peptidase IV Inhibitory and Antioxidant Activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Fitzgerald, R.J. Bioactive Proteins and Peptides from Macroalgae, Fish, Shellfish and Marine Processing Waste. In Marine Proteins and Peptides: Biological Activities and Applications; Kim, S.K., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 5–39. [Google Scholar]
- Wang, Z.; Hu, S.; Gao, Y.; Ye, C.; Wang, H. Effect of Collagen-Lysozyme Coating on Fresh-Salmon Fillets Preservation. LWT–Food Sci. Technol. 2017, 75, 59–64. [Google Scholar]
- Oh, R.; Lee, M.J.; Kim, Y.O.; Nam, B.H.; Kong, H.J.; Kim, J.W.; Park, J.Y.; Seo, J.K.; Kim, D.G. Myticusin-Beta, Antimicrobial Peptide from the Marine Bivalve, Mytilus coruscus. Fish Shellfish Immunol. 2020, 99, 342–352. [Google Scholar] [CrossRef]
- Maselli, V.; Galdiero, E.; Salzano, A.M.; Scaloni, A.; Maione, A.; Falanga, A.; Naviglio, D.; Guida, M.; Di Cosmo, A.; Galdiero, S. OctoPartenopin: Identification and Preliminary Characterization of a Novel Antimicrobial Peptide from the Suckers of Octopus vulgaris. Mar. Drugs 2020, 18, 380. [Google Scholar] [CrossRef]
- Yu, H.H.; Wu, L.Y.; Hsu, P.L.; Lee, C.W.; Su, B.C. Marine Antimicrobial Peptide Epinecidin-1 Inhibits Proliferation Induced by Lipoteichoic Acid and Causes Cell Death in Non-Small Cell Lung Cancer Cells via Mitochondria Damage. Probiotics Antimicrob. Proteins 2024, 16, 1724–1733. [Google Scholar] [CrossRef] [PubMed]
- Kandyliari, A.; Golla, J.P.; Chen, Y.; Papandroulakis, N.; Kapsokefalou, M.; Vasiliou, V. Antiproliferative Activity of Protein Hydrolysates Derived from Fish By-Products on Human Colon and Breast Cancer Cells. Proc. Nutr. Soc. 2020, 79, E282. [Google Scholar]
- Kang, H.K.; Choi, M.C.; Seo, C.H.; Park, Y. Therapeutic Properties and Biological Benefits of Marine-Derived Anticancer Peptides. Int. J. Mol. Sci. 2018, 19, 919. [Google Scholar] [CrossRef]
- Chandika, P.; Tennakoon, P.; Kim, T.-H.; Kim, S.-C.; Je, J.-Y.; Kim, J.-I.; Lee, B.; Ryu, B.; Kang, H.W.; Kim, H.-W.; et al. Marine Biological Macromolecules and Chemically Modified Macromolecules; Potential Anticoagulants. Mar. Drugs 2022, 20, 654. [Google Scholar] [CrossRef] [PubMed]
- Quitério, E.; Soares, C.; Ferraz, R.; Delerue-Matos, C.; Grosso, C. Marine Health-Promoting Compounds: Recent Trends for Their Characterization and Human Applications. Foods 2021, 10, 3100. [Google Scholar] [CrossRef]
- Dagnino-Leone, J.; Figueroa, C.P.; Castañeda, M.L.; Youlton, A.D.; Vallejos-Almirall, A.; Agurto-Muñoz, A.; Pavón Pérez, J.; Agurto-Muñoz, C. Phycobiliproteins: Structural Aspects, Functional Characteristics, and Biotechnological Perspectives. Comput. Struct. Biotechnol. J. 2022, 20, 1506–1527. [Google Scholar]
- Gong, P.X.; Wang, B.K.; Wu, Y.C.; Li, Q.Y.; Qin, B.W.; Li, H.J. Release of Antidiabetic Peptides from Stichopus japonicus by Simulated Gastrointestinal Digestion. Food Chem. 2020, 315, 126273. [Google Scholar] [CrossRef]
- Jin, H.X.; Xu, H.P.; Li, Y.; Zhang, Q.W.; Xie, H. Preparation and Evaluation of Peptides with Potential Antioxidant Activity by Microwave-Assisted Enzymatic Hydrolysis of Collagen from Sea Cucumber Acaudina molpadioides Obtained from Zhejiang Province in China. Mar. Drugs 2019, 17, 169. [Google Scholar] [CrossRef]
- Chim-Chi, Y.; Olivera-Castillo, L.; Betancur-Ancona, D.; Chel-Guerrero, L. Protein Hydrolysate Fractions from Sea Cucumber (Isostichopus badionotus) Inhibit Angiotensin-Converting Enzyme. J. Aquat. Food Prod. Technol. 2017, 26, 1199–1209. [Google Scholar]
- Jayasanta, I.; Sathish, N.; Patterson, J. Identification of Bioactive Peptides in Mussel Species of Kanyakumari Coast. Asian J. Biotechnol. 2020, 12, 75–86. [Google Scholar]
- Sruthy, K.; Chaithanya, E.; Sathyan, N.; Anilkumar, P.; Singh, I.; Philip, R. Isolation, Purification, and Characterization of Antimicrobial Peptides from Indian Ruff, Psenopsis cyanea (Alcock, 1890). Int. J. Res. Biol. Sci. 2015, 5, 1–6. [Google Scholar]
- Bai, C.; Wei, Q.; Ren, X. Selective Extraction of Collagen Peptides with High Purity from Cod Skins by Deep Eutectic Solvents. ACS Sustain. Chem. Eng. 2017, 5, 7220–7227. [Google Scholar]
- Arumugam, V.; Venkatesan, M.; Ramachandran, S.; Sundaresan, U. Bioactive Peptides from Marine Ascidians and Future Drug Development—A Review. Int. J. Pept. Res. Ther. 2018, 24, 13–18. [Google Scholar]
- Srivastava, A.; Mishra, V. Marine Peptides Act as Novel Chemotherapeutic Agents. J. Microbiol. Exp. 2018, 6, 267–270. [Google Scholar]
- Kendler, S.; Kobbenes, S.M.M.; Jakobsen, A.N.; Mukhatov, K.; Lerfall, J. The Application of Microwave and Ultrasound Technologies for Extracting Collagen from European Plaice By-Products. Front. Sustain. Food Syst. 2023, 7, 1257635. [Google Scholar] [CrossRef]
- Al Khawli, F.; Pateiro, M.; Domínguez, R.; Lorenzo, J.M.; Gullón, P.; Kousoulaki, K.; Ferrer, E.; Berrada, H.; Barba, F.J. Innovative Green Technologies of Intensification for Valorization of Seafood and Their By-Products. Mar. Drugs 2019, 17, 689. [Google Scholar] [CrossRef] [PubMed]
- Lakmal, C.H.; Samarakoon, K.W.; Jeon, Y.J. Enzyme-Assisted Extraction to Prepare Bioactive Peptides from Microalgae. In Marine Algae Extracts; Kim, S.K., Chojnacka, K., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 305–318. [Google Scholar]
- Melgosa, R.; Trigueros, E.; Sanz, M.T.; Cardeira, M.; Rodrigues, L.; Fernández, N.; Matias, A.A.; Bronze, M.R.; Marques, M.; Paiva, A.; et al. Supercritical CO2 and Subcritical Water Technologies for the Production of Bioactive Extracts from Sardine (Sardina pilchardus) Waste. J. Supercrit. Fluids 2020, 164, 104943. [Google Scholar]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from Fish By-Product Protein Hydrolysates and Its Functional Properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar]
- Petrova, I.; Tolstorebrov, I.; Eikevik, T.M. Production of Fish Protein Hydrolysates Step by Step: Technological Aspects, Equipment Used, Major Energy Costs and Methods of Their Minimizing. Int. Aquat. Res. 2018, 10, 223–241. [Google Scholar]
- Wisuthiphaet, N.; Klinchan, S.; Kongruang, S. Production of Fish Protein Hydrolysates by Acid and Enzymatic Hydrolysis. Int. J. Appl. Sci. Technol. 2015, 4, 261–270. [Google Scholar] [CrossRef]
- Wang, J.; Xu, M.; Liang, R.; Zhao, M.; Zhang, Z.; Li, Y. Oral Administration of Marine Collagen Peptides Prepared from Chum Salmon (Oncorhynchus keta) Improves Wound Healing Following Cesarean Section in Rats. Food Nutr. Res. 2015, 59, 26411. [Google Scholar] [CrossRef] [PubMed]
- Villamil, O.; Vaquiro, H.; Solanilla, J.F. Fish Viscera Protein Hydrolysates: Production, Potential Applications, and Functional and Bioactive Properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef]
- Jack, O.C.; Meaney, S.; Williams, G.A.; Hayes, M. Extraction of Protein from Four Different Seaweeds Using Three Different Physical Pre-Treatment Strategies. Molecules 2020, 25, 2005. [Google Scholar] [CrossRef] [PubMed]
- Abdelhedi, O.; Nasri, M. Basic and Recent Advances in Marine Antihypertensive Peptides: Production, Structure-Activity Relationship, and Bioavailability. Trends Food Sci. Technol. 2019, 88, 543–557. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Emire, S.A. Production and Processing of Antioxidant Bioactive Peptides: A Driving Force for the Functional Food Market. Heliyon 2020, 6, e04765. [Google Scholar] [CrossRef]
- Slizyte, R.; Rommi, K.; Mozuraityte, R.; Eck, P.; Five, K.; Rustad, T. Bioactivities of Fish Protein Hydrolysates from Defatted Salmon Backbones. Biotechnol. Rep. 2016, 11, 99–109. [Google Scholar] [CrossRef]
- Wang, K.; Siddanakoppalu, P.N.; Ahmed, I.; Pavase, T.R.; Lin, H.; Li, Z. Purification and Identification of Anti-Allergic Peptide from Atlantic Salmon (Salmo salar) Byproduct Enzymatic Hydrolysates. J. Funct. Foods 2020, 72, 104084. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef]
- Shin, D.; Byun, W.S.; Moon, K.; Kwon, Y.; Bae, M.; Um, S.; Lee, S.K.; Oh, D.C. Coculture of marine Streptomyces sp. with Bacillus sp. produces a new piperazic acid-bearing cyclic peptide. Front. Chem. 2018, 6, 498. [Google Scholar] [CrossRef]
- Raju, T.S. Proteolysis of Proteins. In Co and Post-Translational Modifications of Therapeutic Antibodies and Proteins; Raju, T.S., Ed.; Wiley: Hoboken, NJ, USA, 2019; pp. 183–202. [Google Scholar]
- Lin, Y.H.; Chen, C.A.; Tsai, J.S.; Chen, G.W. Preparation and Identification of Novel Antihypertensive Peptides from the In Vitro Gastrointestinal Digestion of Marine Cobia Skin Hydrolysates. Nutrients 2019, 11, 1351. [Google Scholar] [CrossRef]
- Xia, E.; Zhai, L.; Huang, Z.; Liang, H.; Yang, H.; Song, G.; Li, W.; Tang, H. Optimization and Identification of Antioxidant Peptide from Underutilized Dunaliella salina Protein: Extraction, In Vitro Gastrointestinal Digestion, and Fractionation. Biomed. Res. Int. 2019, 2019, 6424651. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Sun, N.; Dong, L.; Gao, Y.; Lin, S. Production of Bioactive Peptides from Sea Cucumber and Its Potential Health Benefits: A Comprehensive Review. J. Agric. Food Chem. 2022, 70, 7607–7625. [Google Scholar] [CrossRef]
- Lee, H.G.; Kim, H.S.; Oh, J.Y.; Lee, D.S.; Yang, H.W.; Kang, M.C.; Kim, E.A.; Kang, N.; Kim, J.; Heo, S.J.; et al. Potential Antioxidant Properties of Enzymatic Hydrolysates from Stichopus japonicus Against Hydrogen Peroxide-Induced Oxidative Stress. Antioxidants 2021, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, K.; Inbaraj, B.S.; Chen, B.H. Recent Developments on Production, Purification, and Biological Activity of Marine Peptides. Food Res. Int. 2021, 147, 110468. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant Protein-Derived Antioxidant Peptides: Isolation, Identification, Mechanism of Action, and Application in Food Systems: A Review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Bo, W.; Chen, L.; Qin, D.; Geng, S.; Li, J.; Mei, H.; Li, B.; Liang, G. Application of Quantitative Structure-Activity Relationship to Food-Derived Peptides: Methods, Situations, Challenges, and Prospects. Trends Food Sci. Technol. 2021, 114, 176–188. [Google Scholar] [CrossRef]
- Suo, S.K.; Zhao, Y.Q.; Wang, Y.M.; Pan, X.Y.; Chi, C.F.; Wang, B. Seventeen Novel Angiotensin Converting Enzyme (ACE) Inhibitory Peptides from the Protein Hydrolysate of Mytilus edulis: Isolation, Identification, Molecular Docking Study, and Protective Function on HUVECs. Food Funct. 2022, 13, 7831–7846. [Google Scholar] [CrossRef]
- Balde, A.; Ghosh, P.; Aishwarya, P.; Vaishnavi, V.; Nazeer, R.A. Utilization of Diamondback Puffer (Lagocephalus guentheri) Biomass for the Production of Bioactive Oligopeptides and Their Inflammation Suppressing Effects In Vitro. Biocatal. Agric. Biotechnol. 2024, 58, 103155. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, H.; Wen, Y.; Yuan, W.; Chen, H.; Lin, L.; Guo, F.; Zheng, Z.P.; Zhao, C. The Novel Angiotensin-I-Converting Enzyme Inhibitory Peptides from Scomber japonicus Muscle Protein Hydrolysates: QSAR-Based Screening, Molecular Docking, Kinetic and Stability Studies. Food Chem. 2024, 447, 138873. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, C.; Huang, B.; Chen, Y.; Liu, Z.; Chen, H.; Chen, J. Biodirected Screening and Preparation of Larimichthys crocea Angiotensin-I-Converting Enzyme-Inhibitory Peptides by a Combined In Vitro and In Silico Approach. Molecules 2024, 29, 1134. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. Antioxidant and ACE-Inhibitory Activity of Protein Hydrolysates Produced from Atlantic Sea Cucumber (Cucumaria frondosa). Molecules 2023, 28, 5263. [Google Scholar] [CrossRef] [PubMed]
- Mune Mune, M.A.; Miyabe, Y.; Shimizu, T.; Matsui, W.; Kumagai, Y.; Kishimura, H. Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda. Mar. Drugs 2023, 21, 49. [Google Scholar] [CrossRef]
- Yuan, L.; Chu, Q.; Yang, B.; Zhang, W.; Sun, Q.; Gao, R. Purification and Identification of Anti-Inflammatory Peptides from Sturgeon (Acipenser schrenckii) Cartilage. Food Sci. Hum. Wellness 2023, 12, 2175–2183. [Google Scholar]
- Villaró, S.; Jiménez-Márquez, S.; Musari, E.; Bermejo, R.; Lafarga, T. Production of Enzymatic Hydrolysates with In Vitro Antioxidant, Antihypertensive, and Antidiabetic Properties from Proteins Derived from Arthrospira platensis. Food Res. Int. 2023, 163, 112270. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of Bioactive Peptides from Porphyridium sp. and Assessment of Their Potential for Use in the Prevention of Hypertension, Inflammation, and Pain. Mar. Drugs 2023, 21, 422. [Google Scholar] [CrossRef] [PubMed]
- Aurino, E.; Mora, L.; Marzocchella, A.; Kuchendorf, C.M.; Ackermann, B.; Hayes, M. Functional and Bioactive Benefits of Selected Microalgal Hydrolysates Assessed In Silico and In Vitro. Mar. Drugs 2025, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Suttisuwan, R.; Phunpruch, S.; Saisavoey, T.; Sangtanoo, P.; Thongchul, N.; Karnchanatat, A. Isolation and Characterization of Anti-Inflammatory Peptides Derived from Trypsin Hydrolysis of Microalgae Protein (Synechococcus sp. VDW). Food Biotechnol. 2019, 33, 303–324. [Google Scholar] [CrossRef]
- Pei, Y.; Cai, S.; Ryu, B.; Zhou, C.; Hong, P.; Qian, Z.J. An ACE Inhibitory Peptide from Isochrysis zhanjiangensis Exhibits Antihypertensive Effect via Anti-Inflammation and Anti-Apoptosis in HUVEC and Hypertensive Rats. J. Funct. Foods 2022, 92, 105061. [Google Scholar]
- Veeruraj, A.; Liu, L.; Zheng, J.; Wu, J.; Arumugam, M. Evaluation of Astaxanthin Incorporated Collagen Film Developed from the Outer Skin Waste of Squid Doryteuthis singhalensis for Wound Healing and Tissue Regenerative Applications. Mater. Sci. Eng. C 2019, 95, 29–42. [Google Scholar]
- Li, Y.; Aiello, G.; Fassi, E.M.A.; Boschin, G.; Bartolomei, M.; Bollati, C.; Roda, G.; Arnoldi, A.; Grazioso, G.; Lammi, C. Investigation of Chlorella pyrenoidosa Protein as a Source of Novel Angiotensin I-Converting Enzyme (ACE) and Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Peptides. Nutrients 2021, 13, 1624. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Production and Identification of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides from Discarded Sardine pilchardus Protein. Food Chem. 2020, 328, 127096. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, T.R.L.; Dave, D.; Shahidi, F. Antioxidant Potential and Physicochemical Properties of Protein Hydrolysates from Body Parts of North Atlantic Sea Cucumber (Cucumaria frondosa). Food Prod. Process. Nutr. 2021, 3, 3. [Google Scholar]
- Yu, F.; He, K.; Dong, X.; Zhang, Z.; Wang, F.; Tang, Y.; Chen, Y.; Ding, G. Immunomodulatory Activity of Low Molecular-Weight Peptides from Nibea japonica Skin in Cyclophosphamide-Induced Immunosuppressed Mice. J. Funct. Foods 2020, 68, 103888. [Google Scholar] [CrossRef]
- Joshi, I.; Nazeer, R.A. EGLLGDVF: A Novel Peptide from Green Mussel Perna viridis Foot Exerts Stability and Anti-Inflammatory Effects on LPS-Stimulated RAW264.7 Cells. Protein Pept. Lett. 2020, 27, 851–859. [Google Scholar] [PubMed]
- Lee, H.G.; Nagahawatta, D.P.; Yang, F.; Jayawardhana, H.H.A.C.K.; Liyanage, N.M.; Lee, D.S.; Lee, J.M.; Yim, M.J.; Ko, S.C.; Kim, J.Y.; et al. Antioxidant Potential of Hydrolysate-Derived Seahorse (Hippocampus abdominalis) Peptide: Protective Effects Against AAPH-Induced Oxidative Damage In Vitro and In Vivo. Food Chem. 2023, 407, 135130. [Google Scholar] [PubMed]
- Singh, A.; Kadam, D.; Gautam, A.R.; Rengasamy, K.R.R.; Aluko, R.E.; Benjakul, S. Angiotensin-I-Converting Enzyme and Renin Inhibitions by Antioxidant Shrimp Shell Protein Hydrolysate and Ultrafiltration Peptide Fractions. Food Biosci. 2024, 60, 104524. [Google Scholar] [CrossRef]
- Sasaki, C.; Tamura, S.; Tohse, R.; Fujita, S.; Kikuchi, M.; Asada, C.; Nakamura, Y. Isolation and Identification of an Angiotensin I-Converting Enzyme Inhibitory Peptide from Pearl Oyster (Pinctada fucata) Shell Protein Hydrolysate. Process Biochem. 2019, 77, 137–142. [Google Scholar]
- Lima, K.O.; da Costa de Quadros, C.; Rocha, M.; de Lacerda, J.T.J.G.; Juliano, M.A.; Dias, M.; Mendes, M.A.; Prentice, C. Bioactivity and Bioaccessibility of Protein Hydrolysates from Industrial Byproducts of Stripped Weakfish (Cynoscion guatucupa). LWT Food Sci. Technol. 2019, 111, 48–413. [Google Scholar]
- Ma, R.; Chen, Q.; Dai, Y.; Huang, Y.; Hou, Q.; Huang, Y.; Zhong, K.; Huang, Y.; Gao, H.; Bu, Q. Identification of Novel Antioxidant Peptides from Sea Squirt (Halocynthia roretzi) and Its Neuroprotective Effect in 6-OHDA-Induced Neurotoxicity. Food Funct. 2022, 13, 6008–6021. [Google Scholar]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Fractionation and Identification of Antioxidant Peptides from an Enzymatically Hydrolysed Palmaria palmata Protein Isolate. Food Res. Int. 2017, 100, 416–422. [Google Scholar]
- Hossain, A.; Dave, D.; Shahidi, F. Northern Sea Cucumber (Cucumaria frondosa): A Potential Candidate for Functional Food, Nutraceutical, and Pharmaceutical Sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Januka, T.; Kumari, D.; de Camargo, A.C.; Shahidi, F. Phenolic Antioxidants of Bael Fruit Herbal Tea and Effects on Postprandial Glycemia and Plasma Antioxidant Status in Healthy Adults. J. Food Bioact. 2020, 11, 75–83. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative Stress Mitigation by Antioxidants—An Overview on Their Chemistry and Influences on Health Status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar]
- Shrivastava, A.; Mishra, S.P.; Pradhan, S.; Choudhary, S.; Singla, S.; Zahra, K.; Aggarwal, L.M. An Assessment of Serum Oxidative Stress and Antioxidant Parameters in Patients Undergoing Treatment for Cervical Cancer. Free Radic. Biol. Med. 2021, 167, 29–35. [Google Scholar]
- Taira, Y.; Yamashita, T.; Bian, Y.; Shang, J.; Matsumoto, N.; Sasaki, R.; Tadokoro, K.; Nomura, E.; Tsunoda, K.; Omote, Y.; et al. Antioxidative Effects of a Novel Dietary Supplement Neumentix in a Mouse Stroke Model. J. Stroke Cerebrovasc. Dis. 2020, 29, 104818. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Shahidi, F. Bioactive Compounds and Bioactive Properties of Chaga (Inonotus obliquus) Mushroom: A Review. J. Food Bioact. 2020, 12, 9–75. [Google Scholar]
- Peña-Bautista, C.; Lopez-Cuevas, R.; Cuevas, A.; Baquero, M.; Chafer-Pericas, C. Lipid Peroxidation Biomarkers Correlation with Medial Temporal Atrophy in Early Alzheimer Disease. Neurochem. Int. 2019, 129, 104519. [Google Scholar] [CrossRef]
- Vijayan, D.K.; Perumcherry Raman, S.; Dara, P.K.; Jacob, R.M.; Mathew, S.; Rangasamy, A.; Chandragiri Nagarajarao, R. In Vivo Anti-Lipidemic and Antioxidant Potential of Collagen Peptides Obtained from Great Hammerhead Shark Skin Waste. J. Food Sci. Technol. 2021, 59, 1140–1151. [Google Scholar]
- Wang, Q.; Shi, J.; Zhong, H.; Abdullah; Zhuang, J.; Zhang, J.; Wang, J.; Zhang, X.; Feng, F. High-Degree Hydrolysis Sea Cucumber Peptides Improve Exercise Performance and Exert Antifatigue Effect via Activating the NRF2 and AMPK Signaling Pathways in Mice. J. Funct. Foods 2021, 86, 104677. [Google Scholar]
- Jegani, K.T.; Balde, A.; Nazeer, R.A. A Review on Anti-Inflammatory and Antioxidant Peptides Derived from Marine Organisms: Mechanism of Action and Therapeutic Applications. Food Biosci. 2025, 63, 105745. [Google Scholar]
- Pratama, I.S.; Putra, Y.; Pangestuti, R.; Kim, S.K.; Siahaan, E.A. Bioactive Peptides-Derived from Marine By-Products: Development, Health Benefits and Potential Application in Biomedicine. Fish. Aquat. Sci. 2022, 25, 357–379. [Google Scholar] [CrossRef]
- Safari, R.; Yaghoubzadeh, Z. Antioxidant Activity of Bioactive Peptides Extracted from Sea Cucumber (Holothuria leucospilata). Int. J. Pept. Res. Ther. 2020, 26, 2393–2398. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Shahidi, F. Bioactive Peptides from Shrimp Shell Processing Discards: Antioxidant and Biological Activities. J. Funct. Foods 2017, 34, 7–17. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhao, G.X.; Suo, S.K.; Wang, Y.M.; Chi, C.F.; Wang, B. Purification, Identification, Activity Evaluation, and Stability of Antioxidant Peptides from Alcalase Hydrolysate of Antarctic Krill (Euphausia superba) Proteins. Mar. Drugs 2021, 19, 347. [Google Scholar] [CrossRef] [PubMed]
- Guanghua, X.; Zhang, X.; Zhenghua, D.; Xuanri, S. Comparative Study on the Antioxidant Activity of Peptides from Pearl Oyster (Pinctada martensii) Mantle Type V Collagen and Tilapia (Oreochromis niloticus) Scale Type I Collagen. J. Ocean Univ. China 2017, 16, 1175–1182. [Google Scholar]
- Zhang, Y.; He, S.; Bonneil, É.; Simpson, B.K. Generation of Antioxidative Peptides from Atlantic Sea Cucumber Using Alcalase Versus Trypsin: In Vitro Activity, De Novo Sequencing, and In Silico Docking for In Vivo Function Prediction. Food Chem. 2020, 306, 125581. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Isolation, Purification and Identification of Three Novel Antioxidative Peptides from Patin (Pangasius sutchi) Myofibrillar Protein Hydrolysates. LWT-Food Sci. Technol. 2015, 60, 452–461. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Sohn, J.H.; Kim, J.S.; Choi, J.S. Identification and Characterization of Novel Antioxidant Peptides from Mackerel (Scomber japonicus) Muscle Protein Hydrolysates. Food Chem. 2020, 323, 126809. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, X.; Zhuang, Y. Purification and Characterization of Novel Antioxidant Peptides from Enzymatic Hydrolysates of Tilapia (Oreochromis niloticus) Skin Gelatin. Peptides 2012, 38, 13–21. [Google Scholar] [CrossRef]
- Saidi, S.; Saoudi, M.; Ben Amar, R. Valorisation of Tuna Processing Waste Biomass: Isolation, Purification and Characterisation of Four Novel Antioxidant Peptides from Tuna By-Product Hydrolysate. Environ. Sci. Pollut. Res. 2018, 25, 17383–17392. [Google Scholar] [CrossRef]
- Kim, N.Y.; Jung, H.Y.; Kim, J.K. Identification and Characterization of a Novel Heptapeptide from Mackerel By-Product Hydrolysate and Its Potential as a Functional Fertilizer Component. J. Chromatogr. B 2021, 1180, 122881. [Google Scholar] [CrossRef] [PubMed]
- Terriente-Palacios, C.; Rubiño, S.; Hortós, M.; Peteiro, C.; Castellari, M. Taurine, Homotaurine, GABA, and Hydrophobic Amino Acids Content Influence in Vitro Antioxidant and SIRT1 Modulation Activities of Enzymatic Protein Hydrolysates from Algae. Sci. Rep. 2022, 12, 20832. [Google Scholar]
- Zhang, L.; Yang, D.; Wang, Q.; Yuan, Z.; Wu, H.; Pei, D.; Cong, M.; Li, F.; Ji, C.; Zhao, J. A Defensin from Clam Venerupis philippinarum: Molecular Characterization, Localization, Antibacterial Activity, and Mechanism of Action. Dev. Comp. Immunol. 2015, 51, 29–38. [Google Scholar]
- Wang, K.; Han, L.; Tan, Y.; Hong, H.; Fan, H.; Luo, Y. Novel Hypocholesterolemic Peptides Derived from Silver Carp Muscle: The Modulatory Effects on Enterohepatic Cholesterol Metabolism In Vitro and In Vivo. J. Agric. Food Chem. 2023, 71, 5565–5575. [Google Scholar] [PubMed]
- Kumar, P.; Kannan, M.; ArunPrasanna, V.; Vaseeharan, B.; Vijayakumar, S. Proteomics Analysis of Crude Squid Ink Isolated from Sepia esculenta for Their Antimicrobial, Antibiofilm, and Cytotoxic Properties. Microb. Pathog. 2018, 116, 345–350. [Google Scholar] [PubMed]
- Djellouli, M.; López-Caballero, M.E.; Arancibia, M.Y.; Karam, N.; Martínez-Alvarez, O. Antioxidant and Antimicrobial Enhancement by Reaction of Protein Hydrolysates Derived from Shrimp By-Products with Glucosamine. Waste Biomass Valorization 2020, 11, 2491–2505. [Google Scholar] [CrossRef]
- Rodriguez, A.; Gmelin, N.; Martell-Huguet, E.M.; Gonz, M.; Alp, D.; Alba, A.; Vazquez, A.A.; Grieshober, M.; Spellerberg, B.; Stenger, S.; et al. Identification and Characterization of Three New Antimicrobial Peptides from the Marine Mollusk. Int. J. Mol. Sci. 2023, 24, 3852. [Google Scholar] [CrossRef]
- Ma, L.; Ye, X.; Sun, P.; Xu, P.; Wang, L.; Liu, Z.; Huang, X.; Bai, Z.; Zhou, C. Antimicrobial and Antibiofilm Activity of the EeCentrocin 1 Derived Peptide EC1-17KV via Membrane Disruption. EBioMedicine 2020, 55, 102775. [Google Scholar]
- Solstad, R.G.; Johansen, C.; Stensvåg, K.; Strøm, M.B.; Haug, T. Structure-Activity Relationship Studies of Shortened Analogues of the Antimicrobial Peptide EeCentrocin 1 from the Sea Urchin Echinus esculentus. J. Pept. Sci. 2020, 26, e3233. [Google Scholar]
- Nikapitiya, C.; Dananjaya, S.H.S.; Chandrarathna, H.P.S.U.; de Zoysa, M.; Whang, I. Octominin: A Novel Synthetic Anticandidal Peptide Derived from Defense Protein of Octopus minor. Mar. Drugs 2020, 18, 56. [Google Scholar] [CrossRef]
- Rajapaksha, D.C.; Jayathilaka, E.H.T.T.; Edirisinghe, S.L.; Nikapitiya, C.; Lee, J.; Whang, I.; De Zoysa, M. Octopromycin: Antibacterial and Antibiofilm Functions of a Novel Peptide Derived from Octopus minor against Multidrug-Resistant Acinetobacter baumannii. Fish Shellfish Immunol. 2021, 117, 82–94. [Google Scholar] [CrossRef]
- Zheng, L.; Li, Y.; Wang, J.; Pan, Y.; Chen, J.; Zheng, W.; Lin, L. Antibacterial and Antiparasitic Activities Analysis of a Hepcidin-Like Antimicrobial Peptide from Larimichthys crocea. Acta Oceanol. Sin. 2020, 39, 129–139. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, Q.J.; Qiao, Y.; Chen, J.; Li, M.Y. The Host Defense Peptide β-Defensin Confers Protection Against Vibrio anguillarum in Ayu, Plecoglossus altivelis. Dev. Comp. Immunol. 2020, 103, 103511. [Google Scholar] [CrossRef] [PubMed]
- Marggraf, M.B.; Panteleev, P.V.; Emelianova, A.A.; Sorokin, M.I.; Bolosov, I.A.; Buzdin, A.A.; Kuzmin, D.V.; Ovchinnikova, T.V. Cytotoxic Potential of the Novel Horseshoe Crab Peptide Polyphemusin III. Mar. Drugs 2018, 16, 466. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, R.; Song, R. Novel Antibacterial Peptides Isolated from the Maillard Reaction Products of Half-Fin Anchovy (Setipinna taty) Hydrolysates/Glucose and Their Mode of Action in Escherichia coli. Mar. Drugs 2019, 17, 47. [Google Scholar] [CrossRef]
- Sila, A.; Nedjar-Arroume, N.; Hedhili, K.; Chataigné, G.; Balti, R.; Nasri, M.; Dhulster, P.; Bougatef, A. Antibacterial Peptides from Barbel Muscle Protein Hydrolysates: Activity Against Some Pathogenic Bacteria. LWT Food Sci. Technol. 2014, 55, 183–188. [Google Scholar] [CrossRef]
- Jemil, I.; Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Aristoy, M.C.; Toldrá, F.; Nasri, M. Novel Bioactive Peptides from Enzymatic Hydrolysate of Sardinelle (Sardinella aurita) Muscle Proteins Hydrolyzed by Bacillus subtilis A26 Proteases. Food Res. Int. 2017, 100, 121–133. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Domper Arnal, M.J.; Hijos-Mallada, G.; Lanas, A. Gastrointestinal and Cardiovascular Adverse Events Associated with NSAIDs. Expert Opin. Drug Saf. 2022, 21, 373–384. [Google Scholar] [CrossRef]
- Narayanasamy, A.; Balde, A.; Raghavender, P.; Shashanth, D.; Abraham, J.; Joshi, I.; Nazeer, R.A. Isolation of Marine Crab (Charybdis natator) Leg Muscle Peptide and Its Anti-Inflammatory Effects on Macrophage Cells. Biocatal. Agric. Biotechnol. 2020, 25, 101577. [Google Scholar] [CrossRef]
- Shen, J.; Zeng, M.; Huang, P.; Chen, B.; Xia, Z.; Cao, Y.; Miao, J. Purification and Activity Evaluation of Novel Anti-Inflammatory Peptides from Pearl Oyster (Pinctada martensii) Hydrolysates. Food Funct. 2023, 14, 4242–4253. [Google Scholar] [CrossRef] [PubMed]
- Joshi, I.; Sudhakar, S.; Nazeer, R.A. Anti-Inflammatory Properties of Bioactive Peptide Derived from Gastropod Influenced by Enzymatic Hydrolysis. Appl. Biochem. Biotechnol. 2016, 180, 1128–1140. [Google Scholar]
- Wan, H.; Han, J.; Tang, S.; Bao, W.; Lu, C.; Zhou, J.; Ming, T.; Li, Y.; Su, X. Comparisons of Protective Effects Between Two Sea Cucumber Hydrolysates Against Diet-Induced Hyperuricemia and Renal Inflammation in Mice. Food Funct. 2020, 11, 1074–1086. [Google Scholar]
- Chen, J.; Bai, W.; Cai, D.; Yu, Z.; Xu, B. Characterization and Identification of Novel Anti-Inflammatory Peptides from Baijiao Sea Bass (Lateolabrax maculatus). LWT-Food Sci. Technol. 2021, 147, 111521. [Google Scholar]
- Ahn, C.B.; Cho, Y.S.; Je, J.Y. Purification and Anti-Inflammatory Action of Tripeptide from Salmon Pectoral Fin Byproduct Protein Hydrolysate. Food Chem. 2015, 168, 151–156. [Google Scholar] [PubMed]
- Gao, R.; Shu, W.; Shen, Y.; Sun, Q.; Bai, F.; Wang, J.; Li, D.; Li, Y.; Jin, W.; Yuan, L. Sturgeon Protein-Derived Peptides Exert Anti-Inflammatory Effects in LPS-Stimulated RAW264.7 Macrophages via the MAPK Pathway. J. Funct. Foods 2020, 72, 104044. [Google Scholar]
- Park, Y.R.; Park, C.I.; Soh, Y. Antioxidant and Anti-Inflammatory Effects of NCW Peptide from Clam Worm (Marphysa sanguinea). J. Microbiol. Biotechnol. 2020, 30, 1387–1394. [Google Scholar] [PubMed]
- Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Karnchanatat, A. Anti-Inflammatory Action of Two Novel Peptides Derived from Peanut Worms (Sipunculus nudus) in Lipopolysaccharide-Induced RAW264.7 Macrophages. Food Funct. 2020, 11, 552–560. [Google Scholar]
- Vieira, E.F.; Van Camp, J.; Ferreira, I.M.; Grootaert, C. Protein Hydrolysate from Canned Sardine and Brewing By-Products Improves TNF-α-Induced Inflammation in an Intestinal–Endothelial Co-Culture Cell Model. Eur. J. Nutr. 2018, 57, 2275–2286. [Google Scholar] [CrossRef]
- Feng, S.; Limwachiranon, J.; Luo, Z.; Shi, X.; Ru, Q. Preparation and Purification of Angiotensin-Converting Enzyme Inhibitory Peptides from Hydrolysate of Shrimp (Litopenaeus vannamei) Shell Waste. Int. J. Food Sci. Technol. 2016, 51, 1610–1617. [Google Scholar]
- Deng, Z.; Liu, Y.; Wang, J.; Wu, S.; Geng, L.; Sui, Z.; Zhang, Q. Antihypertensive Effects of Two Novel Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Gracilariopsis lemaneiformis (Rhodophyta) in Spontaneously Hypertensive Rats (SHRs). Mar. Drugs 2018, 16, 299. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tan, L.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Mechanism Analysis of a Novel Angiotensin-I-Converting Enzyme Inhibitory Peptide from Isochrysis zhanjiangensis Microalgae for Suppressing Vascular Injury in Human Umbilical Vein Endothelial Cells. J. Agric. Food Chem. 2020, 68, 4411–4423. [Google Scholar]
- Cermeno, M.; Stack, J.; Tobin, P.R. Peptide Identification from a Porphyra dioica Protein Hydrolysate with Antioxidant, Angiotensin Converting Enzyme, and Dipeptidyl Peptidase IV Inhibitory Activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chen, G.W.; Yeh, C.H.; Song, H.; Tsai, J.S. Purification and Identification of Angiotensin I-Converting Enzyme Inhibitory Peptides and the Antihypertensive Effect of Chlorella sorokiniana Protein Hydrolysates. Nutrients 2018, 10, 1397. [Google Scholar] [CrossRef]
- Sadegh, V.M.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Angiotensin-I Converting Enzyme (ACE) Inhibitory and Anti-Hypertensive Effect of Protein Hydrolysate from Actinopyga lecanora (Sea Cucumber) in Rats. Mar. Drugs 2016, 14, 176. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Abdul Hamid, A.; Saari, N. Purification and Characterization of Angiotensin Converting Enzyme-Inhibitory Peptides Derived from Stichopus horrens: Stability Study Against the ACE and Inhibition Kinetics. J. Funct. Foods 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Shaik, M.I.; Sarbon, N.M. A Review on Purification and Characterization of Anti-Proliferative Peptides Derived from Fish Protein Hydrolysate. Food Rev. Int. 2022, 38, 1389–1409. [Google Scholar]
- Malaguti, M.; Dinelli, G.; Leoncini, E.; Bregola, V.; Bosi, S.; Cicero, A.; Hrelia, S. Bioactive Peptides in Cereals and Legumes: Agronomical, Biochemical and Clinical Aspects. Int. J. Mol. Sci. 2014, 15, 21120–21135. [Google Scholar] [CrossRef]
- Méresse, S.; Fodil, M.; Fleury, F.; Chénais, B. Fucoxanthin, a Marine-Derived Carotenoid from Brown Seaweeds and Microalgae: A Promising Bioactive Compound for Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 9273. [Google Scholar] [CrossRef]
- Li, M.; Zhou, M.; Wei, Y.; Jia, F.; Yan, Y.; Zhang, R.; Cai, M.; Gu, R. The Beneficial Effect of Oyster Peptides and Oyster Powder on Cyclophosphamide-Induced Reproductive Impairment in Male Rats: A Comparative Study. J. Food Biochem. 2020, 44, e13468. [Google Scholar] [CrossRef]
- Fuente, D.L.; Berrada, H.; Barba, F.J. Marine Resources and Cancer Therapy: From Current Evidence to Challenges for Functional Foods Development. Curr. Opin. Food Sci. 2022, 44, 100805. [Google Scholar] [CrossRef]
- Hamzeh, A.; Rezaei, M.; Khodabandeh, S.; Motamedzadegan, A.; Noruzinia, M. Antiproliferative and Antioxidative Activities of Cuttlefish (Sepia pharaonis) Protein Hydrolysates as Affected by Degree of Hydrolysis. J. Food Meas. Charact. 2018, 12, 721–727. [Google Scholar] [CrossRef]
- Eghtedari, M.; Jafari Porzani, S.; Nowruzi, B. Anticancer Potential of Natural Peptides from Terrestrial and Marine Environments: A Review. Phytochem. Lett. 2021, 42, 87–103. [Google Scholar] [CrossRef]
- Ting, C.H.; Chen, J.Y. Nile Tilapia-Derived TP4 Shows Broad Cytotoxicity Toward Non-Small-Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 506. [Google Scholar] [CrossRef] [PubMed]
- Su, B.C.; Wu, T.H.; Hsu, C.H.; Chen, J.Y. Distribution of Positively Charged Amino Acid Residues in Antimicrobial Peptide Epinecidin-1 Is Crucial for in Vitro Glioblastoma Cytotoxicity and Its Underlying Mechanisms. Chem. Biol. Interact. 2020, 315, 108904. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Y.; Ye, L.; Tang, Y.; Ding, G.; Zhang, X.; Yang, Z. A Novel Anti-Proliferative Pentapeptide (ILYMP) Isolated from Cyclina sinensis Protein Hydrolysate Induces Apoptosis of DU-145 Prostate Cancer Cells. Mol. Med. Rep. 2018, 18, 771–778. [Google Scholar]
- Qu, B.; Yuan, J.; Liu, X.; Zhang, S.; Ma, X.; Lu, L. Anticancer Activities of Natural Antimicrobial Peptides from Animals. Front. Microbiol. 2024, 14, 1321386. [Google Scholar] [CrossRef]
- Schweikart, K.; Guo, L.; Shuler, Z.; Abrams, R.; Chiao, E.T.; Kolaja, K.L.; Davis, M. The Effects of Jaspamide on Human Cardiomyocyte Function and Cardiac Ion Channel Activity. Toxicol. Vitr. 2013, 27, 745–751. [Google Scholar] [CrossRef]
- Hansen, I.K.Ø.; Isaksson, J.; Poth, A.G.; Hansen, K.; Andersen, A.J.C.; Richard, C.S.M.; Blencke, H.M.; Stensvåg, K.; Craik, D.J.; Haug, T. Isolation and Characterization of Antimicrobial Peptides with Unusual Disulfide Connectivity from the Colonial Ascidian Synoicum turgens. Mar. Drugs 2020, 18, 51. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Z.; Chen, Y.; Wu, T.; Fersht, V.; Jin, Y.; Meng, J.; Zhang, M. Sea Cucumber Peptides Inhibit the Malignancy of NSCLC by Regulating miR-378a-5p Targeted TUSC2. Food Funct. 2021, 12, 12362–12371. [Google Scholar] [CrossRef]
- Wargasetia, T.L.; Ratnawati, H.; Widodo, N.; Widyananda, M.H. Bioinformatics Study of Sea Cucumber Peptides as Antibreast Cancer Through Inhibiting the Activity of Overexpressed Proteins (EGFR, PI3K, AKT1, and CDK4). Cancer Inform. 2021, 20, 11769351211031864. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Lin, S.J.; Yang, Z.S.; Jin, H.X. Preparation of Antioxidant Peptide by Microwave-Assisted Hydrolysis of Collagen and Its Protective Effect Against H2O2-Induced Damage of RAW264.7 Cells. Mar. Drugs 2019, 17, 642. [Google Scholar] [CrossRef]
- Cai, N.; Luo, W.; Yao, L.; Li, X.; Wang, Z.; Xu, H.; Li, H.; Hu, Z.; Bao, W.; Xu, X. Activation of Murine RAW264.7 Macrophages by Oligopeptides from Sea Cucumber (Apostichopus japonicus) and Its Molecular Mechanisms. J. Funct. Foods 2020, 75, 104229. [Google Scholar]
- He, L.X.; Zhang, Z.F.; Sun, B.; Chen, Q.H.; Liu, R.; Ren, J.W.; Wang, J.B.; Li, Y. Sea Cucumber (Codonopsis pilosula) Oligopeptides: Immunomodulatory Effects Based on Stimulating Th Cells, Cytokine Secretion, and Antibody Production. Food Funct. 2016, 7, 1208–1216. [Google Scholar]
- Du, X.; Lian, F.; Li, Y.; Li, D.; Wu, D.; Feng, Q.; Feng, Z.; Li, Y.; Bu, G.; Meng, F.; et al. Peptides from Colochirus robustus Enhance Immune Function via Activating CD3zeta- and ZAP-70-Mediated Signaling in C57BL/6 Mice. Int. J. Mol. Sci. 2017, 18, 2110. [Google Scholar] [PubMed]
- Ahn, C.B.; Je, J.Y. Anti-Adipogenic Peptides from Ark Shell Protein Hydrolysate: Purification, Identification and Anti-Adipogenic Effect. Process Biochem. 2021, 109, 143–147. [Google Scholar]
- Woo, M.; Seol, B.G.; Kang, K.H.; Choi, Y.H.; Cho, E.J.; Noh, J.S. Effects of Collagen Peptides from Skate (Raja kenojei) Skin on Improvements of the Insulin Signaling Pathway via Attenuation of Oxidative Stress and Inflammation. Food Funct. 2020, 11, 2017–2025. [Google Scholar] [CrossRef]
- Villarruel-López, A.; Ascencio, F.; Nuño, K. Microalgae, a Potential Natural Functional Food Source–A Review. Pol. J. Food Nutr. Sci. 2017, 67, 251–263. [Google Scholar]
- Pavlicevic, M.; Maestri, E.; Marmiroli, M. Marine Bioactive Peptides—An Overview of Generation, Structure and Application with a Focus on Food Sources. Mar. Drugs 2020, 18, 424. [Google Scholar] [CrossRef]
- Woo, M.; Song, Y.O.; Kang, K.H.; Noh, J.S. Anti-Obesity Effects of Collagen Peptide Derived from Skate (Raja kenojei) Skin Through Regulation of Lipid Metabolism. Mar. Drugs 2018, 16, 306. [Google Scholar] [CrossRef]
- Lee, E.J.; Hur, J.; Ham, S.A.; Jo, Y.; Lee, S.; Choi, M.J.; Seo, H.G. Fish Collagen Peptide Inhibits the Adipogenic Differentiation of Preadipocytes and Ameliorates Obesity in High Fat Diet-Fed Mice. Int. J. Biol. Macromol. 2017, 104, 281–286. [Google Scholar] [PubMed]
- Lassoued, I.; Trigui, M.; Ghlissi, Z.; Nasri, R.; Jamoussi, K.; Kessis, M.; Sahnoun, Z.; Rebai, T.; Boualga, A.; Lamri-Senhadji, M.; et al. Evaluation of Hypocholesterolemic Effect and Antioxidant Activity of Boops boops Proteins in Cholesterol-Fed Rats. Food Funct. 2014, 5, 1224–1231. [Google Scholar]
- Athmani, N.; Dehiba, F.; Allaoui, A.; Barkia, A.; Bougatef, A.; Lamri-Senhadji, M.Y.; Nasri, M.; Boualga, A. Sardina pilchardus and Sardinella aurita Protein Hydrolysates Reduce Cholesterolemia and Oxidative Stress in Rats Fed High Cholesterol Diet. J. Exp. Integr. Med. 2015, 5, 47–54. [Google Scholar]
- Hu, Z.; Yang, P.; Zhou, C.; Li, S.; Hong, P. Marine Collagen Peptides from the Skin of Nile Tilapia (Oreochromis niloticus): Characterization and Wound Healing Evaluation. Mar. Drugs 2017, 15, 102. [Google Scholar] [CrossRef]
- Geahchan, S.; Baharlouei, P.; Rahman, A. Marine Collagen: A Promising Biomaterial for Wound Healing, Skin Anti-Aging, and Bone Regeneration. Mar. Drugs 2022, 20, 61. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, K.; Li, B.; Hou, H. Effects of Oral Administration of Peptides with Low Molecular Weight from Alaska Pollock (Theragra chalcogramma) on Cutaneous Wound Healing. J. Funct. Foods 2018, 48, 682–691. [Google Scholar] [CrossRef]
- Pozzolini, M.; Millo, E.; Oliveri, C.; Mirata, S.; Salis, A.; Damonte, G.; Arkel, M.; Scarfì, S. Elicited ROS Scavenging Activity, Photoprotective, and Wound-Healing Properties of Collagen-Derived Peptides from the Marine Sponge Chondrosia reniformis. Mar. Drugs 2018, 16, 465. [Google Scholar] [CrossRef]
- Dang, Q.F.; Liu, H.; Yan, J.Q.; Liu, C.S.; Liu, Y.; Li, J.; Li, J.J. Characterization of Collagen from Haddock Skin and Wound Healing Properties of Its Hydrolysates. Biomater. Sci. 2015, 10, 015022. [Google Scholar] [CrossRef]
- Chen, J.; Gao, K.; Liu, S.; Wang, S.; Elango, J.; Bao, B.; Dong, J.; Liu, N.; Wu, W. Fish Collagen Surgical Compress Repairing Characteristics on Wound Healing Process in Vivo. Mar. Drugs 2019, 17, 33. [Google Scholar] [CrossRef]
- Melotti, L.; Martinello, T.; Perazzi, A.; Iacopetti, I.; Ferrario, C.; Sugni, M.; Sacchetto, R.; Patruno, M. A Prototype Skin Substitute, Made of Recycled Marine Collagen, Improves the Skin Regeneration of Sheep. Animals 2021, 11, 1219. [Google Scholar] [CrossRef]
- Hu, S.; Fan, X.; Qi, P.; Zhang, X. Identification of Anti-Diabetes Peptides from Spirulina platensis. J. Funct. Foods 2019, 56, 333–341. [Google Scholar]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Purification and Identification of Dipeptidyl Peptidase (DPP) IV Inhibitory Peptides from the Macroalga Palmaria palmata. Food Chem. 2015, 172, 400–406. [Google Scholar] [PubMed]
- Cian, R.E.; Nardo, A.E.; Garzón, A.G.; Añón, M.C.; Drago, S.R. Identification and in silico Study of a Novel Dipeptidyl Peptidase IV Inhibitory Peptide Derived from Green Seaweed Ulva spp. Hydrolysates. LWT Food Sci. Technol. 2022, 154, 112738. [Google Scholar]
- Balansa, W. Exploring Metabolites of Green Algae Caulerpa spp. to Discover Putative Inhibitors of DPP-4, a New Antidiabetic Target Protein: Molecular Docking Compounds from Marine Green Algae. J. Ilm. Platax 2023, 11, 333–346. [Google Scholar]
- Ou, Y.; Ren, Z.; Wang, J.; Yang, X. Phycocyanin Ameliorates Alloxan-Induced Diabetes Mellitus in Mice: Involved in Insulin Signaling Pathway and GK Expression. Chem. Biol. Interact. 2016, 247, 49–54. [Google Scholar]
- Sadek, K.M.; Lebda, M.A.; Nasr, S.M.; Shoukry, M. Spirulina platensis Prevents Hyperglycemia in Rats by Modulating Gluconeogenesis and Apoptosis via Modification of Oxidative Stress and MAPK Pathways. Biomed. Pharmacother. 2017, 92, 1085–1094. [Google Scholar] [CrossRef]
- Aissaoui, O.; Amiali, M.; Bouzid, N.; Belkacemi, K.; Bitam, A. Effect of Spirulina platensis Ingestion on the Abnormal Biochemical and Oxidative Stress Parameters in the Pancreas and Liver of Alloxan-Induced Diabetic Rats. Pharm. Biol. 2017, 55, 1304–1312. [Google Scholar]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; O’Keeffe, M.B.; Le Gouic, A.V.; Allsopp, P.J.; McSorley, E.M.; Sharkey, S.; Whooley, J.; McGovern, B.; O’Harte, F.P.M.; et al. Identification and Characterisation of Peptides from a Boarfish (Capros aper) Protein Hydrolysate Displaying In Vitro Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory and Insulinotropic Activity. Food Res. Int. 2020, 131, 108989. [Google Scholar]
- Neves, A.C.; Harnedy, P.A.; FitzGerald, R.J. Angiotensin Converting Enzyme and Dipeptidyl Peptidase-IV Inhibitory, and Antioxidant Activities of a Blue Mussel (Mytilus edulis) Meat Protein Extract and Its Hydrolysates. J. Aquat. Food Prod. Technol. 2016, 25, 1221–1233. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Atlantic Salmon (Salmo salar) Co-Product-Derived Protein Hydrolysates: A Source of Antidiabetic Peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar]
- Yang, L.; Zhang, L.; Yan, L.; Zheng, H.; Lu, P.; Chen, J.; Dai, J.; Sun, H.; Xu, Y.; Yang, T. Stability Assessment of a New Antithrombotic Small Peptide, Arg-Gly-Asp-Trp-Arg (RGDWR), and Its Derivative. Biotechnol. Lett. 2017, 39, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, S.; Huma, N.; Pasha, I.; Shahid, M.; Xiao, H. Angiotensin-Converting Enzyme-Inhibitory and Antithrombotic Activities of Soluble Peptide Extracts from Buffalo and Cow Milk Cheddar Cheeses. Int. J. Dairy Technol. 2017, 70, 380–388. [Google Scholar] [CrossRef]
- Indumathi, P.; Mehta, A. A Novel Anticoagulant Peptide from the Nori Hydrolysate. J. Funct. Foods 2016, 20, 606–617. [Google Scholar] [CrossRef]
- Cheng, S.; Tu, M.; Chen, H.; Xu, Z.; Wang, Z.; Liu, H.; Zhao, G.; Zhu, B.; Du, M. Identification and Inhibitory Activity against α-Thrombin of a Novel Anticoagulant Peptide Derived from Oyster (Crassostrea gigas) Protein. Food Funct. 2018, 9, 6391–6400. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wang, Y.; Chen, H.; Liu, H.; Wang, L.; Battino, M.; Yao, X.; Zhu, B.; Du, M. Anticoagulant Dodecapeptide Suppresses Thrombosis In Vivo by Inhibiting the Thrombin Exosite-I Binding Site. J. Agric. Food Chem. 2021, 69, 10920–10931. [Google Scholar] [CrossRef]
- Cheng, S.; Wu, D.; Yuan, L.; Liu, H.; Ei-Seedi, H.R.; Du, M. Crassostrea gigas-Based Bioactive Peptide Protected Thrombin-Treated Endothelial Cells Against Thrombosis and Cell Barrier Dysfunction. J. Agric. Food Chem. 2022, 70, 9664–9673. [Google Scholar] [CrossRef]
- Qiao, M.; Tu, M.; Chen, H.; Mao, F.; Yu, C.; Du, M. Identification and In Silico Prediction of Anticoagulant Peptides from the Enzymatic Hydrolysates of Mytilus edulis Proteins. Int. J. Mol. Sci. 2018, 19, 2100. [Google Scholar] [CrossRef]
- Qiao, M.; Tu, M.; Wang, Z.; Mao, F.; Chen, H.; Qin, L.; Du, M. Identification and Antithrombotic Activity of Peptides from Blue Mussel (Mytilus edulis) Protein. Int. J. Mol. Sci. 2018, 19, 138. [Google Scholar] [CrossRef]
- Cheng, S.; Yuan, L.; Li-Gao, R.; Chen, S.; Li, H.; Du, M. Nutrition and Cardiovascular Disease: The Potential Role of Marine Bioactive Proteins and Peptides in Thrombosis Prevention. J. Agric. Food Chem. 2024, 72, 6815–6832. [Google Scholar] [CrossRef]
- Harish, B.S.; Uppuluri, K.B. Potential Anticoagulant Activity of Trypsin Inhibitor Purified from an Isolated Marine Bacterium Oceanimonas sp. BPMS22 and Its Kinetics. Mar. Biotechnol. 2018, 20, 780–791. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, H.; Yu, B.; Chen, G.; Liang, Z. Purification and Characterization of a Fibrinolytic Enzyme from Marine Bacillus velezensis Z01 and Assessment of Its Therapeutic Efficacy In Vivo. Microorganisms 2022, 10, 843. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Haridas, M.; Abdulhameed, S. A Novel Fibrinolytic Enzyme from Marine Pseudomonas aeruginosa KU1 and Its Rapid In Vivo Thrombolysis with Little Haemolysis. Int. J. Biol. Macromol. 2020, 162, 470–479. [Google Scholar]
- Dhamodharan, D.; Naine, J.S.; Keziah, M.S.; Devi, S.C. Novel Fibrinolytic Protease Producing Streptomyces radiopugnans VITSD8 from Marine Sponges. Mar. Drugs 2019, 17, 164. [Google Scholar] [CrossRef]
- Joji, K.; Santhiagu, A.; Salim, N. Computational Modeling of Culture Media for Enhanced Production of Fibrinolytic Enzyme from Marine Bacterium Fictibacillus sp. Strain SKA27 and In Vitro Evaluation of Fibrinolytic Activity. Biotech 2019, 9, 323. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Belur, P.D.; Rai, P.; Rekha, P.D. Production of Fibrinolytic Enzyme by the Marine Isolate Serratia marcescens subsp. sakuensis and Its In Vitro Anticoagulant and Thrombolytic Potential. J. Pure Appl. Microbiol. 2017, 11, 1987–1998. [Google Scholar] [CrossRef]
- Silva, P.E.D.C.E.; Barros, R.C.; Albuquerque, W.W.C.; Brandão, R.M.P.; Bezerra, R.P.; Porto, A.L.F. In Vitro Thrombolytic Activity of a Purified Fibrinolytic Enzyme from Chlorella vulgaris. J. Chromatogr. B 2018, 1092, 524–529. [Google Scholar]
- Koukouraki, P.; Tsoupras, A.; Sotiroudis, G.; Demopoulos, C.A.; Sotiroudis, T.G. Antithrombotic Properties of Spirulina Extracts Against Platelet-Activating Factor and Thrombin. Food Biosci. 2020, 37, 100686. [Google Scholar]
- Zhao, Y.; Dong, Y.; Ge, Q.; Cui, P.; Sun, N.; Lin, S. Neuroprotective Effects of NDEELNK from Sea Cucumber Ovum Against Scopolamine-Induced PC12 Cell Damage Through Enhancing Energy Metabolism and Upregulation of the PKA/BDNF/NGF Signaling Pathway. Food Funct. 2021, 12, 7676–7687. [Google Scholar]
- Zhao, Y.; Lu, Z.; Xu, X.; Sun, N.; Lin, S. Sea Cucumber-Derived Peptide Attenuates Scopolamine-Induced Cognitive Impairment by Preventing Hippocampal Cholinergic Dysfunction and Neuronal Cell Death. J. Agric. Food Chem. 2022, 70, 567–576. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, S.; Sun, N.; Zhu, B.; Lin, S. Neuroprotective Function of a Novel Hexapeptide QMDDQ from Shrimp via Activation of the PKA/CREB/BDNF Signaling Pathway and Its Structure-Activity Relationship. J. Agric. Food Chem. 2020, 68, 6759–6769. [Google Scholar]
- Yang, M.; Xu, Z.; Wu, D.; Dong, Y.; Wang, Z.; Du, M. Characterizations and the Mechanism Underlying Osteogenic Activity of Peptides from Enzymatic Hydrolysates of Stichopus japonicus. J. Agric. Food Chem. 2021, 69, 15611–15623. [Google Scholar] [PubMed]
- Lin, L.; Yang, K.; Zheng, L.; Zhao, M.; Sun, W.; Zhu, Q.; Liu, S. Anti-Aging Effect of Sea Cucumber (Cucumaria frondosa) Hydrolysate on Fruit Flies and D-Galactose-Induced Aging Mice. J. Funct. Foods 2018, 47, 11–18. [Google Scholar]
- Lin, L.; Zhu, Q.; Zheng, L.; Zhao, M.; Fan, J.; Liu, S. Preparation of Sea Cucumber (Stichopus variegatus) Peptide Fraction with Desired Organoleptic Property and Its Anti-Aging Activity in Fruit Flies and D-Galactose-Induced Aging Mice. J. Funct. Foods 2020, 69, 103954. [Google Scholar]
- Li, J.; Chen, Y.; Yuan, N.; Zeng, M.; Zhao, Y.; Yu, R.; Liu, Z.; Wu, H.; Dong, S. A Novel Natural Influenza A H1N1 Virus Neuraminidase Inhibitory Peptide Derived from Cod Skin Hydrolysates and Its Antiviral Mechanism. Mar. Drugs 2018, 16, 377. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Jayawardena, T.U.; Wu, J. Marine Proteins and Peptides: Production, Biological Activities, and Potential Applications. Food Innov. Adv. 2023, 2, 69–84. [Google Scholar] [CrossRef]
- Nobile, V.; Duclos, E.; Michelotti, A.; Bizzaro, G.; Negro, M.; Soisson, F. Supplementation with a Fish Protein Hydrolysate (Micromesistius poutassou): Effects on Body Weight, Body Composition, and CCK/GLP-1 Secretion. Food Nutr. Res. 2016, 60, 29857. [Google Scholar] [CrossRef]
- Cudennec, B.; Balti, R.; Ravallec, R.; Caron, J.; Bougatef, A.; Dhulster, P.; Nedjar, N. In Vitro Evidence for Gut Hormone Stimulation Release and Dipeptidyl-Peptidase IV Inhibitory Activity of Protein Hydrolysate Obtained from Cuttlefish (Sepia officinalis) Viscera. Food Res. Int. 2015, 78, 238–245. [Google Scholar] [CrossRef]
- Wu, J.Y.; Tso, R.; Yong, Y.N.; Lim, S.P.S.; Wheeler, T.; Nag, A.; Cheng, L.; Talukder, M.M.R.; Huffman, L.; Quek, S.Y.; et al. Effects of the Consumption of Algal Biomass Versus Protein Concentrate on Postprandial Satiety and Metabolism. Future Foods 2024, 10, 100436. [Google Scholar]
- Kim, K.S.; Bae, W.J.; Kim, S.J.; Kang, K.H.; Kim, S.K.; Cho, H.J.; Hong, S.H.; Lee, J.Y.; Kim, S.W. Improvement of Erectile Dysfunction by the Active Peptide from Urechis unicinctus by High Temperature/Pressure and Ultra-Wave Assisted Lysis in Streptozotocin-Induced Diabetic Rats. Int. Braz. J. Urol. 2016, 42, 825–837. [Google Scholar] [CrossRef]
- Xue, C.; Wei, Y. Oyster Peptide Prevents the Occurrence of Exercise-Hypogonadal Male Condition by Improving the Function of Pituitary Gonadal Axis in Male Rats. Andrologia 2021, 53, e14005. [Google Scholar]
- Sudirman, S.; Su, C.Y.; Tsou, D.; Lee, M.C.; Kong, Z.L. Hippocampus Kuda Protein Hydrolysate Improves Male Reproductive Dysfunction in Diabetic Rats. Biomed. Pharmacother. 2021, 140, 111760. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Li, X.Y.; Wang, J.; He, Y.; Chi, C.F.; Wang, B. Antioxidant Peptides from Protein Hydrolysate of Skipjack Tuna Milt: Purification, Identification, and Cytoprotection on H2O2-Damaged Human Umbilical Vein Endothelial Cells. Process Biochem. 2022, 113, 258–269. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhao, Y.Q.; Wang, Y.M.; Yang, X.R.; Chi, C.F.; Wang, B. Gelatins and Antioxidant Peptides from Skipjack Tuna (Katsuwonus pelamis) Skins: Purification, Characterization, and Cytoprotection on Ultraviolet-A Injured Human Skin Fibroblasts. Food Biosci. 2022, 50, 102138. [Google Scholar]
- Phetchthumrongchai, T.; Tachapuripunya, V.; Chintong, S.; Roytrakul, S.; E-Kobon, T.; Klaypradit, W. Properties of Protein Hydrolysates and Bioinformatics Prediction of Peptides Derived from Thermal and Enzymatic Process of Skipjack Tuna (Katsuwonus pelamis) Roe. Fishes 2022, 7, 255. [Google Scholar] [CrossRef]
- Sheng, Y.; Qiu, Y.T.; Wang, Y.M.; Chi, C.F.; Wang, B. Novel Antioxidant Collagen Peptides of Siberian Sturgeon (Acipenser baerii) Cartilages: The Preparation, Characterization, and Cytoprotection of H2O2-Damaged Human Umbilical Vein Endothelial Cells (HUVECs). Mar. Drugs 2022, 20, 325. [Google Scholar]
- Oh, Y.; Ahn, C.B.; Je, J.Y. Cytoprotective Role of Edible Seahorse (Hippocampus abdominalis)-Derived Peptides in H2O2-Induced Oxidative Stress in Human Umbilical Vein Endothelial Cells. Mar. Drugs 2021, 19, 86. [Google Scholar] [CrossRef]
- Ma, Y.; Huang, K.; Wu, Y. In Vivo/In Vitro Properties of Novel Antioxidant Peptide from Pinctada fucata. J. Microbiol. Biotechnol. 2021, 31, 33–42. [Google Scholar]
- Sukkhown, P.; Pirak, T.; Jangchud, K.; Prinyawiwatkul, W. Novel Peptides from Dried Squid Head By-Products Obtained from Snack Process. Int. J. Food Sci. Technol. 2021, 56, 5506–5517. [Google Scholar]
- He, Y.; Pan, X.; Chi, C.F.; Sun, K.L.; Wang, B. Ten New Pentapeptides from Protein Hydrolysate of Miiuy Croaker (Miichthys miiuy) Muscle: Preparation, Identification, and Antioxidant Activity Evaluation. LWT-Food Sci. Technol. 2019, 105, 1–8. [Google Scholar]
- Qian, Z.J.; Zhang, Y.Y.; Oh, G.W.; Heo, S.Y.; Park, W.S.; Choi, I.W.; Jung, W.K. Antioxidant and Angiotensin I Converting Enzyme Inhibition Effects and Antihypertensive Effect in Spontaneously Hypertensive Rats of Peptide Isolated from Boiled Abalone By-Products, Haliotis discus hannai. J. Aquat. Food Prod. Technol. 2018, 27, 946–960. [Google Scholar] [CrossRef]
- Kim, S.S.; Ahn, C.B.; Moon, S.W.; Je, J.Y. Purification and Antioxidant Activities of Peptides from Sea Squirt (Halocynthia roretzi) Protein Hydrolysates Using Pepsin Hydrolysis. Food Biosci. 2018, 25, 128–133. [Google Scholar] [CrossRef]
- Lima, D.A.S.; Santos, M.M.F.; Duvale, R.L.F.; Bezerra, T.K.A.; Araújo, Í.B.D.S.; Madruga, M.S.; da Silva, F.A.P. Technological Properties of Protein Hydrolysate from the Cutting Byproduct of Serra Spanish Mackerel (Scomberomorus brasiliensis). J. Food Sci. Technol. 2021, 58, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Jalili, H.; Ranaei-Siadat, S.O.; Amrane, A. Potential Health Effects of Enzymatic Protein Hydrolysates from Chlorella vulgaris. Appl. Food Biotechnol. 2016, 3, 160–169. [Google Scholar]
- López-Abarrategui, C.; McBeth, C.; Mandal, S.M.; Sun, Z.J.; Heffron, G.; Alba-Menéndez, A.; Migliolo, L.; Reyes-Acosta, O.; García-Villarino, M.; Nolasco, D.O.; et al. Cm-p5: An Antifungal Hydrophilic Peptide Derived from the Coastal Mollusk Cenchritis muricatus (Gastropoda: Littorinidae). FASEB J. 2015, 29, 3315–3325. [Google Scholar] [CrossRef]
- Joshi, I.; Janagaraj, K.; Nazeer, R.A. Isolation and Characterization of Angiotensin I-Converting Enzyme (ACE-I) Inhibition and Antioxidant Peptide from By-Catch Shrimp (Oratosquilla woodmasoni) Waste. Biocatal. Agric. Biotechnol. 2020, 29, 101770. [Google Scholar] [CrossRef]
- Kang, N.; Ko, S.C.; Kim, H.S.; Yang, H.W.; Ahn, G.; Lee, S.C.; Lee, T.G.; Lee, J.S.; Jeon, Y.J. Structural Evidence for Antihypertensive Effect of an Antioxidant Peptide Purified from the Edible Marine Animal Styela clava. J. Med. Food 2020, 23, 132–138. [Google Scholar] [CrossRef]
- Li, M.; Liang, Q.; Zhang, Y.; Jiang, X.; Gu, Y.; Song, X.; Wang, X. Screening of Potential Angiotensin-Converting Enzyme-Inhibitory Peptides in Squid (Todarodes pacificus) Skin Hydrolysates: Preliminary Study of Its Mechanism of Inhibition. Mar. Drugs 2025, 23, 81. [Google Scholar] [CrossRef]
- Fan, Y.; Yu, Z.; Zhao, W.; Ding, L.; Zheng, F.; Li, J.; Liu, J. Identification and Molecular Mechanism of Angiotensin-Converting Enzyme Inhibitory Peptides from Larimichthys crocea Titin. Food Sci. Hum. Wellness 2020, 9, 257–263. [Google Scholar] [CrossRef]
- Purcell, D.; Packer, M.A.; Hayes, M. Angiotensin-I-Converting Enzyme Inhibitory Activity of Protein Hydrolysates Generated from the Macroalga Laminaria digitata (Hudson) JV Lamouroux 1813. Foods 2022, 11, 1792. [Google Scholar]
- Purcell, D.; Packer, M.A.; Hayes, M. Identification of Bioactive Peptides from a Laminaria digitata Protein Hydrolysate Using In Silico and In Vitro Methods to Identify Angiotensin-1-Converting Enzyme (ACE-1) Inhibitory Peptides. Mar. Drugs 2023, 21, 90. [Google Scholar] [CrossRef]
- Ishak, N.H.; Shaik, M.I.; Yellapu, N.K.; Howell, N.K.; Sarbon, N.M. Purification, Characterization, and Molecular Docking Study of Angiotensin-I Converting Enzyme (ACE) Inhibitory Peptide from Shortfin Scad (Decapterus macrosoma) Protein Hydrolysate. J. Food Sci. Technol. 2021, 58, 4567–4577. [Google Scholar] [CrossRef]
- Su, Y.; Chen, S.; Cai, S.; Liu, S.; Pan, N.; Su, J.; Qiao, K.; Xu, M.; Chen, B.; Yang, S.; et al. A Novel Angiotensin-I-Converting Enzyme (ACE) Inhibitory Peptide from Takifugu flavidus. Mar. Drugs 2021, 19, 651. [Google Scholar] [CrossRef]
- Kaewsahnguan, T.; Noitang, S.; Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Choowongkomon, K.; Karnchanatat, A. A Novel Angiotensin I-Converting Enzyme Inhibitory Peptide Derived from the Trypsin Hydrolysates of Salmon Bone Proteins. PLoS ONE 2021, 16, e0256595. [Google Scholar] [CrossRef] [PubMed]
- Naik, A.S.; Whitaker, R.D.; Albrektsen, S.; Solstad, R.G.; Thoresen, L.; Hayes, M. Mesopelagic Fish Protein Hydrolysates and Extracts: A Source of Novel Antihypertensive and Anti-Diabetic Peptides. Front. Mar. Sci. 2021, 8, 719608. [Google Scholar] [CrossRef]
- Kumagai, Y.; Kitade, Y.; Kobayashi, M.; Watanabe, K.; Kurita, H.; Takeda, H.; Yasui, H.; Kishimura, H. Identification of ACE Inhibitory Peptides from Red Alga Mazzaella japonica. Eur. Food Res. Technol. 2020, 246, 2225–2231. [Google Scholar] [CrossRef]
- Sun, S.; Xu, X.; Sun, X.; Zhang, X.; Chen, X.; Xu, N. Preparation and Identification of ACE Inhibitory Peptides from the Marine Macroalga Ulva intestinalis. Mar. Drugs 2019, 17, 179. [Google Scholar] [CrossRef]
- Messina, C.M.; Manuguerra, S.; Arena, R.; Renda, G.; Ficano, G.; Randazzo, M.; Fricano, S.; Sadok, S.; Santulli, A. In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolysates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “from Waste to Profit”. Mar. Drugs 2021, 19, 216. [Google Scholar] [CrossRef]
- Wu, Q.; Cai, Q.-F.; Yoshida, A.; Sun, L.-C.; Liu, Y.-X.; Liu, G.-M.; Su, W.-J.; Cao, M.-J. Purification and characterization of two novel angiotensin I-converting enzyme inhibitory peptides derived from R-phycoerythrin of red algae (Bangia fuscopurpurea). Eur. Food Res. Technol. 2017, 243, 779–789. [Google Scholar] [CrossRef]
- Ko, S.C.; Kang, M.C.; Kang, N.; Kim, H.S.; Lee, S.H.; Ahn, G.; Jung, W.K.; Jeon, Y.J. Effect of Angiotensin I-Converting Enzyme (ACE) Inhibition and Nitric Oxide (NO) Production of 6,6′-Bieckol, a Marine Algal Polyphenol, and Its Anti-Hypertensive Effect in Spontaneously Hypertensive Rats. Process Biochem. 2017, 58, 326–332. [Google Scholar] [CrossRef]
- Mesquita, J.X.; de Brito, T.V.; Fontenelle, T.P.C.; Damasceno, R.O.S.; de Souza, M.H.L.P.; de Souza Lopes, J.L.; Beltramini, L.M.; Barbosa, A.L.d.R.; Freitas, A.L.P. Lectin from Red Algae Amansia multifida Lamouroux: Extraction, Characterization, and Anti-Inflammatory Activity. Int. J. Biol. Macromol. 2021, 170, 532–539. [Google Scholar] [CrossRef]
- Balde, A.; Raghavender, P.; Dasireddy, S.; Abraham, J.; Prasad, S.; Joshi, I.; Abdul, N.R. Crab Pentapeptide and Its Anti-Inflammatory Activity on 16Macrophage Cells. Int. J. Pept. Res. Ther. 2021, 27, 2595–2605. [Google Scholar]
- Yuan, L.; Chu, Q.; Wu, X.; Yang, B.; Zhang, W.; Jin, W.; Gao, R. Anti-Inflammatory and Antioxidant Activity of Peptides from Ethanol-Soluble Hydrolysates of Sturgeon (Acipenser schrenckii) Cartilage. Front. Nutr. 2021, 8, 689648. [Google Scholar]
- Balde, A.; Chatterjee, A.; Shukla, S.; Joshi, I.; Benjakul, S.; Kim, S.K.; Nazeer, R.A. Purification and Identification of Bioactive Oligopeptide from Indian Halibut (Psettodes erumei) Muscle Tissue and Its Inflammation Suppressing Effect In Vitro. Int. J. Pept. Res. Ther. 2023, 29, 80. [Google Scholar]
- Durand, R.; Pellerin, G.; Thibodeau, J.; Fraboulet, E.; Marette, A.; Bazinet, L. Screening for Metabolic Syndrome Application of a Herring By-Product Hydrolysate After Its Separation by Electrodialysis with Ultrafiltration Membrane and Identification of Novel Anti-Inflammatory Peptides. Sep. Purif. Technol. 2020, 235, 116205. [Google Scholar] [CrossRef]
- Lugo, J.M.; Tafalla, C.; Oliva, A.; Pons, T.; Oliva, B.; Aquilino, C.; Morales, R.; Estrada, M.P. Evidence for Antimicrobial and Anticancer Activity of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) from North African Catfish (Clarias gariepinus): Its Potential Use as Novel Therapeutic Agent in Fish and Humans. Fish Shellfish Immunol. 2019, 86, 559–570. [Google Scholar]
- Huang, F.; Ding, G.; Yang, Z.; Yu, F. Two Novel Peptides Derived from Sinonovacula constricta Inhibit the Proliferation and Induce Apoptosis of Human Prostate Cancer Cells. Mol. Med. Rep. 2017, 16, 6697–6707. [Google Scholar]
- Wang, Z.; Zhang, X. Isolation and Identification of Anti-Proliferative Peptides from Spirulina platensis Using Three-Step Hydrolysis. J. Sci. Food Agric. 2017, 97, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with α-Amylase Inhibitory Potential from Enzymatic Protein Hydrolysates of Red Seaweed (Porphyra spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar]
- Feng, L.; Tu, M.; Qiao, M.; Fan, F.; Chen, H.; Song, W.; Du, M. Thrombin Inhibitory Peptides Derived from Mytilus edulis Proteins: Identification, Molecular Docking, and In Silico Prediction of Toxicity. Eur. Food Res. Technol. 2018, 244, 207–217. [Google Scholar]
- Fan, X.; Cui, Y.; Zhang, R.; Zhang, X. Purification and Identification of Antiobesity Peptides Derived from Spirulina platensis. J. Funct. Foods 2018, 47, 350–360. [Google Scholar]
- Zhao, Y.; Lu, K.; Piao, X.; Song, Y.; Wang, L.; Zhou, R.; Gao, P.; Khong, H.Y. Collagens for Surimi Gel Fortification: Type-Dependent Effects and the Difference Between Type I and Type II. Food Chem. 2023, 407, 135157. [Google Scholar] [PubMed]
- Sousa, S.C.; Fragoso, S.P.; Penna, C.R.A.; Arcanjo, N.M.O.; Silva, F.A.P.; Ferreira, V.C.S.; Barreto, M.D.S.; Araújo, Í.B.S. Quality Parameters of Frankfurter-Type Sausages with Partial Replacement of Fat by Hydrolyzed Collagen. LWT-Food Sci. Technol. 2017, 76, 320–325. [Google Scholar]
- Dey, P.; Kadharbasha, S.; Bajaj, M.; Das, J.; Chakraborty, T.; Bhat, C.; Banerjee, P. Contribution of Quasifibrillar Properties of Collagen Hydrolysates Towards Lowering of Interface Tension in Emulsion-Based Food Leading to Shelf-Life Enhancement. Food Bioprocess Technol. 2021, 14, 1566–1586. [Google Scholar]
- Shori, A.B.; Tin, Y.P.; Baba, A.S. Codonopsis Pilosula and Fish Collagen Yogurt: Proteolytic, Potential Angiotensin-I Converting Enzyme (ACE) Inhibitory Activity and Sensory Properties. LWT Food Sci. Technol. 2022, 165, 113729. [Google Scholar]
- Bhagwat, P.K.; Dandge, P.B. Isolation, Characterization and Valorizable Applications of Fish Scale Collagen in Food and Agriculture Industries. Biocatal. Agric. Biotechnol. 2016, 7, 234–240. [Google Scholar]
- Sae-Leaw, T.; Aluko, R.E.; Chantakun, K.; Benjakul, S. Physicochemical, Antioxidant and Sensory Properties of Ready-to-Drink Chrysanthemum Tea Fortified with Hydrolyzed Collagen from Salmon Scale Ossein. J. Aquat. Food Prod. Technol. 2021, 30, 1159–1172. [Google Scholar]
- Bhuimbar, M.V.; Bhagwat, P.K.; Dandge, P.B. Extraction and Characterization of Acid Soluble Collagen from Fish Waste: Development of Collagen-Chitosan Blend as Food Packaging Film. J. Environ. Chem. Eng. 2019, 7, 102983. [Google Scholar]
- Song, Z.; Liu, H.; Huang, A.; Zhou, C.; Hong, P.; Deng, C. Collagen/Zein Electrospun Films Incorporated with Gallic Acid for Tilapia (Oreochromis niloticus) Muscle Preservation. J. Food Eng. 2022, 317, 110860. [Google Scholar]
- Ben, S.E.; Sadok, S. Collagen from Cartilaginous Fish By-Products for a Potential Application in Bioactive Film Composite. Mar. Drugs 2018, 16, 211. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Gobe, G.; Masci, P.; Osborne, S.A. Marine Bioactive Compounds and Health Promoting Perspectives; Innovation Pathways for Drug Discovery. Trends Food Sci. Technol. 2016, 50, 44–55. [Google Scholar]
- Nurdiani, R.; Prihanto, A.A.; Firdaus, M. Seafood as Source of Protein-Based Functional Foods. In Encyclopedia of Marine Biotechnology; Kim, S.K., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 2987–2997. [Google Scholar]
- Raja, K.; Kadirvel, V.; Subramaniyan, T. Seaweeds, an Aquatic Plant-Based Protein for Sustainable Nutrition—A Review. Future Foods 2022, 5, 100142. [Google Scholar] [CrossRef]
- Bălașa, A.F.; Chircov, C.; Grumezescu, A.M. Marine Biocompounds for Neuroprotection—A Review. Mar. Drugs 2020, 18, 290. [Google Scholar] [CrossRef] [PubMed]
- Al-Nimry, S.; Dayah, A.A.; Hasan, I.; Daghmash, R. Cosmetic, Biomedical and Pharmaceutical Applications of Fish Gelatin/Hydrolysates. Mar. Drugs 2021, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Mathew-Steiner, S.S.; Roy, S.; Sen, C.K. Collagen in Wound Healing. Bioengineering 2021, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Sable, R.; Parajuli, P.; Jois, S. Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Mar. Drugs 2017, 15, 124. [Google Scholar] [CrossRef]
- Cappello, E.; Nieri, P. From Life in the Sea to the Clinic: The Marine Drugs Approved and under Clinical Trial. Life 2021, 11, 1390. [Google Scholar] [CrossRef]
- Biondini, M.; Kiepas, A.; El-Houjeiri, L.; Annis, M.G.; Hsu, B.E.; Fortier, A.M.; Morin, G.; Martina, J.A.; Sirois, I.; Aguilar-Mahecha, A.; et al. HSP90 Inhibitors Induce GPNMB Cell-Surface Expression by Modulating Lysosomal Positioning and Sensitize Breast Cancer Cells to Glembatumumab Vedotin. Oncogene 2022, 41, 1701–1717. [Google Scholar]
- White, K.M.; Rosales, R.; Yildiz, S.; Kehrer, T.; Miorin, L.; Moreno, E.; Jangra, S.; Uccellini, M.B.; Rathnasinghe, R.; Coughlan, L.; et al. Plitidepsin Has Potent Preclinical Efficacy Against SARS-CoV-2 by Targeting the Host Protein eEF1A. Science 2021, 371, 926–931. [Google Scholar]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed Marine Natural Products in the Pharmaceutical and Cosmeceutical Industries: Tips for Success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef]
- van Andel, L.; Fudio, S.; Rosing, H.; Munt, S.; Miguel-Lillo, B.; González, I.; Tibben, M.M.; de Vries, N.; de Vries Schultink, A.H.M.; Schellens, J.H.M.; et al. Pharmacokinetics and Excretion of 14C-Plitidepsin in Patients with Advanced Cancer. Investig. New Drugs 2017, 35, 589–598. [Google Scholar]
- Alves, A.L.; Marques, A.L.; Martins, E.; Silva, T.H.; Reis, R.L. Cosmetic Potential of Marine Fish Skin Collagen. Cosmetics 2017, 4, 39. [Google Scholar] [CrossRef]
- Aguirre-Cruz, G.; León-López, A.; Cruz-Gómez, V.; Jiménez-Alvarado, R.; Aguirre-Álvarez, G. Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants 2020, 9, 181. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H. Protective Effect of Gelatin Polypeptides from Pacific Cod (Gadus macrocephalus) Against UV Irradiation-Induced Damages by Inhibiting Inflammation and Improving Transforming Growth Factor-β/Smad Signaling Pathway. J. Photochem. Photobiol. B Biol. 2016, 162, 633–640. [Google Scholar]
- Zhang, D.; Wu, X.; Chen, J.; Lin, K. The Development of Collagen-Based Composite Scaffolds for Bone Regeneration. Bioact. Mater. 2018, 3, 129–138. [Google Scholar]
- Romanenko, N.Y.; Mezenova, O.Y.; Nekrasova, Y.O. Specialized Sports Nutrition Products Using Protein Hydrolysis Compositions of Collagen-Containing Fish Raw Materials. Vestnik MGTU 2021, 24, 414–427. [Google Scholar]
- Restaino, O.F.; Finamore, R.; Stellavato, A.; Diana, P.; Bedini, E.; Trifuoggi, M.; De Rosa, M.; Schiraldi, C. European Chondroitin Sulfate and Glucosamine Food Supplements: A Systematic Quality and Quantity Assessment Compared to Pharmaceuticals. Carbohydr. Polym. 2019, 222, 114984. [Google Scholar] [CrossRef]

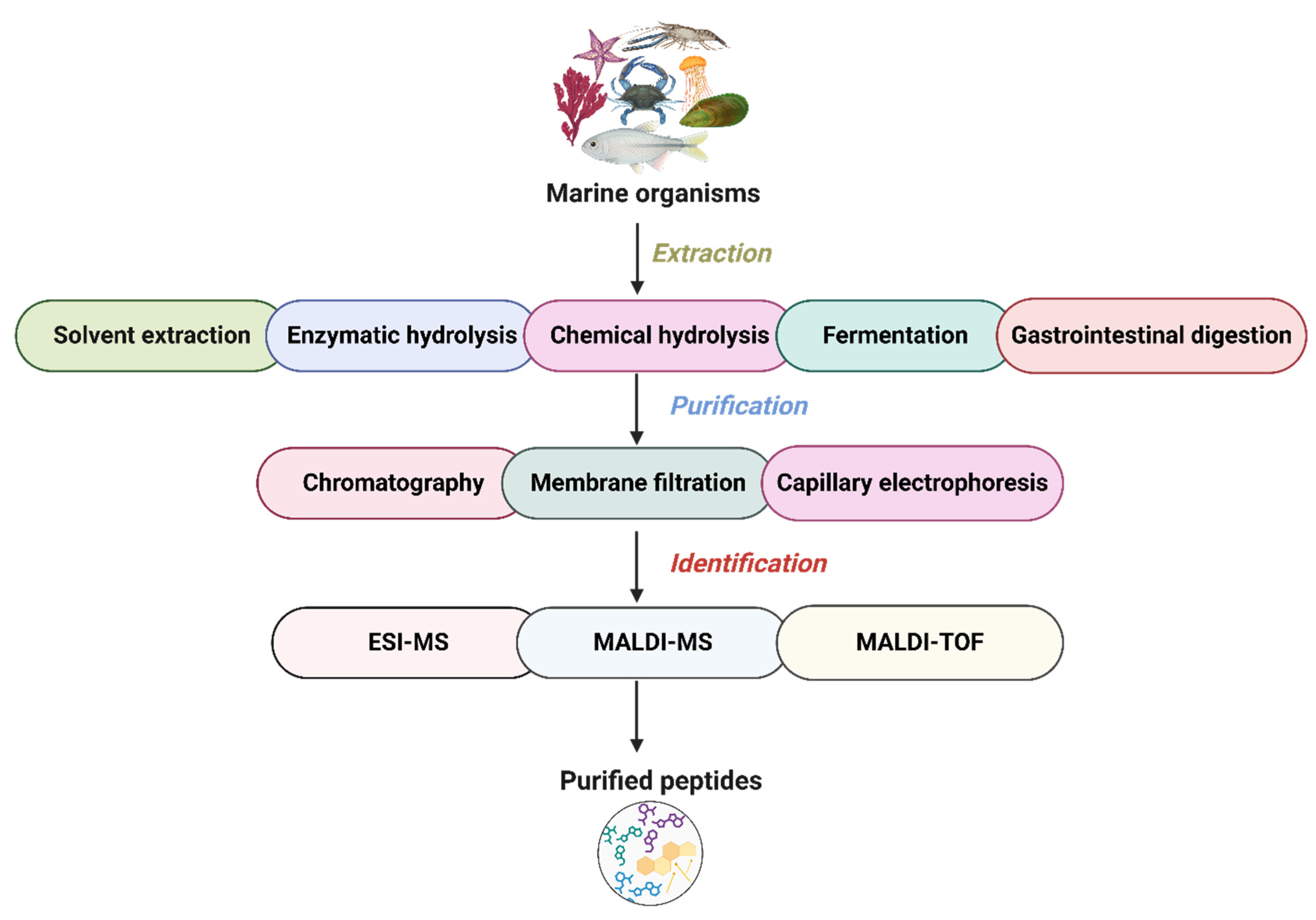

| Marine Source | Compound/Drug | Product Type | Application | Status/Clinical Trial | References |
|---|---|---|---|---|---|
| Aplidium albicans | Plitidepsin(Aplidin®) | Natural product | Antiviral(SARS-CoV-2) | Phase III | [280,283] |
| Plitidepsin (Aplidin®) | Natural product | Advanced cancer | Phase III | [284] | |
| Dolabella auricularia | Brentuximab vedotin | Derivative | Cancer treatment | FDA approved | [279] |
| Conus magus | Ziconotide | Natural product | Analgesic | FDA approved | [279] |
| Bonito (Katsuobushi) | Katsuobushioligopeptide | Natural product | Antihypertensive | Sold as nutraceutical | [2] |
| Fish hydrolysate | Nutripeptin®/Hydro MN Peptide® | Natural product | Postprandial blood glucose control | Sold as nutraceutical | [2] |
| Chlorella vulgaris | Dermochlorella® | Natural product | Skin toner andfirmer | Sold as skin care product | [283] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahidi, F.; Saeid, A. Bioactivity of Marine-Derived Peptides and Proteins: A Review. Mar. Drugs 2025, 23, 157. https://doi.org/10.3390/md23040157
Shahidi F, Saeid A. Bioactivity of Marine-Derived Peptides and Proteins: A Review. Marine Drugs. 2025; 23(4):157. https://doi.org/10.3390/md23040157
Chicago/Turabian StyleShahidi, Fereidoon, and Abu Saeid. 2025. "Bioactivity of Marine-Derived Peptides and Proteins: A Review" Marine Drugs 23, no. 4: 157. https://doi.org/10.3390/md23040157
APA StyleShahidi, F., & Saeid, A. (2025). Bioactivity of Marine-Derived Peptides and Proteins: A Review. Marine Drugs, 23(4), 157. https://doi.org/10.3390/md23040157