

Enhancing CO2 Fixation in Microalgal Systems: Mechanistic Insights and Bioreactor Strategies

Abstract
1. Introduction
2. Methodologies and Devices for Enhancing CO2 Mass Transfer in Microalgal Systems
2.1. CO2 Mass-Transfer Process
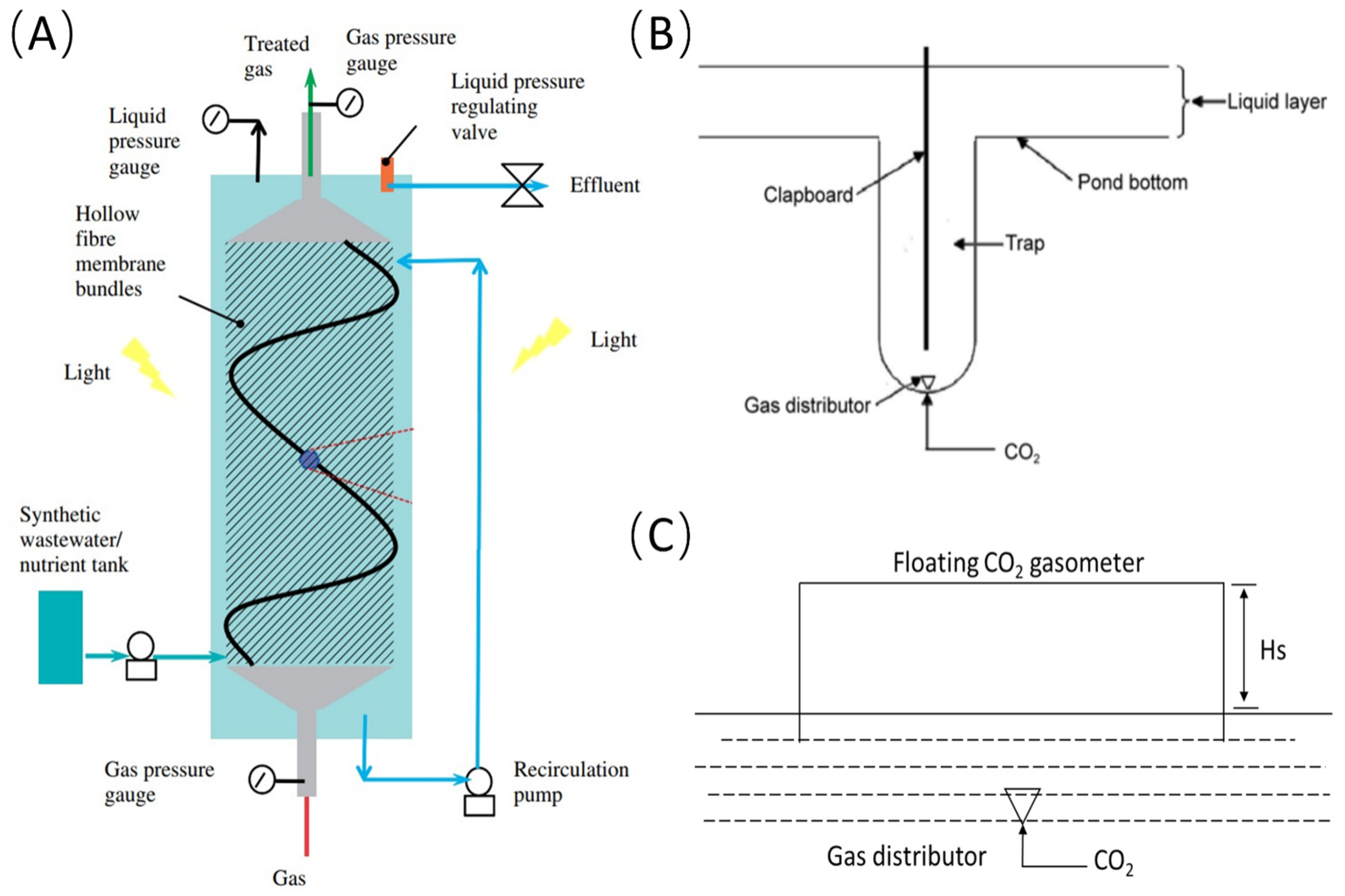
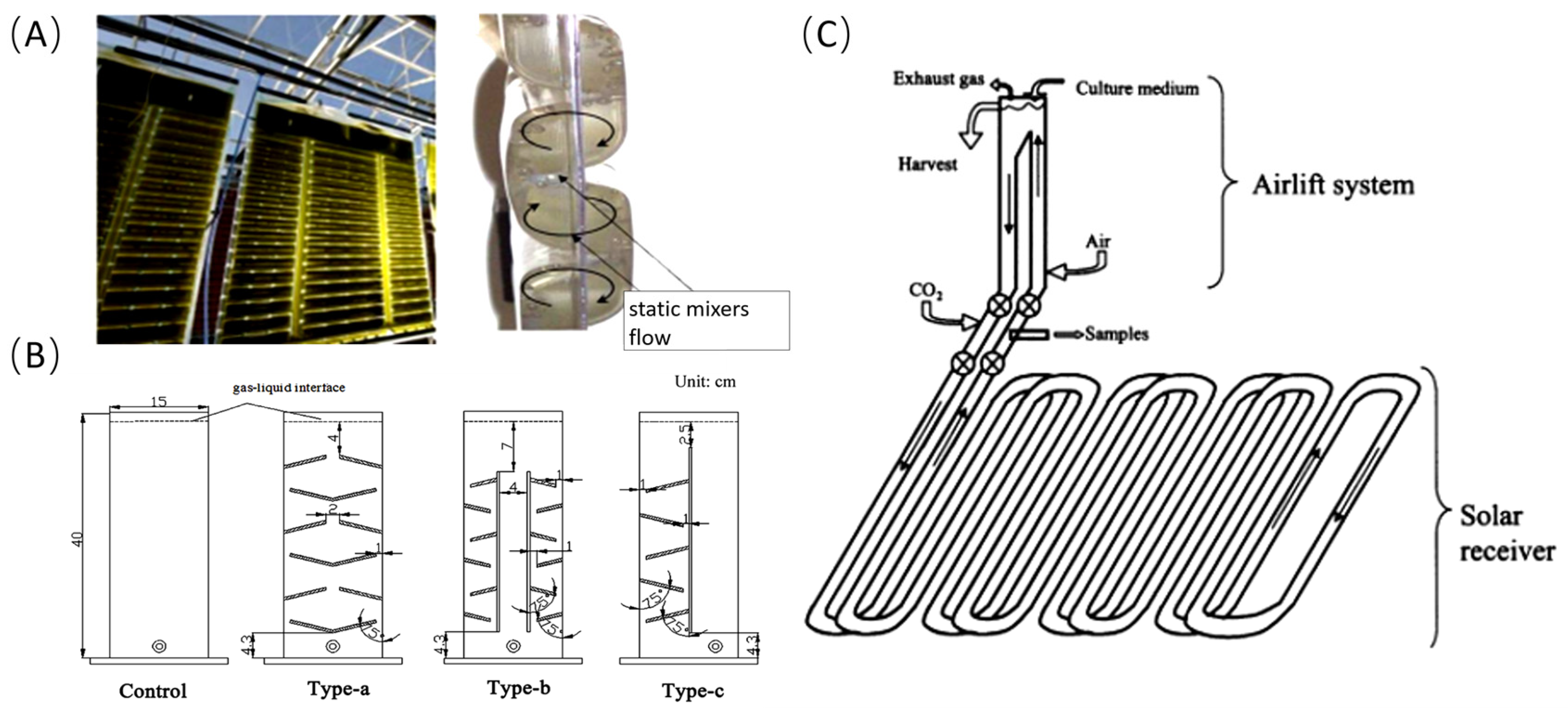
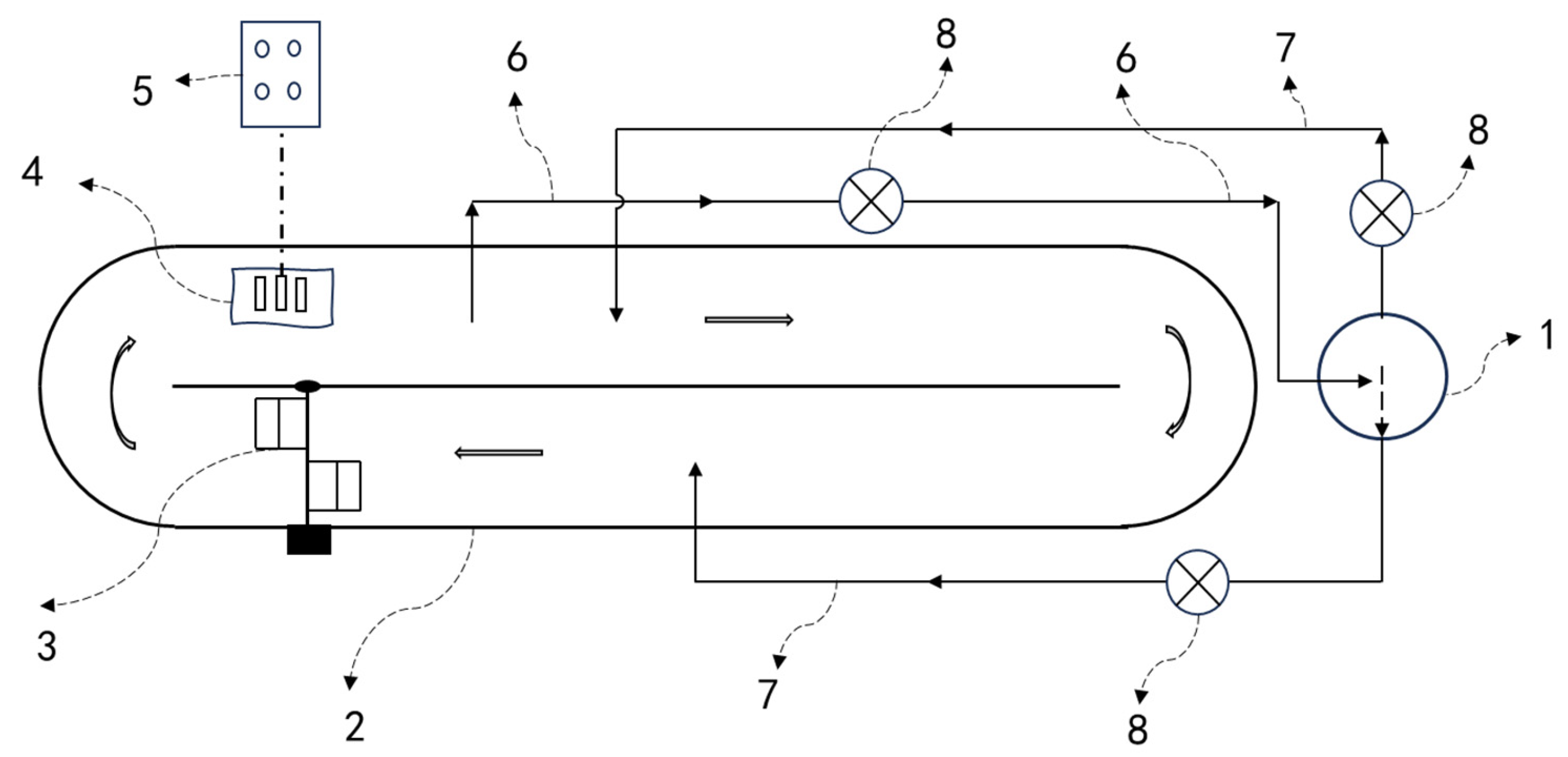
2.2. In Situ CO2 Supplementation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgal Culture System | Methodologies or Devices | Species | Biomass | CO2 Utilization Efficiency | Mechanisms | Reference |
|---|---|---|---|---|---|---|
| Photobioreactor | Hollow fiber membrane | Spirulina platensis | 2131 mg/ L ↑ | 85% ↑ | Increase the interfacial contact area available for gas transfer | [19,20] |
| Raceway pond | An ascending channel | Scenedesmus sp. | 0.16 ± 0.03 g/(L·d) ↑ | 50% ↑ | Increase mixing intensity | [21] |
| Raceway pond | CO2 supplementation trap device | Spirulina platensis | 3.45–6.04 g/(m2·d) ↑ | 90% ↑ | Prolong gas–liquid contact time | [22] |
| Open pond | Leak-proof cover | Cyanobacterium sp. | 2.5 g/L ↑ | 80% ↑ | Create a large gas–liquid exchange area | [23] |
| Open pond | Submerged cover-type | Spirulina platensis | 13.3 g/(m2·d) ↑ | 92% ↑ | Prolong gas–liquid contact time | [24] |
| Photobioreactor | Multiple chambers | Nannochloropsis salina | 0.12 g/(L·d) ↑ | 80% ↑ | Enhance bubble residence time | [25] |
| Flat-plate PBRs | Inclined baffles | Chlorella pyrenoidosa. | 1.3 g/ L | No data | Increase mixing intensity | [26] |
| Raceway pond | Vertical absorption tower | Chlorella pyrenoidosa. | 20 g/(m2·d) ↑ | 83% ↑ | Prolong gas–liquid contact time | [28] |
| Open pond | Absorption tank | Spirulina platensis | 6–12 g/(m2·d) | >50% | Increase mixing intensity | [4,29] |
2.3. Ex Situ CO2 Supplementation
3. Strategies for Enhancing CO2 Mass Transfer Using Chemical Solvents
3.1. Novel Enhancing Mechanism of Introducing Chemical Reaction
3.2. Novel Enhancing Mechanism of Altering the Medium’s Physical Properties
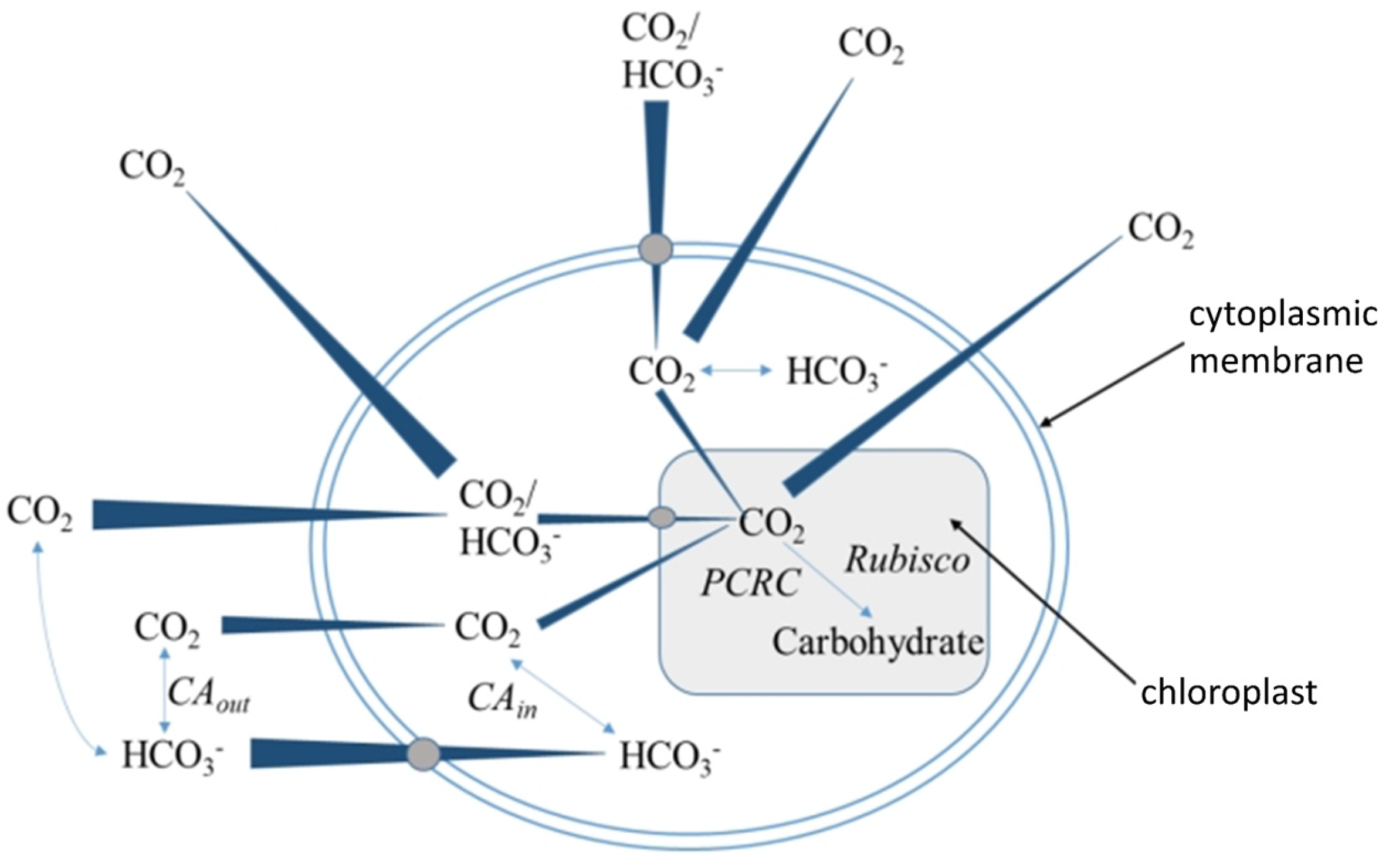
4. Carbonic Anhydrase-Assisted CO2 Absorption and Conversion
5. Challenges and Future Prospects
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, D.Y.Y.; Khoo, K.S.; Chew, K.W.; Tao, Y.; Ho, S.-H.; Show, P.L. Potential utilization of bioproducts from microalgae for the quality enhancement of natural products. Bioresour. Technol. 2020, 304, 122997. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Ağbulut, Ü.; Sirohi, R.; Lichtfouse, E.; Chen, W.-H.; Len, C.; Show, P.L.; Le, A.T.; Nguyen, X.P.; Hoang, A.T. Microalgae bio-oil production by pyrolysis and hydrothermal liquefaction: Mechanism and characteristics. Bioresour. Technol. 2023, 376, 128860. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Hong, Y.; He, Y.T.; Liu, Y. Growth and high-valued products accumulation characteristics of microalgae in saline-alkali leachate from Inner Mongolia. Environ. Sci. Pollut. Res. 2019, 26, 36985–36992. [Google Scholar] [CrossRef]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a future food source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Agbebi, T.V.; Ojo, E.O.; Watson, I.A. Towards optimal inorganic carbon delivery to microalgae culture. Algal Res. 2022, 67, 102841. [Google Scholar] [CrossRef]
- Su, Z.F.; Kang, R.J.; Shi, S.Y.; Cong, W.; Cai, Z.L. An effective device for gas-liquid oxygen removal in enclosed microalgae culture. Appl. Biochem. Biotechnol. 2010, 160, 428–437. [Google Scholar] [CrossRef]
- Su, Z.F.; Kang, R.J.; Shi, S.Y.; Cong, W.; Cai, Z.L. Study on the destabilization mixing in the flat plate photobioreactor by means of CFD. Biomass Bioenergy 2010, 34, 1879–1884. [Google Scholar] [CrossRef]
- Vasumathi, K.K.; Premalatha, M.; Subramanian, P. Parameters influencing the design of photobioreactor for the growth of microalgae. Renew. Sustain. Energy Rev. 2012, 16, 5443–5450. [Google Scholar] [CrossRef]
- Demirbas, A. Production economics of high-quality microalgae. Energy Sources Part B Econ. Plan. Policy 2017, 12, 395–401. [Google Scholar] [CrossRef]
- Soares, F.R.; Martins, G.; Seo, E.S.M. An assessment of the economic aspects of CO2 sequestration in a route for biodiesel production from microalgae. Environ. Technol. 2013, 34, 1777–1781. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.E.; Capasso, C.; Marzocchella, A.; Salatino, P. Immobilization of carbonic anhydrase for CO2 capture and utilization. Appl. Microbiol. Biotechnol. 2022, 106, 3419–3430. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kentish, S.E.; Martin, G.J.O. Direct air capture of CO2 by microalgae with buoyant beads encapsulating carbonic anhydrase. ACS Sustain. Chem. Eng. 2021, 9, 9698–9706. [Google Scholar] [CrossRef]
- Valdés, F.J.; Hernández, M.R.; Catalá, L.; Marcilla, A. Estimation of CO2 stripping/CO2 microalgae consumption ratios in a bubble column photobioreactor using the analysis of the pH profiles. Application to Nannochloropsis oculata microalgae culture. Bioresour. Technol. 2012, 119, 1–6. [Google Scholar] [CrossRef]
- Ughetti, A.; Roncaglia, F.; Anderlini, B.; D’Eusanio, V.; Russo, A.L.; Forti, L. Integrated Carbonate-Based CO2 Capture-Biofixation through Cyanobacteria. Appl. Sci. 2023, 13, 10779. [Google Scholar] [CrossRef]
- Fu, J.W.; Huang, Y.; Liao, Q.; Xia, A.; Fu, Q.; Zhu, X. Photo-bioreactor design for microalgae: A review from the aspect of CO2 transfer and conversion. Bioresour. Technol. 2019, 292, 121947. [Google Scholar] [CrossRef]
- Ndiaye, M.; Gadoin, E.; Gentric, C. CO2 gas-liquid mass transfer and K_L,a estimation: Numerical investigation in the context of airlift photobioreactor scale-up. Chem. Eng. Res. Des. 2018, 133, 90–102. [Google Scholar] [CrossRef]
- Thobie, C.; Gadoin, E.; Blel, W.; Pruvost, J.; Gentric, C. Global characterization of hydrodynamics and gas-liquid mass transfer in a thin-gap bubble column intended for microalgae cultivation. Chem. Eng. Process. 2017, 122, 76–89. [Google Scholar] [CrossRef]
- Kumar, A.; Yuan, X.; Sahu, A.K.; Dewulf, J.; Ergas, S.J.; Van Langenhove, H. A hollow fiber membrane photo-bioreactor for CO2 sequestration from combustion gas coupled with wastewater treatment: A process engineering approach. J. Chem. Technol. Biotechnol. 2010, 85, 387–394. [Google Scholar] [CrossRef]
- Ferreira, B.S.; Fernandes, H.L.; Reis, A.; Mateus, M. Microporous hollow fibres for carbon dioxide absorption: Mass transfer model fitting and the supplying of carbon dioxide to microalgal cultures. J. Chem. Technol. Biotechnol. 1998, 71, 61–70. [Google Scholar] [CrossRef]
- Ketheesan, B.; Nirmalakhandan, N. Development of a new airlift-driven raceway reactor for algal cultivation. Appl. Energy 2011, 88, 3370–3376. [Google Scholar] [CrossRef]
- Bao, Y.L.; Liu, M.; Wu, X.; Cong, W.; Ning, Z.X. In situ carbon supplementation in large-scale cultivations of Spirulina platensis in open raceway pond. Biotechnol. Bioprocess Eng. 2012, 17, 93–99. [Google Scholar] [CrossRef]
- Chen, Z.S.; Li, T.; Yang, B.J.; Jin, X.J.; Wu, H.L.; Wu, J.Y.; Lu, Y.D.; Xiang, W.Z. Isolation of a novel strain of Cyanobacterium sp. with good adaptation to extreme alkalinity and high polysaccharide yield. J. Oceanol. Limnol. 2021, 39, 1131–1142. [Google Scholar] [CrossRef]
- Cong, W. Horizontal Submerged Hood Carbon Supplementation Device for Open Pond Microalgae Cultivation and Its Carbon Supplementation Method. CN201210138845.8, 30 April 2014. [Google Scholar]
- Bergmann, P.; Ripplinger, P.; Beyer, L.; Trösch, W. Disposable Flat Panel Airlift Photobioreactors. Chem. Ing. Tech. 2013, 85, 202–205. [Google Scholar]
- Huang, J.; Li, Y.; Wan, M.; Yan, Y.; Feng, F.; Qu, X.; Wang, J.; Shen, G.; Li, W.; Fan, J.; et al. Novel flat-plate photobioreactors for microalgae cultivation with special mixers to promote mixing along the light gradient. Bioresour. Technol. 2014, 159, 8–16. [Google Scholar] [CrossRef]
- Razzak, S.A.; Al-Aslani, I.; Hossain, M.M. Hydrodynamics and mass transfer of CO2 in water in a tubular photobioreactor. Eng. Life Sci. 2016, 16, 355–363. [Google Scholar] [CrossRef]
- Putt, R.; Singh, M.; Chinnasamy, S.; Das, K.C. An efficient system for carbonation of high-rate algae pond water to enhance CO2 mass transfer. Bioresour. Technol. 2011, 102, 3240–3245. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal industry in China: Challenges and prospects. J. Appl. Phycol. 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Yan, C.H.; Wang, Z.; Wu, X.; Wen, S.M.; Yu, J.; Cong, W. Outdoor cultivation of Chlorella sp. in an improved thin-film flat-plate photobioreactor in desertification areas. J. Biosci. Bioeng. 2020, 129, 619–623. [Google Scholar] [CrossRef]
- Gao, F.Z.; Ge, B.S.; Xiang, W.Z.; Qin, S. Development of microalgal industries in the past 60 years due to biotechnological research in China: A review. Sci. Sin. Vitae 2021, 51, 26–39. [Google Scholar]
- Basheer, C.; Kamran, M.; Ashraf, M.; Lee, H.K. Enhancing liquid-phase microextraction efficiency through chemical reactions. TrAC Trends Anal. Chem. 2019, 118, 426–433. [Google Scholar] [CrossRef]
- Morales, D.O.; Regalado-Méndez, A.; Pérez-Alonso, C.; Natividad, R. Physical and reactive absorption of CO2 in capillaries: Mass transfer, modelling, and produced chemical species. Chem. Eng. Res. Des. 2023, 198, 247–258. [Google Scholar] [CrossRef]
- Zhu, C.B.; Chen, S.L.; Ji, Y.; Schwaneberg, U.; Chi, Z.Y. Progress toward a bicarbonate-based microalgae production system. Trends Biotechnol. 2022, 40, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.B.; Zhang, R.L.; Cheng, L.Y.; Chi, Z.Y. A recycling culture of Neochloris oleoabundans in a bicarbonate-based integrated carbon capture and algae production system with harvesting by auto-flocculation. Biotechnol. Biofuels 2018, 11, 204. [Google Scholar] [CrossRef]
- Zhu, C.B.; Zhu, H.; Cheng, L.Y.; Chi, Z.Y. Bicarbonate-based carbon capture and algal production system on ocean with floating inflatable-membrane photobioreactor. J. Appl. Phycol. 2018, 30, 875–885. [Google Scholar] [CrossRef]
- Yang, J.; Xu, M.; Zhang, X.Z.; Hu, Q.A.; Sommerfeld, M.; Chen, Y.S. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef]
- Guieysse, B.; Béchet, Q.; Shilton, A. Variability and uncertainty in water demand and water footprint assessments of fresh algae cultivation based on case studies from five climatic regions. Bioresour. Technol. 2013, 128, 317–323. [Google Scholar] [CrossRef]
- Sun, Z.L.; Sun, L.Q.; Liu, Y.H. The potential impact of replacing nitrate with ammonium hydroxide in microalgae production on the biomass productivity and CO2 utilization efficiency. Algal Res. 2022, 67, 102870. [Google Scholar] [CrossRef]
- Bao, Y.L.; Wen, S.M.; Cong, W.; Wu, X.; Ning, Z.X. An Optical-Density-Based Feedback Feeding Method for Ammonium Concentration Control in Spirulina platensis Cultivation. J. Microbiol. Biotechnol. 2012, 22, 967–974. [Google Scholar] [CrossRef]
- Geider, R.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef]
- Ho, T.-Y.; Quigg, A.; Finkel, Z.V.; Milligan, A.J.; Wyman, K.; Falkowski, P.G.; Morel, F.M.M. The elemental composition of some marine phytoplankton. J. Phycol. 2003, 39, 1145–1159. [Google Scholar] [CrossRef]
- Dziejarski, B.; Krzyżyńska, R.; Andersson, K. Current status of carbon capture, utilization, and storage technologies in the global economy: A survey of technical assessment. Fuel 2023, 342, 127776. [Google Scholar] [CrossRef]
- Sun, Z.L.; Xue, S.Z.; Yan, C.H.; Cong, W.; Kong, D.Z. Utilisation of tris(hydroxymethyl)aminomethane as a gas carrier in microalgal cultivation to enhance CO2 utilisation and biomass production. RSC Adv. 2016, 6, 2703–2711. [Google Scholar] [CrossRef]
- Sun, Z.L.; Zhang, D.M.; Yan, C.H.; Cong, W.; Lu, Y.M. Promotion of microalgal biomass production and efficient use of CO2 from flue gas by monoethanolamine. J. Chem. Technol. Biotechnol. 2015, 90, 730–738. [Google Scholar] [CrossRef]
- Cardias, B.B.; Morais, M.G.d.; Costa, J.A.V. CO2 conversion by the integration of biological and chemical methods: Spirulina sp. LEB 18 cultivation with diethanolamine and potassium carbonate addition. Bioresour. Technol. 2018, 267, 77–83. [Google Scholar] [CrossRef]
- Yin, Q.; Mao, W.; Chen, D.; Song, C. Effect of adding tertiary amine TMEDA and space hindered amine DACH on the CO2 chemical absorption-microalgae conversion system. Energy 2023, 263, 125726. [Google Scholar] [CrossRef]
- Kim, G.; Choi, W.; Lee, C.-H.; Lee, K. Enhancement of dissolved inorganic carbon and carbon fixation by green alga Scenedesmus sp. in the presence of alkanolamine CO2 absorbents. Biochem. Eng. J. 2013, 78, 18–23. [Google Scholar] [CrossRef]
- Rosa, G.M.d.; Morais, M.G.d.; Costa, J.A.V. Fed-batch cultivation with CO2 and monoethanolamine: Influence on Chlorella fusca LEB 111 cultivation, carbon biofixation, and biomolecules production. Bioresour. Technol. 2019, 273, 627–633. [Google Scholar] [CrossRef]
- Bai, L.; Lu, S.J.; Qiu, S.; Li, J.; Chen, S.M. Single-step integrated technology for enhanced CO2 biofixation and efficient lipid extraction in microalgal system including a water-immiscible solvent. Chem. Eng. J. 2022, 432, 134374. [Google Scholar] [CrossRef]
- Rosa, G.M.d.; Moraes, L.; de Souza, M.d.R.A.Z.; Costa, J.A.V. Spirulina cultivation with a CO2 absorbent: Influence on growth parameters and macromolecule production. Bioresour. Technol. 2016, 200, 528–534. [Google Scholar] [CrossRef]
- Rosa, G.M.d.; Morais, M.G.d.; Costa, J.A.V. Green alga cultivation with monoethanolamine: Evaluation of CO2 fixation and macromolecule production. Bioresour. Technol. 2018, 261, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Gao, S.; Ma, Q.; Wei, Y.; Li, G. Techno-economic analysis of carbon capture and utilization technologies and implications for China. Renew. Sustain. Energy Rev. 2024, 199, 114550. [Google Scholar] [CrossRef]
- Madejski, P.; Chmiel, K.; Subramanian, N.; Kus, T. Methods and Techniques for CO2 Capture: Review of Potential Solutions and Applications in Modern Energy Technologies. Energies 2022, 15, 887. [Google Scholar] [CrossRef]
- Sun, Z.L.; Xin, M.R.; Li, P.; Sun, L.Q.; Wang, S.K. Enhancing CO2 utilization by a physical absorption-based technique in microalgae culture. Bioprocess Biosyst. Eng. 2021, 44, 1901–1912. [Google Scholar] [CrossRef]
- Li, J.; Zhao, X.; Chang, J.-S.; Miao, X. A Two-Stage Culture Strategy for Scenedesmus sp. FSP3 for CO2 Fixation and the Simultaneous Production of Lutein under Light and Salt Stress. Molecules 2022, 27, 7497. [Google Scholar] [CrossRef]
- Wang, Y.J.; Stessman, D.J.; Spalding, M.H. The CO2 concentrating mechanism and photosynthetic carbon assimilation in limiting CO2: How Chlamydomonas works against the gradient. Plant J. 2015, 82, 429–448. [Google Scholar] [CrossRef]
- Raven, J.A.; Beardall, J.; Giordano, M. Energy costs of carbon dioxide concentrating mechanisms in aquatic organisms. Photosynth. Res. 2014, 121, 111–124. [Google Scholar] [CrossRef]
- Yao, D.; Wu, L.; Tan, D.; Yu, Y.; Jiang, Q.; Wu, H.; Wang, Y.; Liu, Y. Enhancing CO2 fixation by microalgae in a Photobioreactor: Molecular mechanisms with exogenous carbonic anhydrase. Bioresour. Technol. 2024, 408, 131176. [Google Scholar] [CrossRef]
- Swarnalatha, G.V.; Hegde, N.S.; Chauhan, V.S.; Sarada, R. The effect of carbon dioxide rich environment on carbonic anhydrase activity, growth and metabolite production in indigenous freshwater microalgae. Algal Res. 2015, 9, 151–159. [Google Scholar] [CrossRef]
- van Hille, R.; Fagan, M.; Bromfield, L.; Pott, R. A modified pH drift assay for inorganic carbon accumulation and external carbonic anhydrase activity in microalgae. J. Appl. Phycol. 2014, 26, 377–385. [Google Scholar] [CrossRef]
- González, J.M.; Fisher, S.Z. Carbonic Anhydrases in Industrial Applications. In Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications; Frost, S.C., McKenna, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 405–426. [Google Scholar]
- Lin, J.Y.; Sri Wahyu Effendi, S.; Ng, I.S. Enhanced carbon capture and utilization (CCU) using heterologous carbonic anhydrase in Chlamydomonas reinhardtii for lutein and lipid production. Bioresour. Technol. 2022, 351, 127009. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.H.; Yang, J.; Jeon, H.; Kim, H.S.; Pack, S.P.; Jin, E.; Kim, J. Stabilized and immobilized carbonic anhydrase on electrospun nanofibers for enzymatic CO2 conversion and utilization in expedited microalgal growth. Environ. Sci. Technol. 2020, 54, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.W.; Li, M.J.; Hung, T.C. Enhancing CO2 dissolution and inorganic carbon conversion by metal–organic frameworks improves microalgal growth and carbon fixation efficiency. Bioresour. Technol. 2024, 407, 131113. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.R. Study on the Effect of Exogenous Carbonic Anhydrase in Promoting Carbon Absorption in Microalgae Cells. Master’s Thesis, Yantai University, Yantai, China, 2022. [Google Scholar] [CrossRef]
- Wang, X.; Li, M.; Liu, Z.; Shi, Z.; Yu, D.; Ge, B.; Huang, F. Carbonic anhydrase encapsulation using bamboo cellulose scaffolds for efficient CO2 capture and conversion. Int. J. Biol. Macromol. 2024, 277, 134410. [Google Scholar] [CrossRef]
| Absorbents or Immobilized Enzymes | CO2 Content | Species | Biomass | CO2 Utilization Efficiency | Reference |
|---|---|---|---|---|---|
| 100 mg/L MEA | 10% | Scenedesmus dimorphus | 0.293 g/(L·d) ↑ | 76.1% ↑ | [44] |
| 6 mmol/L THAM | 100% | Scenedesmus dimorphus | 11.57 g/(m2·d) ↑ | 35.58% ↑ | [45] |
| 2 mmol/L TEA | 4% | Scenedesmus sp. | 0.664 g/(L·d) ↑ | No data | [46] |
| 1.64 mol/L EDA + 0.41 mmol/L K2CO3 | 0.04% | Spirulina sp. LEB18 | 0.174 g/(L·d) ↑ | No data | [47] |
| 12% N-heptane | 15% | Chlorella sp. | 0.084 g/(L·d) ↑ | 64.7% ↑ | [48] |
| 100–150 mg/L MEA | 50% | Chlorella fusca LEB 111 | 0.096–0.122 g/(L·d) ↓ | 37% ↑ | [49] |
| 1 mmol/L TMEDA | 15% | Chlorella sp. L166 | 0.072 g/(L·d) ↑ | 43.29% ↑ | [50] |
| CA–GA beads | Air | Nannochloropsis salina | 0.040 g/(L·d) ↑ | No data | [13] |
| Immobilized CA on ElectrospunNanofibers | 15% | Dunaliella. tertiolecta ATCC 30929 | 6.8 × 105 cells/(mL·d) ↑ | No data | [64] |
| Metal–organic frameworks | 1.50% | Scenedesmus obliquus | 0.240 g/(L·d) ↑ | 21.6% ↑ | [65] |
| CA encapsulation using bamboo cellulose scaffolds | 5% | Chlorella vulgaris | 0.275 g/(L·d) ↑ | No data | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Bo, C.; Cao, S.; Sun, L. Enhancing CO2 Fixation in Microalgal Systems: Mechanistic Insights and Bioreactor Strategies. Mar. Drugs 2025, 23, 113. https://doi.org/10.3390/md23030113
Sun Z, Bo C, Cao S, Sun L. Enhancing CO2 Fixation in Microalgal Systems: Mechanistic Insights and Bioreactor Strategies. Marine Drugs. 2025; 23(3):113. https://doi.org/10.3390/md23030113
Chicago/Turabian StyleSun, Zhongliang, Chenmei Bo, Shuonan Cao, and Liqin Sun. 2025. "Enhancing CO2 Fixation in Microalgal Systems: Mechanistic Insights and Bioreactor Strategies" Marine Drugs 23, no. 3: 113. https://doi.org/10.3390/md23030113
APA StyleSun, Z., Bo, C., Cao, S., & Sun, L. (2025). Enhancing CO2 Fixation in Microalgal Systems: Mechanistic Insights and Bioreactor Strategies. Marine Drugs, 23(3), 113. https://doi.org/10.3390/md23030113