
New Insight into Utilization of Fish By-Product Proteins and Their Skin Health Promoting Effects




Abstract
1. Current Situation of Fish Production
2. Profiles of Fish by-Products
3. Utilization of Fish By-Products
4. Health Benefits of Fish By-Product Proteins
5. Pre-treatment of Fish By-Products
6. Extraction of Fish By-Product Proteins
7. Skin Health Promotion Effects of Fish By-Product Proteins
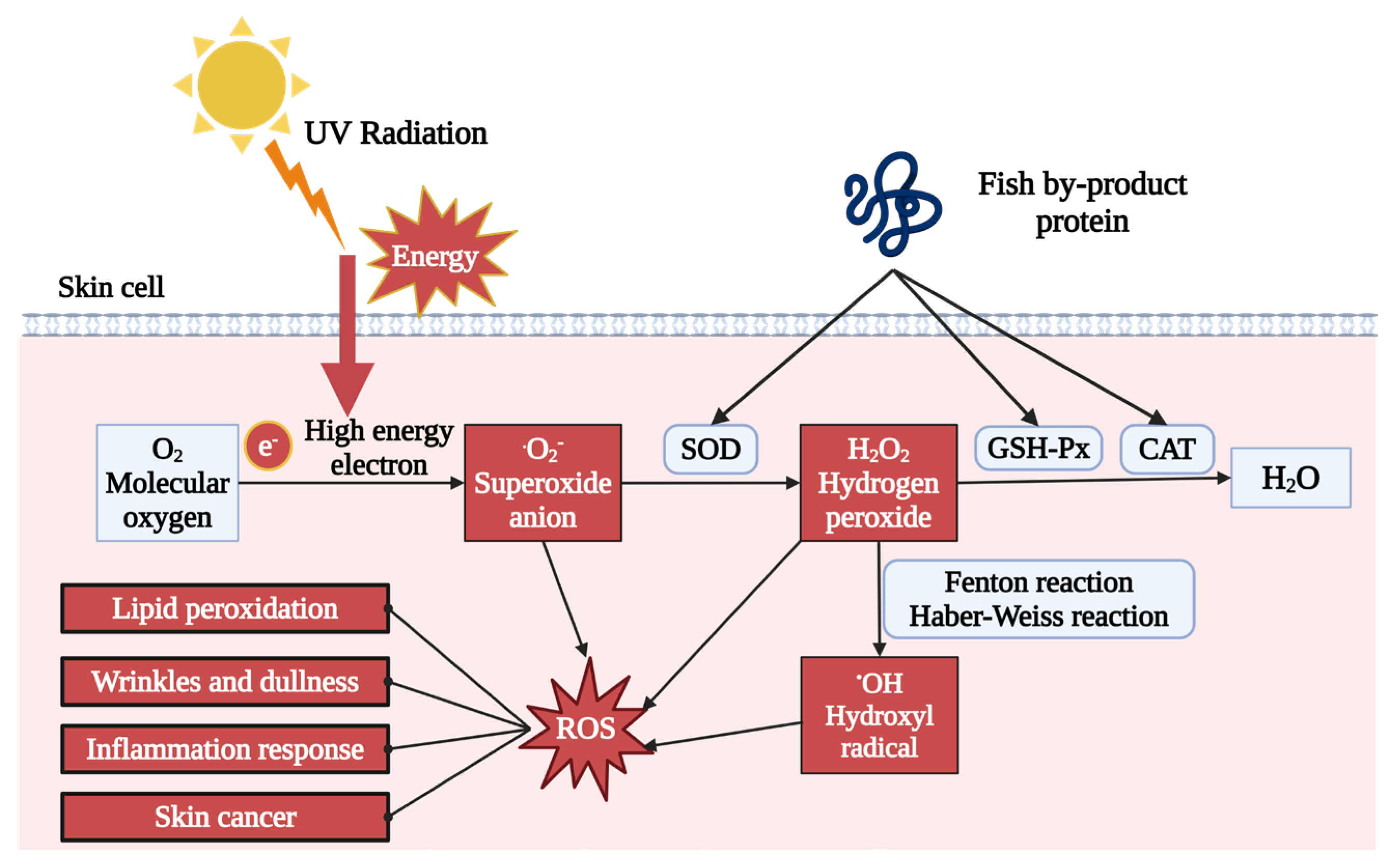
7.1. Fish By-Product Proteins for Skin Oxidative Stress Relief
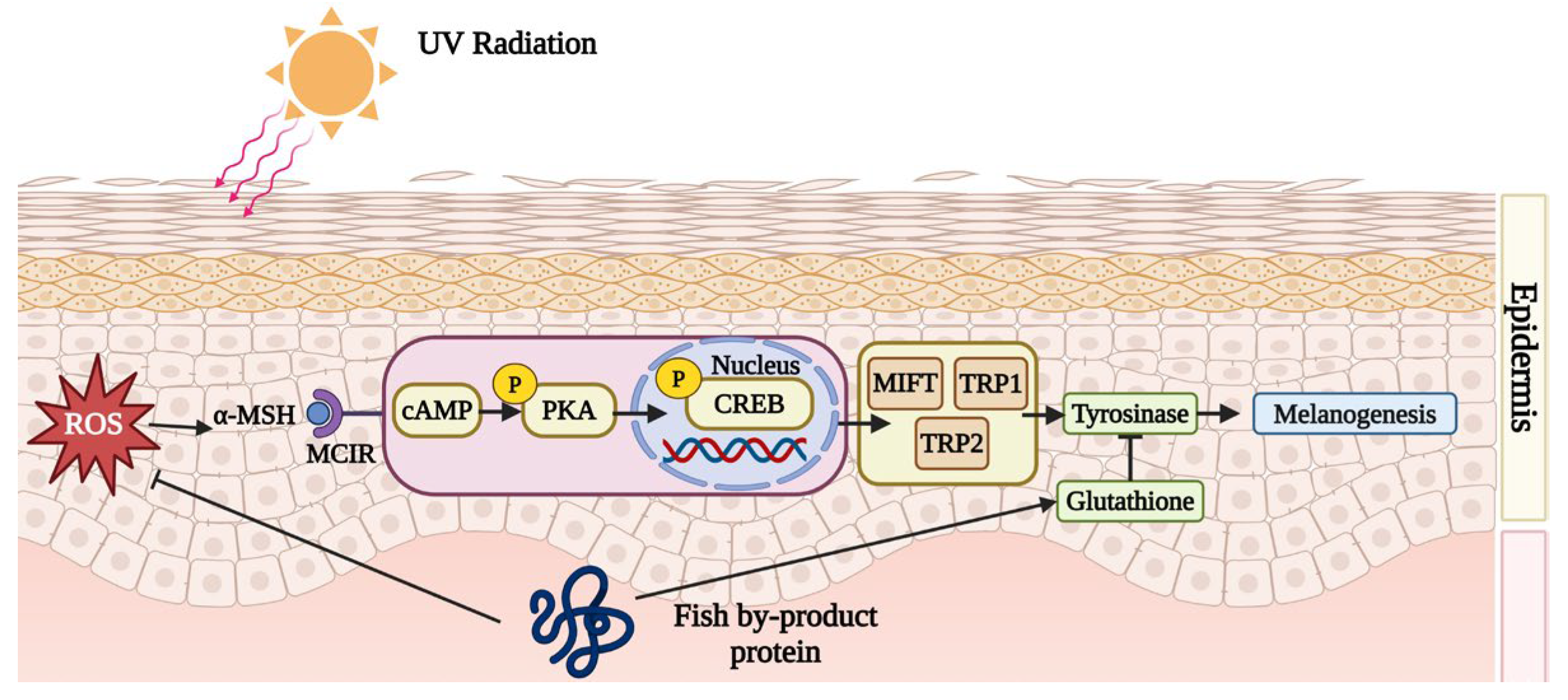
7.2. Melanogenesis Prevention by Fish By-Product Proteins
7.3. Improvement of Skin Hydration by Fish By-Product Proteins
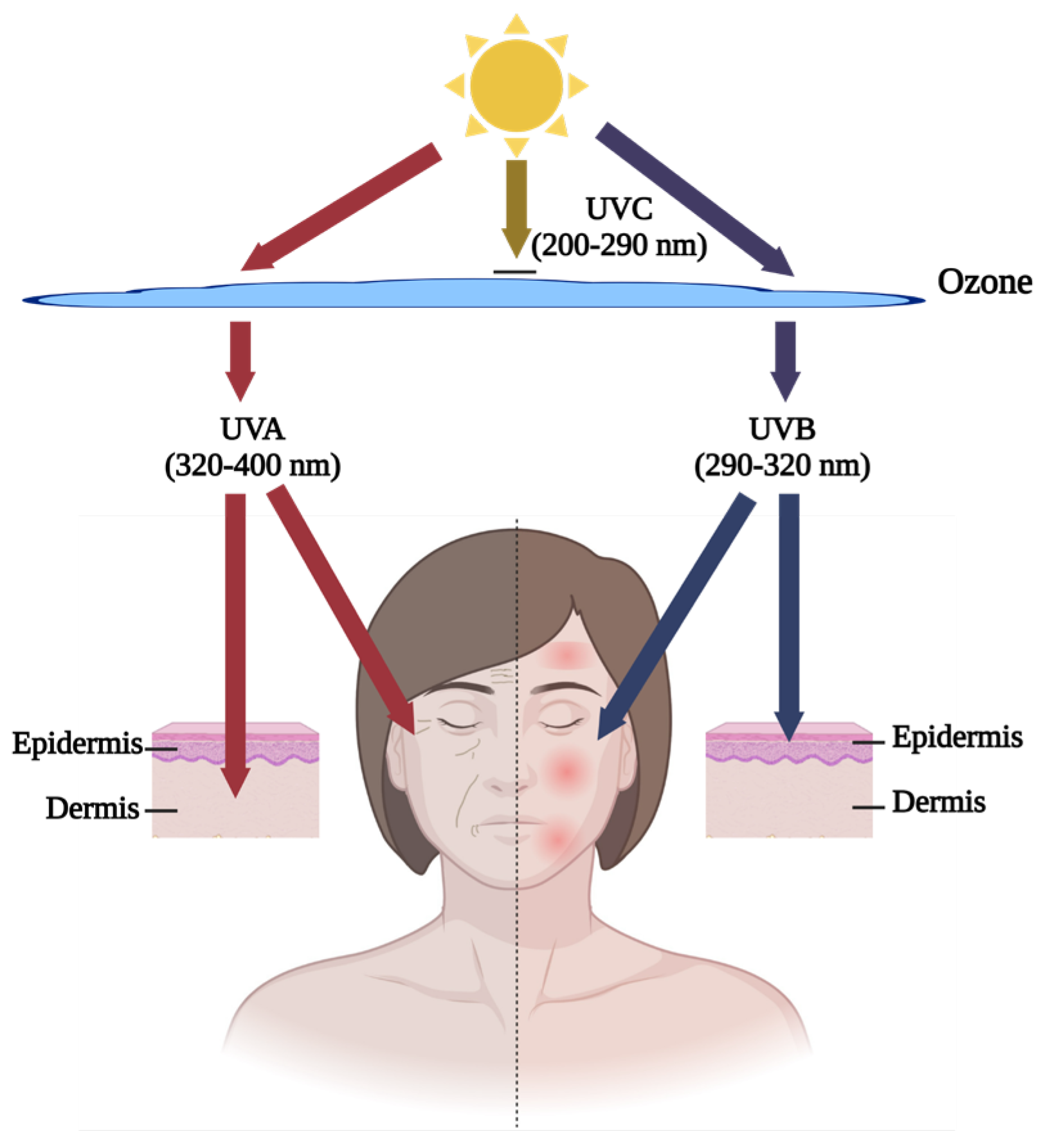
7.4. Influence of Fish By-Product Proteins on Photoaging-Induced DNA Damage
7.5. Impact on Skin Extracellular Matrix by Fish By-Product Proteins
7.6. Fish By-Product Proteins for Photoaging-Induced Skin Inflammatory
7.7. Gut Microbiota Balance and Fish By-Product Proteins
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Novais, C.; Molina, A.K.; Abreu, R.M.; Santo-Buelga, C.; Ferreira, I.C.; Pereira, C.; Barros, L. Natural food colorants and preservatives: A review, a demand, and a challenge. J. Agric. Food Chem. 2022, 70, 2789–2805. [Google Scholar] [CrossRef]
- Aakre, I.; Bøkevoll, A.; Chaira, J.; Bouthir, F.Z.; Frantzen, S.; Kausland, A.; Kjellevold, M. Variation in nutrient composition of seafood from North West Africa: Implications for food and nutrition security. Foods 2020, 9, 1516. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; Part 1. World review; FAO: Rome, Italy, 2022; p. 3. [Google Scholar]
- Salindeho, N.; Mokolensang, J.F.; Manu, L.; Taslim, N.A.; Nurkolis, F.; Gunawan, W.B.; Yusuf, M.; Mayulu, N.; Tsopmo, A. Fish scale rich in functional compounds and peptides: A potential nutraceutical to overcome undernutrition. Front. Nutr. 2022, 9, 1072370. [Google Scholar] [CrossRef] [PubMed]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M. Omega-3 polyunsaturated fatty acid (fish oil) supplementation and the prevention of clinical cardiovascular disease: A science advisory from the American Heart Association. Circulation 2017, 135, e867–e884. [Google Scholar] [CrossRef]
- Kruijssen, F.; Tedesco, I.; Ward, A.; Pincus, L.; Love, D.; Thorne-Lyman, A.L. Loss and waste in fish value chains: A review of the evidence from low and middle-income countries. Glob. Food Secur. 2020, 26, 100434. [Google Scholar] [CrossRef]
- Hou, Y.; Shavandi, A.; Carne, A.; Bekhit, A.A.; Ng, T.B.; Cheung, R.C.F.; Bekhit, A.E.-d.A. Marine shells: Potential opportunities for extraction of functional and health-promoting materials. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1047–1116. [Google Scholar] [CrossRef]
- Martínez, O.; Chamorro, S.; Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 2015, 73, 204–212. [Google Scholar] [CrossRef]
- Olsen, R.L.; Toppe, J.; Karunasagar, I. Challenges and realistic opportunities in the use of by-products from processing of fish and shellfish. Trends Food Sci. Technol. 2014, 36, 144–151. [Google Scholar] [CrossRef]
- Mutalipassi, M.; Esposito, R.; Ruocco, N.; Viel, T.; Costantini, M.; Zupo, V. Bioactive compounds of nutraceutical value from fishery and aquaculture discards. Foods 2021, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Takarina, N.; Fanani, A. Characterization of chitin and chitosan synthesized from red snapper (Lutjanus sp.) scale’s waste. In Proceedings of the AIP Conference Proceedings, Depok, Indonesia, 2 November 2017; p. 030108. [Google Scholar]
- Singhato, A.; Judprasong, K.; Sridonpai, P.; Laitip, N.; Ornthai, N.; Yafa, C.; Chimkerd, C. Effect of different cooking methods on selenium content of fish commonly consumed in Thailand. Foods 2022, 11, 1808. [Google Scholar] [CrossRef]
- Silva, C.M.; Lima, F.G.; Da Costa, K.F.; Ribeiro, V.A.; Leite, R.R.; Petenuci, M.; Gelinski, J.N.; Fonseca, G.G.; Prentice, C. Evaluation of enzymatic hydrolysis applied to fish by-product oil through chemical parameters. Ital. J. Food Sci. 2020, 32. [Google Scholar] [CrossRef]
- Radziemska, M.; Vaverková, M.D.; Adamcová, D.; Brtnický, M.; Mazur, Z. Valorization of fish waste compost as a fertilizer for agricultural use. Waste Biomass Valori. 2019, 10, 2537–2545. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Palma Esposito, F.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish waste: From problem to valuable resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Part 1. World Review; FAO: Rome, Italy, 2020; p. 6. [Google Scholar] [CrossRef]
- Yathisha, U.; Bhat, I.; Karunasagar, I.; Mamatha, B.S. Antihypertensive activity of fish protein hydrolysates and its peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, 2363–2374. [Google Scholar] [CrossRef]
- Chen, J.; Jayachandran, M.; Zhang, W.; Chen, L.; Du, B.; Yu, Z.; Xu, B. Dietary supplementation with sea bass (Lateolabrax maculatus) ameliorates ulcerative colitis and inflammation in macrophages through inhibiting toll-like receptor 4-linked pathways. Int. J. Mol. Sci. 2019, 20, 2907. [Google Scholar] [CrossRef]
- Khalili Tilami, S.; Sampels, S. Nutritional value of fish: Lipids, proteins, vitamins, and minerals. Rev. Fish. Sci. Aquac. 2018, 26, 243–253. [Google Scholar] [CrossRef]
- Heo, S.Y.; Ko, S.C.; Nam, S.Y.; Oh, J.; Kim, Y.M.; Kim, J.I.; Kim, N.; Yi, M.; Jung, W.K. Fish bone peptide promotes osteogenic differentiation of MC3T3-E1 pre-osteoblasts through upregulation of MAPKs and Smad pathways activated BMP-2 receptor. Cell Biochem. Funct. 2018, 36, 137–146. [Google Scholar] [CrossRef]
- Wei, Y.; Ye, Q.; Tang, Z.; Tian, G.; Zhu, Q.; Gao, H.; Wang, D.; Cao, Z. Calcitonin induces collagen synthesis and osteoblastic differentiation in human periodontal ligament fibroblasts. Arch. Oral Biol. 2017, 74, 114–122. [Google Scholar] [CrossRef]
- Salvatore, L.; Gallo, N.; Natali, M.L.; Campa, L.; Lunetti, P.; Madaghiele, M.; Blasi, F.S.; Corallo, A.; Capobianco, L.; Sannino, A. Marine collagen and its derivatives: Versatile and sustainable bio-resources for healthcare. Mater. Sci. Eng. C 2020, 113, 110963. [Google Scholar] [CrossRef]
- Halim, N.; Yusof, H.; Sarbon, N. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Kuo, J.-M.; Wu, S.-J.; Tsai, H.-T. Isolation and characterization of fish scale collagen from tilapia (Oreochromis sp.) by a novel extrusion–hydro-extraction process. Food Chem. 2016, 190, 997–1006. [Google Scholar] [CrossRef]
- He, Y.; Wang, J.; Si, Y.; Wang, X.; Deng, H.; Sheng, Z.; Li, Y.; Liu, J.; Zhao, J. A novel gene recombinant collagen hemostatic sponge with excellent biocompatibility and hemostatic effect. Int. J. Biol. Macromol. 2021, 178, 296–305. [Google Scholar] [CrossRef]
- Ramshaw, J.A.; Werkmeister, J.A.; Glattauer, V. Recent progress with recombinant collagens produced in Escherichia coli. Curr. Opin. Biomed. Eng. 2019, 10, 149–155. [Google Scholar] [CrossRef]
- Huang, Y.-R.; Shiau, C.-Y.; Chen, H.-H.; Huang, B.-C. Isolation and characterization of acid and pepsin-solubilized collagens from the skin of balloon fish (Diodon holocanthus). Food Hydrocoll. 2011, 25, 1507–1513. [Google Scholar] [CrossRef]
- Firdayanti, W.; Trilaksani, W.; Purwaningsih, S. Valorization of emperor fish (Lethrinus lentjan) skin for collagen isolation: Effect of acetic acid ratio and extraction time. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Online, 10 February 2023; p. 012047. [Google Scholar]
- Zhou, Y.; Yang, H. Effects of calcium ion on gel properties and gelation of tilapia (Oreochromis niloticus) protein isolates processed with pH shift method. Food Chem. 2019, 277, 327–335. [Google Scholar] [CrossRef]
- Furtado, M.; Chen, L.; Chen, Z.; Chen, A.; Cui, W. Development of fish collagen in tissue regeneration and drug delivery. Eng. Regen. 2022, 3, 217–231. [Google Scholar] [CrossRef]
- Tan, Y.; Chang, S.K. Isolation and characterization of collagen extracted from channel catfish (Ictalurus punctatus) skin. Food Chem. 2018, 242, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, B.; Song, W.; Si, L.; Hou, H. Characterization of pacific cod (Gadus macrocephalus) skin collagen and fabrication of collagen sponge as a good biocompatible biomedical material. Process Biochem. 2017, 63, 229–235. [Google Scholar] [CrossRef]
- Bhuimbar, M.V.; Bhagwat, P.K. Extraction and characterization of acid soluble collagen from fish waste: Development of collagen-chitosan blend as food packaging film. J. Environ. Chem. Eng. 2019, 7, 102983. [Google Scholar] [CrossRef]
- Muyonga, J.; Cole, C.; Duodu, K. Characterisation of acid soluble collagen from skins of young and adult nile perch (Lates niloticus). Food Chem. 2004, 85, 81–89. [Google Scholar] [CrossRef]
- Jeevithan, E.; Qingbo, Z.; Bao, B.; Wu, W. Biomedical and pharmaceutical application of fish collagen and gelatin: A review. J. Nutr. Ther. 2013, 2, 218–227. [Google Scholar] [CrossRef]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnacion, A.B.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Hemker, A.K.; Nguyen, L.T.; Karwe, M.; Salvi, D. Effects of pressure-assisted enzymatic hydrolysis on functional and bioactive properties of tilapia (Oreochromis niloticus) by-product protein hydrolysates. LWT Food Sci. Technol. 2020, 122, 109003. [Google Scholar] [CrossRef]
- Madende, M.; Hayes, M. Fish by-product use as biostimulants: An overview of the current state of the art, including relevant legislation and regulations within the EU and USA. Molecules 2020, 25, 1122. [Google Scholar] [CrossRef]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from fish by-product protein hydrolysates and its functional properties: An overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Azizi Khesal, M.; Sharifan, A.; Hoseini, E.; Ghavami, A. Optimization of enzymatic hydrolysis conditions of caspian kutum (Rutilus frisii kutum) by-product for production of bioactive peptides with antioxidative properties. Int. J. Pept. Res. Ther. 2020, 26, 1829–1838. [Google Scholar] [CrossRef]
- Robert, M.; Zatylny-Gaudin, C.; Fournier, V.; Corre, E.; Le Corguillé, G.; Bernay, B.; Henry, J. Molecular characterization of peptide fractions of a tilapia (Oreochromis niloticus) by-product hydrolysate and in vitro evaluation of antibacterial activity. Process Biochem. 2015, 50, 487–492. [Google Scholar] [CrossRef]
- Friedman, O. Changes associated with the aging face. Facial Plast. Surg. Clin. N. Am. 2005, 13, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.A.; Yoo, S.-K.; Seo, J.-S. Signatures of photo-aging and intrinsic aging in skin were revealed by transcriptome network analysis. Aging (Albany N.Y.) 2018, 10, 1609. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Del Giudice, R.; Rigano, M.M.; Monti, D.M. Antioxidants from plants protect against skin photoaging. Oxidative Med. Cell. Longev. 2018, 2018, 1454936. [Google Scholar] [CrossRef]
- Poon, F.; Kang, S.; Chien, A.L. Mechanisms and treatments of photoaging. Photodermatol. Photoimmunol. Photomed. 2015, 31, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-M.; Hong, C.-H.; Kang, S.-H.; Seo, D.-S.; Kim, S.-O.; Lee, H.-Y.; Sim, H.-J.; An, H.-J. Anti-photoaging effect of plant extract fermented with lactobacillus buchneri on ccd-986sk fibroblasts and HaCaT keratinocytes. J. Funct. Biomater. 2020, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Yang, S.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.-J. Trehalose against UVB-induced skin photoaging by suppressing MMP expression and enhancing procollagen I synthesis in HaCaT cells. J. Funct. Foods 2020, 74, 104198. [Google Scholar] [CrossRef]
- Jeong, E.H.; Yang, H.; Kim, J.-E.; Lee, K.W. Safflower seed oil and its active compound acacetin inhibit UVB-induced skin photoaging. J. Microbiol. Biotechnol. 2020, 30, 1567–1573. [Google Scholar] [CrossRef]
- Chen, B.; Yu, L.; Wu, J.; Qiao, K.; Cui, L.; Qu, H.; Su, Y.; Cai, S.; Liu, Z.; Wang, Q. Effects of collagen hydrolysate from large hybrid sturgeon on mitigating ultraviolet B-induced photodamage. Front. Bioeng. Biotechnol. 2022, 10, 908033. [Google Scholar] [CrossRef]
- Cho, W.; Park, J.; Lee, M.; Park, S.H.; Jung, J.; Kim, J.; Eun, S.; Kim, J. Gly-Pro-Val-Gly-Pro-Ser peptide fish collagen improves skin moisture and wrinkles with ameliorated the oxidative stress and pro-inflammatory factors in skin photoaging mimic models. Prev. Nutr. Food Sci. 2023, 28, 50–60. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, D.; Lee, M.; Yang, J.H.; Yang, J.S.; Lee, J. GT collagen improves skin moisturization in UVB-irradiated HaCaT cells and SKH-I hairless mice. J. Med. Food 2021, 24, 1313–1322. [Google Scholar] [CrossRef]
- Cho, C.H.; Lim, W.; Sim, W.-J.; Lim, T.-G. Oral administration of collagen peptide in SKH-1 mice suppress UVB-induced wrinkle and dehydration through MAPK and MAPKK signaling pathways, in vitro and in vivo evidence. Food Sci. Biotechnol. 2023, 33, 955–967. [Google Scholar] [CrossRef]
- Zhu, J.-J.; Huang, X.-N.; Yang, T.; Tang, C.-H.; Yin, S.-W.; Jia, X.-J.; Yang, X.-Q. An eco-friendly zein nanoparticle as robust cosmetic ingredient ameliorates skin photoaging. Ind. Crops Prod. 2022, 177, 114521. [Google Scholar] [CrossRef]
- Balupillai, A.; Kanimozhi, G.; Khan, H.A.; Alhomida, A.S.; Prasad, N.R. Opuntiol prevents photoaging of mouse skin via blocking inflammatory responses and collagen degradation. Oxidative Med. Cell. Longev. 2020, 2020, 5275178. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, M.; Yang, J.H.; Yang, J.S.; Kim, O.-K. Dual skin-whitening and anti-wrinkle function of low-molecular-weight fish collagen. J. Med. Food 2022, 25, 192–204. [Google Scholar] [CrossRef]
- Lee, M.; Kim, D.; Park, S.H.; Jung, J.; Cho, W.; Yu, A.R.; Lee, J. Fish collagen peptide (Naticol®) protects the skin from dryness, wrinkle formation, and melanogenesis both in vitro and in vivo. Prev. Nutr. Food Sci. 2022, 27, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Maia Campos, P.M.B.G.; Franco, R.S.B.; Kakuda, L.; Cadioli, G.F.; Costa, G.M.D.A.; Bouvret, E. Oral supplementation with hydrolyzed fish cartilage improves the morphological and structural characteristics of the skin: A double-blind, placebo-controlled clinical study. Molecules 2021, 26, 4880. [Google Scholar] [CrossRef]
- Cai, W.-W.; Hu, X.-M.; Wang, Y.-M.; Chi, C.-F.; Wang, B. Bioactive peptides from skipjack tuna cardiac arterial bulbs: Preparation, identification, antioxidant activity, and stability against thermal, pH, and simulated gastrointestinal digestion treatments. Mar. Drugs 2022, 20, 626. [Google Scholar] [CrossRef]
- Kong, J.; Hu, X.-M.; Cai, W.-W.; Wang, Y.-M.; Chi, C.-F.; Wang, B. Bioactive peptides from skipjack tuna cardiac arterial bulbs (II): Protective function on UVB-irradiated HaCaT cells through antioxidant and anti-apoptotic mechanisms. Mar. Drugs 2023, 21, 105. [Google Scholar] [CrossRef]
- Iosageanu, A.; Ilie, D.; Craciunescu, O.; Seciu-Grama, A.-M.; Oancea, A.; Zarnescu, O.; Moraru, I.; Oancea, F. Effect of fish bone bioactive peptides on oxidative, inflammatory and pigmentation processes triggered by UVB irradiation in skin cells. Molecules 2021, 26, 2691. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Liang, C.-H.; Wu, H.-T.; Pang, H.-Y.; Chen, C.; Wang, G.-H.; Chan, L.-P. Antioxidant and anti-inflammatory capacities of collagen peptides from milkfish (Chanos chanos) scales. J. Food Sci. Technol. 2018, 55, 2310–2317. [Google Scholar] [CrossRef]
- Li, H.-L.; Li, M.-J.; Xiong, G.-Q.; Cai, J.; Liao, T.; Zu, X.-Y. Silver carp (Hypophthalmichthys molitrix) scale collagen peptides-1 (SCPs1) inhibit melanogenesis through downregulation of the cAMP-CREB signaling pathway. Nutrients 2023, 15, 2449. [Google Scholar] [CrossRef]
- Helton, E.S.; Chen, X. P53 modulation of the DNA damage response. J. Cell. Biochem. 2007, 100, 883–896. [Google Scholar] [CrossRef]
- Williams, A.B.; Schumacher, B. P53 in the DNA-damage-repair process. Cold Spring Harb. Perspect. Med. 2016, 6, a026070. [Google Scholar] [CrossRef]
- Chen, T.; Hou, H.; Fan, Y.; Wang, S.; Chen, Q.; Si, L.; Li, B. Protective effect of gelatin peptides from pacific cod skin against photoaging by inhibiting the expression of MMPs via MAPK signaling pathway. J. Photochem. Photobiol. B Biol. 2016, 165, 34–41. [Google Scholar] [CrossRef]
- Xu, D.; Li, C.; Zhao, M. Attenuation of UV-induced skin photoaging in rats by walnut protein hydrolysates is linked to the modulation of MAPK/ap-1 and TGF-β/Smad signaling pathways. Food Funct. 2022, 13, 609–623. [Google Scholar] [CrossRef]
- Zhan, J.Y.-X.; Wang, X.-F.; Liu, Y.-H.; Zhang, Z.-B.; Wang, L.; Chen, J.-N.; Huang, S.; Zeng, H.-F.; Lai, X.-P. Andrographolide sodium bisulfate prevents UV-induced skin photoaging through inhibiting oxidative stress and inflammation. Mediat. Inflamm. 2016, 2016, 3271451. [Google Scholar] [CrossRef]
- Lin, R.-F.; Feng, X.-X.; Li, C.-W.; Zhang, X.-J.; Yu, X.-T.; Zhou, J.-Y.; Zhang, X.; Xie, Y.-L.; Su, Z.-R.; Zhan, J.Y.-X. Prevention of UV radiation-induced cutaneous photoaging in mice by topical administration of patchouli oil. J. Ethnopharmacol. 2014, 154, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Paternò, E.J.; Candi, E.; Falconi, M.; Costanza, G.; Diluvio, L.; Terrinoni, A.; Bianchi, L.; Orlandi, A. The relevance of piroxicam for the prevention and treatment of nonmelanoma skin cancer and its precursors. Drug Des. Dev. Ther. 2015, 9, 5843–5850. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, B.; Volpert, O.V.; Crawford, S.E.; Febbraio, M.; Silverstein, R.L.; Bouck, N. Signals leading to apoptosis-dependent inhibition of neovascularization by thrombospondin-1. Nat. Med. 2000, 6, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Amano, S. Characterization and mechanisms of photoageing-related changes in skin. Damages of basement membrane and dermal structures. Exp. Dermatol. 2016, 25, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Zeeuwen, P.L.; Boekhorst, J.; van den Bogaard, E.H.; de Koning, H.D.; van de Kerkhof, P.M.; Saulnier, D.M.; van Swam, I.I.; van Hijum, S.A.; Kleerebezem, M.; Schalkwijk, J. Microbiome dynamics of human epidermis following skin barrier disruption. Genome Biol. 2012, 13, R101. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pellicer, P.; Navarro-Moratalla, L.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Agüera-Santos, J.; Navarro-López, V. Acne, microbiome, and probiotics: The gut–skin axis. Microorganisms 2022, 10, 1303. [Google Scholar] [CrossRef]
- Qu, L.; Ma, X.; Wang, F. The roles of gut microbiome and metabolites associated with skin photoaging in mice by intestinal flora sequencing and metabolomics. Life Sci. 2024, 341, 122487. [Google Scholar] [CrossRef]
- Geng, R.; Kang, S.-G.; Huang, K.; Tong, T. Dietary isoeugenol supplementation attenuates chronic UVB-induced skin photoaging and modulates gut microbiota in mice. Nutrients 2024, 16, 481. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Song, H.H.; Kim, H.; Kim, B.Y.; Park, S.; Suh, H.J.; Ahn, Y. Oral administration of mixed probiotics improves photoaging by modulating the cecal microbiome and MAPK pathway in UVB-irradiated hairless mice. Mol. Nutr. Food Res. 2023, 67, e2200841. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, J.; Selma-Royo, M.; Simal-Gandara, J.; Collado, M.C.; Barba, F.J. Potential benefits of high-added-value compounds from aquaculture and fish side streams on human gut microbiota. Trends Food Sci. Technol. 2021, 112, 484–494. [Google Scholar] [CrossRef]
- Mei, F.; Duan, Z.; Chen, M.; Lu, J.; Zhao, M.; Li, L.; Shen, X.; Xia, G.; Chen, S. Effect of a high-collagen peptide diet on the gut microbiota and short-chain fatty acid metabolism. J. Funct. Foods 2020, 75, 104278. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Model | Molecular Targets | Biological Activity | References |
|---|---|---|---|---|
| Scale of Nile tilapia (O. niloticus) | UVB-induced HaCaT UVB-induced HS27 UVB-induced SKH-1 mice | ↑: SOD, CAT, GSH-Px | ↑: Antioxidant capacity | [50] |
| ↑: TGF-βRI, Smad3, Collagen I, Pro-collagen I | ↑: ECM synthesis | |||
| ↑: HAS (1–3), LCB1(SPT), Elastin, Fibrillin-1, Hyaluronic acid | ↑: Skin hydration | |||
| ↑: CerS4, DEGS1, Sphingomyelin | ↓: Moisture loss | |||
| ↓: JNK, c-Fos, c-Jun, MMP-(1,2,9) | ↓: ECM breakdown | |||
| ↓: IκB, P65, COX-2, TNF-α, IL-1β, IL-6 | ↓: Inflammation | |||
| Scale of Nile tilapia (O. niloticus) | UVB-induced HaCaT UVB-induced SKH-1 mice | ↑: LCB1, GLcNAc, UGTrel7, Elastin, HAS2, Hyaluronic acid | ↑: Skin hydration | [51] |
| ↑: CerS4, DEGS1, Sphingomyelin | ↓: Moisture loss | |||
| Skin of pangasius (P. bocourti) and tilapia (O. niloticus) | UVB-induced HaCaT UVB-induced HDF UVB-induced SKH-1 mice | ↑: HAS2; COL1A1, HA; ↓: HYAL2, Elastase | ↑: Skin hydration | [52] |
| ↑: SOD, GSH-Px, MDA; ↓: ROS | ↑: Antioxidant capacity | |||
| ↓: ERK, p38, JNK, MEK1/2,3,4, MMP-1,13 | ↓: ECM breakdown | |||
| Scale of Nile tilapia (O. niloticus) | UVB-induced Hs27 IBMX-induced B16F10 UVB-induced SKH-1 mice | ↑: GSH; ↓: PKA, cAMP, CREB, MITF, TRP-1, TRP-2, Melanin | ↓: Melanogenesis | [55] |
| ↑: SOD, GSH-Px, CAT | ↑: Antioxidant capacity | |||
| ↑: TGF-βRI, Smad3, Pro-collagen I, Collagen I | ↑: ECM synthesis | |||
| ↓: JNK, c-Fos, c-Jun, MMP-1,3,9 | ↓: ECM breakdown | |||
| ↓: IL-1β, IL-6, TNF-α, NO | ↓: Inflammation | |||
| Skin of Nile tilapia (O. niloticus) | UVB-induced HaCaT UVB-induced Hs27 IBMX-induced B16F10 UVB-induced SKH-1 mice | ↑: HAS1–3, LCB1(SPT), DEGS1, Fibrillin-1, CerS4, Hyaluronic acid, Sphingomyelin | ↑: Skin hydration | [56] |
| ↑: GSH; ↓: cAMP, MITF, TRP-1,2, PKA, CREB | ↓: Melanogenesis | |||
| ↑: SOD, CAT, GSH-Px | ↑: Antioxidant capacity | |||
| ↑: TGF-βRI, Smad3, Pro-collagen I, Collagen I | ↑: ECM synthesis | |||
| ↓: JNK, c-Fos, c-Jun, MMP-1,3,9 | ↓: ECM breakdown | |||
| ↓: Iκ-B, P65, COX-2, IL-1β, IL-6, TNF-α | ↓: Inflammation |
| Source | Model | Molecular Targets | Biological Activity | References |
|---|---|---|---|---|
| Cardiac arterial bulbs of skipjack tuna (K. pelamis) | UVB-induced HaCaT | ↑: SOD, CAT, GSH-Px, MDA, Mitochondrial membrane potential; ↓: ROS | ↑: Antioxidant capacity | [58,59] |
| ↑: Nrf2, HO-1, NQO1, Bcl-2. ↓: Cleaved-caspase-3,8,9, Bax | ↑: Antioxidant capacity | |||
| ↓: Hoechst 33342 Staining | ↓: DNA damage | |||
| Bone of silver carp (H. molitrix) | UVB-induced HaCaT UVB-induced L929 Human melanoma | ↓: ROS, MDA | ↑: Antioxidant capacity | [60] |
| ↓: TNF-α, IL-1β | ↓: Inflammation | |||
| ↓: Tyrosinase Activity, Melanin | ↓: Melanogenesis | |||
| Skin of large hybrid sturgeon (H. Dauricus) | UVB-induced L929 UVB-induced Zebrafish Embryo | ↑: Pro-collagen I | ↑: ECM synthesis | [49] |
| ↓: ROS, MDA, | ↑: Antioxidant capacity | |||
| ↓: IL-1β, IL-6, TNF-α, Cox-2 | ↑: Antioxidant capacity | |||
| ↓: p38, Erk1/2, Jnk1/2/3, c-Jun, MMP-1,2,3 | ↓: ECM breakdown | |||
| ↓: Hoechst 33342 Staining | ↓: DNA damage | |||
| Scale of milkfish (C. Chanos) | UV-induced HaCaT UV-induced plasmid | ↑: O-form, S-form plasmid | ↓: DNA damage | [61] |
| ↓: iNOS, NO | ↓: Inflammation | |||
| ↓: DPPH, ABTS, ROS | ↑: Antioxidant capacity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Ren, Y.; Zhong, S.; Xu, B. New Insight into Utilization of Fish By-Product Proteins and Their Skin Health Promoting Effects. Mar. Drugs 2024, 22, 215. https://doi.org/10.3390/md22050215
Liu D, Ren Y, Zhong S, Xu B. New Insight into Utilization of Fish By-Product Proteins and Their Skin Health Promoting Effects. Marine Drugs. 2024; 22(5):215. https://doi.org/10.3390/md22050215
Chicago/Turabian StyleLiu, Dongcheng, Yongxin Ren, Saiyi Zhong, and Baojun Xu. 2024. "New Insight into Utilization of Fish By-Product Proteins and Their Skin Health Promoting Effects" Marine Drugs 22, no. 5: 215. https://doi.org/10.3390/md22050215
APA StyleLiu, D., Ren, Y., Zhong, S., & Xu, B. (2024). New Insight into Utilization of Fish By-Product Proteins and Their Skin Health Promoting Effects. Marine Drugs, 22(5), 215. https://doi.org/10.3390/md22050215