
Bioprospection of the Antarctic Diatoms Craspedostauros ineffabilis IMA082A and Craspedostauros zucchelli IMA088A


Abstract

1. Introduction
2. Results and Discussion
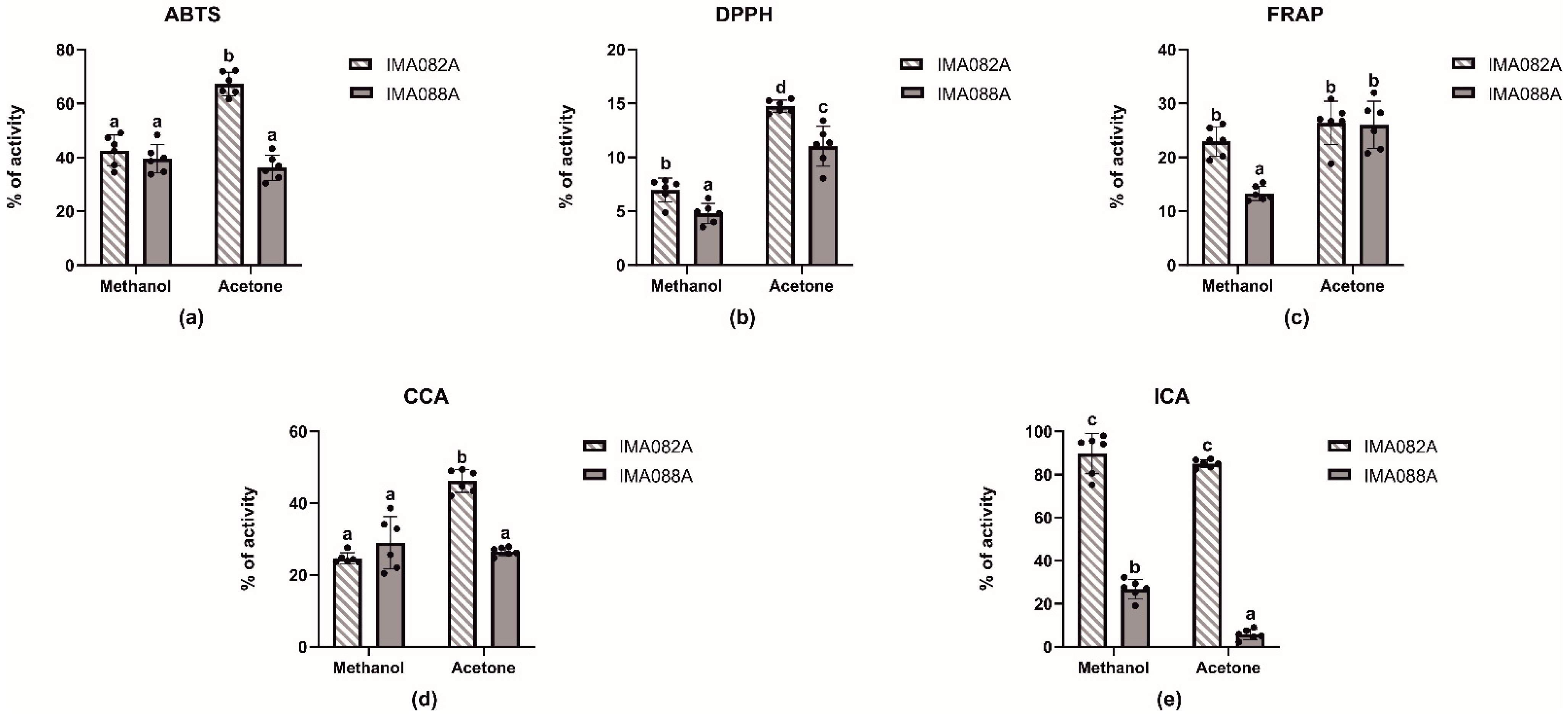
2.1. In Vitro Antioxidant Properties
2.2. Enzymatic Inhibitory Properties of the Extracts
2.3. Chemical Profile
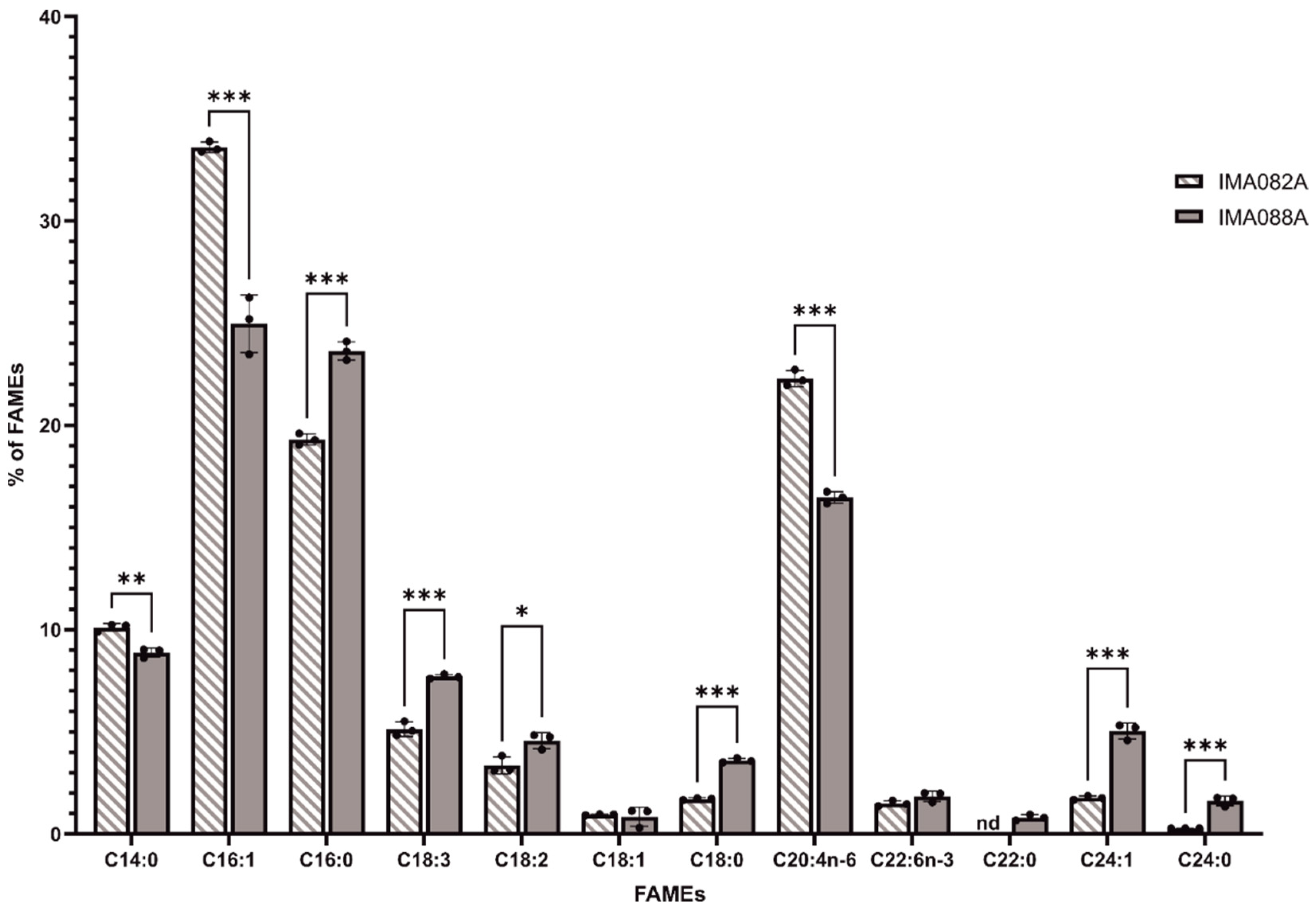
2.4. FAMEs Profile
2.5. Fucoxanthin Content
2.6. Discussion
3. Materials and Methods
3.1. Chemicals
3.2. Biomass Collection and Preparation of the Extracts
3.3. In Vitro Antioxidant Properties
3.3.1. RSA on DPPH• Radical
3.3.2. RSA on ABTS•+ Radical
3.3.3. FRAP
3.3.4. CCA
3.3.5. ICA
3.4. Enzyme Inhibition Assays
3.4.1. AChE and BChE Inhibition
3.4.2. α–Amylase Inhibition
3.4.3. α–Glucosidase Inhibition
3.4.4. Lipase Inhibition
3.4.5. Tyrosinase Inhibition
3.5. UHPLC–HR–MS/MS Profiling of the Extracts
3.6. Fatty Acid Methyl Esters Profiling
3.6.1. Lipids Extraction and Transesterification
3.6.2. Determination of FAMEs Profile by GC–MS
3.7. Fucoxanthin Spectrophotometric Quantification
3.8. Data Presentation and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abida, H.; Ruchaud, S.; Rios, L.; Humeau, A.; Probert, I.; De Vargas, C.; Bach, S.; Bowler, C. Bioprospecting Marine Plankton. Mar. Drugs 2013, 11, 4594–4611. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Dhakar, K.; Joshi, A. Alkaliphiles: Diversity and Bioprospection. In Microbial Diversity in the Genomic Era; Academic Press: Cambridge, MA, USA, 2019; pp. 239–263. [Google Scholar] [CrossRef]
- Hosseini, H.; Al-Jabri, H.M.; Moheimani, N.R.; Siddiqui, S.A.; Saadaoui, I. Marine Microbial Bioprospecting: Exploitation of Marine Biodiversity towards Biotechnological Applications—A Review. J. Basic Microbiol. 2022, 62, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ashforth, E.; Ren, B.; Song, F.; Dai, H.; Liu, M.; Wang, J.; Xie, Q.; Zhang, L. Bioprospecting Microbial Natural Product Libraries from the Marine Environment for Drug Discovery. J. Antibiot. 2010, 63, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bloch, K.; Webster, T.J. Bioprospecting of Novel Algal Species with Nanobiotechnology. In An Integration of Phycoremediation Processes in Wastewater Treatment; Elsevier: Amsterdam, The Netherlands, 2022; pp. 41–74. [Google Scholar] [CrossRef]
- Bhadury, P.; Wright, P.C. Exploitation of Marine Algae: Biogenic Compounds for Potential Antifouling Applications. Planta 2004, 219, 561–578. [Google Scholar] [CrossRef] [PubMed]
- Santhakumaran, P.; Kookal, S.K.; Mathew, L.; Ray, J.G. Bioprospecting of Three Rapid-Growing Freshwater Green Algae, Promising Biomass for Biodiesel Production. Bioenergy Res. 2019, 12, 680–693. [Google Scholar] [CrossRef]
- Mutanda, T.; Ramesh, D.; Karthikeyan, S.; Kumari, S.; Anandraj, A.; Bux, F. Bioprospecting for Hyper-Lipid Producing Microalgal Strains for Sustainable Biofuel Production. Bioresour. Technol. 2011, 102, 57–70. [Google Scholar] [CrossRef]
- Boukid, F.; Castellari, M. Food and Beverages Containing Algae and Derived Ingredients Launched in the Market from 2015 to 2019: A Front-of-Pack Labeling Perspective with a Special Focus on Spain. Foods 2021, 10, 173. [Google Scholar] [CrossRef]
- Europe Algae Products Market Size, Share, Scope & Forecast by 2030. Available online: https://www.databridgemarketresearch.com/reports/europe-algae-products-market (accessed on 31 July 2023).
- Mosey, M.; Douchi, D.; Knoshaug, E.P.; Laurens, L.M.L. Methodological Review of Genetic Engineering Approaches for Non-Model Algae. Algal. Res. 2021, 54, 102221. [Google Scholar] [CrossRef]
- Giordano, D. Bioactive Molecules from Extreme Environments. Mar. Drugs 2020, 18, 640. [Google Scholar] [CrossRef]
- Trentin, R.; Negrisolo, E.; Moschin, E.; Veronese, D.; Cecchetto, M.; Moro, I. Microglena antarctica Sp. Nov. a New Antarctic Green Alga from Inexpressible Island (Terra Nova Bay, Ross Sea) Revealed through an Integrative Approach. Diversity 2022, 14, 337. [Google Scholar] [CrossRef]
- Trentin, R.; Moschin, E.; Duarte Lopes, A.; Schiaparelli, S.; Custódio, L.; Moro, I. Molecular, Morphological and Chemical Diversity of Two New Species of Antarctic Diatoms, Craspedostauros ineffabilis Sp. Nov. and Craspedostauros zucchellii Sp. Nov. J. Mar. Sci. Eng. 2022, 10, 1656. [Google Scholar] [CrossRef]
- Trentin, R.; Moschin, E.; Grapputo, A.; Rindi, F.; Schiaparelli, S.; Moro, I. Multi-Gene Phylogeny Reveals a New Genus and Species of Hapalidiales (Rhodophyta) from Antarctica: Thalassolithon adeliense Gen. & Sp. Nov. Phycologia 2023, 62, 83–98. [Google Scholar] [CrossRef]
- Sciuto, K.; Moschin, E.; Alongi, G.; Cecchetto, M.; Schiaparelli, S.; Caragnano, A.; Rindi, F.; Moro, I. Tethysphytum antarcticum Gen. et Sp. Nov. (Hapalidiales, Rhodophyta), a New Non-Geniculate Coralline Alga from Terra Nova Bay (Ross Sea, Antarctica): Morpho-Anatomical Characterization and Molecular Phylogeny. Eur. J. Phycol. 2021, 56, 416–427. [Google Scholar] [CrossRef]
- Lyon, B.R.; Mock, T. Polar Microalgae: New Approaches towards Understanding Adaptations to an Extreme and Changing Environment. Biology 2014, 3, 56–80. [Google Scholar] [CrossRef] [PubMed]
- Petrou, K.; Doblin, M.A.; Ralph, P.J. Heterogeneity in the Photoprotective Capacity of Three Antarctic Diatoms during Short-Term Changes in Salinity and Temperature. Mar. Biol. 2011, 158, 1029–1041. [Google Scholar] [CrossRef]
- Ferruzzi, M.G.; Failla, M.L.; Schwartz, S.J. Assessment of Degradation and Intestinal Cell Uptake of Carotenoids and Chlorophyll Derivatives from Spinach Puree Using an Invitro Digestion and Caco-2 Human Cell Model. J. Agric. Food Chem. 2001, 49, 2082–2089. [Google Scholar] [CrossRef] [PubMed]
- Queiroz Zepka, L.; Jacob-Lopes, E.; Roca, M. Catabolism and Bioactive Properties of Chlorophylls. Curr. Opin. Food Sci. 2019, 26, 94–100. [Google Scholar] [CrossRef]
- Islam, M.N.; Ishita, I.J.; Jin, S.E.; Choi, R.J.; Lee, C.M.; Kim, Y.S.; Jung, H.A.; Choi, J.S. Anti-Inflammatory Activity of Edible Brown Alga Saccharina Japonica and Its Constituents Pheophorbide a and Pheophytin a in LPS-Stimulated RAW 264.7 Macrophage Cells. Food Chem. Toxicol. 2013, 55, 541–548. [Google Scholar] [CrossRef]
- Custódio, L.; Justo, T.; Silvestre, L.; Barradas, A.; Duarte, C.V.; Pereira, H.; Barreira, L.; Rauter, A.P.; Alberício, F.; Varela, J. Microalgae of Different Phyla Display Antioxidant, Metal Chelating and Acetylcholinesterase Inhibitory Activities. Food Chem. 2012, 131, 134–140. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Alejandro Cifuentes, A.; Ibáñez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Fu, W.; Wichuk, K.; Brynjólfsson, S. Developing Diatoms for Value-Added Products: Challenges and Opportunities. N. Biotechnol. 2015, 32, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Pikula, K.S.; Zakharenko, A.M.; Aruoja, V.; Golokhvast, K.S.; Tsatsakis, A.M. Oxidative Stress and Its Biomarkers in Microalgal Ecotoxicology. Curr. Opin. Toxicol. 2019, 13, 8–15. [Google Scholar] [CrossRef]
- Dahmen-Ben Moussa, I.; Chtourou, H.; Karray, F.; Sayadi, S.; Dhouib, A. Nitrogen or Phosphorus Repletion Strategies for Enhancing Lipid or Carotenoid Production from Tetraselmis Marina. Bioresour. Technol. 2017, 238, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Xiudong, Y.; Min-Cheol, K.; Ki-Wan, L.; Sung-Myung, K.; Won-Woo, L.; You-Jin, J. Antioxidant Activity and Cell Protective Effect of Loliolide Isolated from Sargassum Ringgoldianum Subsp. Coreanum. ALGAE 2011, 26, 201–208. [Google Scholar] [CrossRef]
- Percot, A.; Yalçın, A.; Aysel, V.; Erduğan, H.; Dural, B.; Güven, K.C. Loliolide in Marine Algae. Nat. Prod. Res. 2009, 23, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Hulkko, L.S.S.; Rocha, R.M.; Trentin, R.; Fredsgaard, M.; Chaturvedi, T.; Custódio, L.; Thomsen, M.H. Bioactive Extracts from Salicornia ramosissima J. Woods Biorefinery as a Source of Ingredients for High-Value Industries. Plants 2023, 12, 1251. [Google Scholar] [CrossRef]
- Hassan, M.; Raza, H.; Abbasi, M.A.; Moustafa, A.A.; Seo, S.Y. The Exploration of Novel Alzheimer’s Therapeutic Agents from the Pool of FDA Approved Medicines Using Drug Repositioning, Enzyme Inhibition and Kinetic Mechanism Approaches. Biomed. Pharmacother. 2019, 109, 2513–2526. [Google Scholar] [CrossRef]
- Pagano, G.; Rengo, G.; Pasqualetti, G.; Femminella, G.D.; Monzani, F.; Ferrara, N.; Tagliati, M. Cholinesterase Inhibitors for Parkinson’s Disease: A Systematic Review and Meta-Analysis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 767–773. [Google Scholar] [CrossRef]
- Kawee-ai, A.; Kuntiya, A.; Kim, S.M. Anticholinesterase and Antioxidant Activities of Fucoxanthin Purified from the Microalga Phaeodactylum tricornutum. Nat. Prod. Commun. 2013, 8, 1381–1386. [Google Scholar] [CrossRef]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase In Vitro. Mar. Drugs 2016, 14, 67. [Google Scholar] [CrossRef]
- He, S.; Yang, M.; Xuan, Z.; Wang, Q.; Yan, S.; Zhou, D.; Naman, C.B.; Zhang, J.; Yan, X.; Cui, W.; et al. Fucoxanthin Has Potential for Therapeutic Efficacy in Neurodegenerative Disorders by Acting on Multiple Targets. Nutr. Neurosci. 2021, 25, 2167–2180. [Google Scholar] [CrossRef]
- Fang, Z.; Jeong, S.Y.; Jung, H.A.; Choi, J.S.; Min, B.S.; Woo, M.H. Anticholinesterase and Antioxidant Constituents from Gloiopeltis Furcata. Chem. Pharm. Bull. 2010, 58, 1236–1239. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Narwal, S.; Kumar, V.; Prakash, O. α-Glucosidase Inhibitors from Plants: A Natural Approach to Treat Diabetes. Pharmacogn. Rev. 2011, 5, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, P.; Surendran, L.; Sudhagar, B.; Ranjith Santhosh Kumar, D.S. Facile Green Synthesis of Gold Nanoparticles from Marine Algae Gelidiella Acerosa and Evaluation of Its Biological Potential. SN Appl. Sci. 2019, 1, 284. [Google Scholar] [CrossRef]
- Liu, B.; Kongstad, K.T.; Wiese, S.; Jäger, A.K.; Staerk, D. Edible Seaweed as Future Functional Food: Identification of α-Glucosidase Inhibitors by Combined Use of High-Resolution α-Glucosidase Inhibition Profiling and HPLC–HRMS–SPE–NMR. Food Chem. 2016, 203, 16–22. [Google Scholar] [CrossRef]
- Leporini, M.; Loizzo, M.R.; Tundis, R.; La Torre, C.; Fazio, A.; Plastina, P. Non-Pungent n-3 Polyunsaturated Fatty Acid (PUFA)-Derived Capsaicin Analogues as Potential Functional Ingredients with Antioxidant and Carbohydrate-Hydrolysing Enzyme Inhibitory Activities. Antioxidants 2019, 8, 162. [Google Scholar] [CrossRef]
- Sergent, T.; Vanderstraeten, J.; Winand, J.; Beguin, P.; Schneider, Y.J. Phenolic Compounds and Plant Extracts as Potential Natural Anti-Obesity Substances. Food Chem. 2012, 135, 68–73. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hosokawa, M.; Matsukawa, N.; Hagio, M.; Shinoki, A.; Nishimukai, M.; Miyashita, K.; Yajima, T.; Hara, H. Suppressive Effects of the Marine Carotenoids, Fucoxanthin and Fucoxanthinol on Triglyceride Absorption in Lymph Duct-Cannulated Rats. Eur. J. Nutr. 2010, 49, 243–249. [Google Scholar] [CrossRef]
- Azmi, N.; Hashim, P.; Hashim, D.M.; Halimoon, N.; Nik Majid, N.M. Anti-Elastase, Anti-Tyrosinase and Matrix Metalloproteinase-1 Inhibitory Activity of Earthworm Extracts as Potential New Anti-Aging Agent. Asian Pac. J. Trop. Biomed. 2014, 4, S348–S352. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A Comprehensive Review on Tyrosinase Inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for Bioactive Compounds from Algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Park, D.-C.; Ji, C.-I.; Kim, S.-H.; Jung, K.-J.; Lee, T.-G.; Kim, I.-S.; Park, Y.-H.; Kim, S.-B. Characteristics of Tyrosinase Inhibitory Extract from Ecklonia stolonifera. Fish Aquat. Sci. 2000, 3, 195–199. [Google Scholar]
- Lopes, D.; Rey, F.; Leal, M.C.; Lillebø, A.I.; Calado, R.; Domingues, M.R. Bioactivities of Lipid Extracts and Complex Lipids from Seaweeds: Current Knowledge and Future Prospects. Mar. Drugs 2021, 19, 686. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Albarano, L.; Esposito, R.; Zupo, V.; Costantini, M.; Ianora, A. Multiple Roles of Diatom-Derived Oxylipins within Marine Environments and Their Potential Biotechnological Applications. Mar. Drugs 2020, 18, 342. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Coppola, D. Biotechnological Applications of Products Released by Marine Microorganisms for Cold Adaptation Strategies: Polyunsaturated Fatty Acids, Antioxidants, and Antifreeze Proteins. J. Mar. Sci. Eng. 2023, 11, 1399. [Google Scholar] [CrossRef]
- Collins, T.; Margesin, R. Psychrophilic Lifestyles: Mechanisms of Adaptation and Biotechnological Tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Teoh, M.L.; Chu, W.L.; Marchant, H.; Phang, S.M. Influence of Culture Temperature on the Growth, Biochemical Composition and Fatty Acid Profiles of Six Antarctic Microalgae. J. Appl. Phycol. 2004, 16, 421–430. [Google Scholar] [CrossRef]
- Schulze, P.S.C.; Hulatt, C.J.; Morales-Sánchez, D.; Wijffels, R.H.; Kiron, V. Fatty Acids and Proteins from Marine Cold Adapted Microalgae for Biotechnology. Algal. Res. 2019, 42, 101604. [Google Scholar] [CrossRef]
- Li, H.Y.; Lu, Y.; Zheng, J.W.; Yang, W.D.; Liu, J.S. Biochemical and Genetic Engineering of Diatoms for Polyunsaturated Fatty Acid Biosynthesis. Mar. Drugs 2014, 12, 153–166. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Custódio, L.; Patarra, J.; Alberício, F.; Neng, N.d.R.; Nogueira, J.M.F.; Romano, A. Phenolic Composition, Antioxidant Potential and in Vitro Inhibitory Activity of Leaves and Acorns of Quercus Suber on Key Enzymes Relevant for Hyperglycemia and Alzheimer’s Disease. Ind. Crops Prod. 2015, 64, 45–51. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Megías, C.; Pastor-Cavada, E.; Torres-Fuentes, C.; Girón-Calle, J.; Alaiz, M.; Juan, R.; Pastor, J.; Vioque, J. Chelating, Antioxidant and Antiproliferative Activity of Vicia Sativa Polyphenol Extracts. Eur. Food Res. Technol. 2009, 230, 353–359. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Trentin, R.; Custódio, L.; Rodrigues, M.J.; Moschin, E.; Sciuto, K.; da Silva, J.P.; Moro, I. Exploring Ulva Australis Areschoug for Possible Biotechnological Applications: In Vitro Antioxidant and Enzymatic Inhibitory Properties, and Fatty Acids Contents. Algal. Res. 2020, 50, 101980. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. A Quantitative Starch–Iodine Method for Measuring Alpha-Amylase and Glucoamylase Activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Soszynski, A.; Martins, A.; Rauter, A.P.; Neng, N.R.; Nogueira, J.M.F.; Varela, J.; Barreira, L.; Custódio, L. Unravelling the Antioxidant Potential and the Phenolic Composition of Different Anatomical Organs of the Marine Halophyte Limonium Algarvense. Ind. Crops Prod. 2015, 77, 315–322. [Google Scholar] [CrossRef]
- McDougall, G.J.; Kulkarni, N.N.; Stewart, D. Berry Polyphenols Inhibit Pancreatic Lipase Activity in Vitro. Food Chem. 2009, 115, 193–199. [Google Scholar] [CrossRef]
- Zengin, G. A Study on in Vitro Enzyme Inhibitory Properties of Asphodeline Anatolica: New Sources of Natural Inhibitors for Public Health Problems. Ind. Crops Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Silva, S.G.; Paula, P.; da Silva, J.P.; Mil-Homens, D.; Teixeira, M.C.; Fialho, A.M.; Costa, R.; Keller-Costa, T. Insights into the Antimicrobial Activities and Metabolomes of Aquimarina (Flavobacteriaceae, Bacteroidetes) Species from the Rare Marine Biosphere. Mar. Drugs 2022, 20, 423. [Google Scholar] [CrossRef]
- Chambers, M.C.; MacLean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Schmid, R.; Heuckeroth, S.; Korf, A.; Smirnov, A.; Myers, O.; Dyrlund, T.S.; Bushuiev, R.; Murray, K.J.; Hoffmann, N.; Lu, M.; et al. Integrative Analysis of Multimodal Mass Spectrometry Data in MZmine 3. Nat. Biotechnol. 2023, 41, 447–449. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A Public Repository for Sharing Mass Spectral Data for Life Sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Microbial Metabolites through Database Search of Mass Spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A Rapid Tool for Turning Tandem Mass Spectra into Metabolite Structure Information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Böcker, S.; Letzel, M.C.; Lipták, Z.; Pervukhin, A. SIRIUS: Decomposing Isotope Patterns for Metabolite Identification. Bioinformatics 2009, 25, 218–224. [Google Scholar] [CrossRef]
- Ludwig, M.; Nothias, L.F.; Dührkop, K.; Koester, I.; Fleischauer, M.; Hoffmann, M.A.; Petras, D.; Vargas, F.; Morsy, M.; Aluwihare, L.; et al. Database-Independent Molecular Formula Annotation Using Gibbs Sampling through ZODIAC. Nat. Mach. Intell. 2020, 2, 629–641. [Google Scholar] [CrossRef]
- Hoffmann, M.A.; Nothias, L.-F.; Ludwig, M.; Fleischauer, M.; Gentry, E.C.; Witting, M.; Dorrestein, P.C.; Dührkop, K.; Böcker, S. Assigning Confidence to Structural Annotations from Mass Spectra with COSMIC. bioRxiv 2021. [Google Scholar] [CrossRef]
- Dührkop, K.; Nothias, L.F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C.; et al. Systematic Classification of Unknown Metabolites Using High-Resolution Fragmentation Mass Spectra. Nat. Biotechnol. 2020, 39, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated Chemical Classification with a Comprehensive, Computable Taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved Recovery of Fatty Acid through Direct Transesterification without Prior Extraction or Purification. J. Lipid. Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Fan, Y.; Parsons, R.L.; Hu, G.R.; Zhang, P.Y.; Li, F.L. A Rapid Method for the Determination of Fucoxanthin in Diatom. Mar. Drugs 2018, 16, 33. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Extract | ABTS | ICA |
|---|---|---|---|
| C. ineffabilis IMA082A | Acetone 80% | 6.79 ± 0.21 | 5.73 ± 0.53 b |
| Methanol 50% | – | 2.06 ± 0.70 a | |
| C. zucchelli IMA088A | Acetone 80% | – | – |
| Methanol 50% | – | – |
| Species | Extract | AChE | BChE | α-Amylase |
|---|---|---|---|---|
| C. ineffabilis IMA082A | Acetone 80% | 7.99 ± 2.62 a | 1.81 ± 0.46 a | 6.87 ± 0.17 |
| Methanol 50% | 6.03 ± 1.34 a | – | – | |
| C. zucchelli IMA088A | Acetone 80% | – | 4.34 ± 0.39 b | – |
| Methanol 50% | 9.70 ± 1.44 a | 6.59 ± 0.35 c | – |
| Peank No. | m/z Value | RT (min) | Adduct | Annotated Molecular Formula | Annotated Compound | IMA082A | IMA088A |
|---|---|---|---|---|---|---|---|
| 1 | 197.1172 | 13.3438 | [M + H]+ | C11H16O3 | Loliolide | 🗸 | 🗸 |
| 2 | 323.221 | 22.055 | [M + H]+ | C19H30O4 | MG(16:4) | 🗸 | 🗸 |
| 4 | 213.1632 | 22.3055 | [M − H4O2 + H]+ | C16H24O2 | 6,9,12,15-Hexadecatetraenoic acid | 🗸 | 🗸 |
| 5 | 327.252 | 22.4687 | [M − H4O2 + H]+ | C19H34O4 | MG(16:2) | 🗸 | 🗸 |
| 6 | 337.2362 | 22.4864 | [M − H4O2 + H]+ | C20H34O5 | Prostaglandin D1 | 🗸 | |
| 7 | 299.1996 | 22.6948 | [M − H4O2 + H]+ | C20H30O4 | Resolvin E3 | 🗸 | |
| 8 | 321.2416 | 22.8543 | [M − H4O2 + H]+ | C20H34O4 | 11,12-Dheta | 🗸 | 🗸 |
| 9 | 325.2362 | 22.9608 | [M + H]+ | C19H32O4 | MG(16:3) | 🗸 | 🗸 |
| 10 | 509.2702 | 22.9608 | [M + Na]+ | C25H42O9 | MGMG(16:3) | 🗸 | 🗸 |
| 11 | 303.231 | 23.0676 | [M + H]+ | C20H30O2 | Eicosapentaenoic acid | 🗸 | |
| 12 | 530.331 | 23.6646 | [M + H3N + H]+ | C27H44O9 | MGMG(18:4) | 🗸 | 🗸 |
| 13 | 351.2522 | 23.6919 | [M + H]+ | C21H34O4 | MG(18:4) | 🗸 | 🗸 |
| 14 | 516.307 | 23.8108 | [M + H]+ | C26H46NO7P | LPC(18:4) | 🗸 | 🗸 |
| 15 | 465.26 | 23.9435 | [M + H]+ | C22H43O9P | LPG(16:1) | 🗸 | |
| 16 | 542.3229 | 24.7478 | [M + H]+ | C28H48NO7P | LPC(20:5) | 🗸 | 🗸 |
| 17 | 508.3471 | 24.7865 | [M + H3N + H]+ | C25H46O9 | MGMG(16:1) | 🗸 | 🗸 |
| 18 | 518.323 | 24.9074 | [M + H]+ | C26H48NO7P | LPC(18:3) | 🗸 | 🗸 |
| 19 | 301.2157 | 25.0337 | [M − H4O2 + H]+ | C20H30O3 | 11-HEPE | 🗸 | 🗸 |
| 20 | 574.3244 | 25.2321 | [M + H3N + H]+ | C25H48O11S | SQMG(16:0) | 🗸 | 🗸 |
| 21 | 494.3219 | 25.4718 | [M + H]+ | C24H48NO7P | LPC(16:1) | 🗸 | 🗸 |
| 22 | 505.2522 | 25.6538 | [M + Na]+ | C22H43O9P | LPG(16:1) | 🗸 | 🗸 |
| 23 | 447.2491 | 25.6819 | [M − H4O2 + H]+ | C22H43O9P | PG(16:1) | 🗸 | 🗸 |
| 24 | 544.3382 | 25.7362 | [M + H]+ | C28H50NO7P | LPC(20:4) | 🗸 | 🗸 |
| 25 | 568.3384 | 25.789 | [M + H]+ | C30H50NO7P | LPC(22:6) | 🗸 | 🗸 |
| 26 | 520.3636 | 26.2199 | [M + H]+ | C26H50NO7P | LPC(18:2) | 🗸 | 🗸 |
| 27 | 449.2647 | 26.357 | [M − H4O2 + H]+ | C22H45O9P | PG(16:0) | 🗸 | 🗸 |
| 28 | 296.2577 | 26.6736 | [M + H3N + H]+ | C18H30O2 | alpha-Linolenic acid | 🗸 | 🗸 |
| 29 | 522.3537 | 28.4701 | [M + H]+ | C26H52NO7P | LPC(18:1) | 🗸 | 🗸 |
| 30 | 496.3379 | 28.6544 | [M + H]+ | C24H50NO7P | LPC(16:0) | 🗸 | 🗸 |
| 31 | 609.2693 | 30.5862 | [M + H]+ | C35H36N4O6 | 10-Hydroxyphaeophorbide | 🗸 | 🗸 |
| Peank No. | m/z Value | RT (min) | Adduct | Annotated Molecular Formula | Annotated Compound | IMA082A | IMA088A |
|---|---|---|---|---|---|---|---|
| 1 | 148.0598 | 0.9564 | [M + H]+ | C5H9NO4 | Glutamate | 🗸 | 🗸 |
| 2 | 132.1012 | 1.6404 | [M + H]+ | C6H13NO2 | Leucine | 🗸 | 🗸 |
| 3 | 197.1162 | 13.3722 | [M + H]+ | C11H16O3 | Loliolide | 🗸 | 🗸 |
| 4 | 507.2532 | 22.3166 | [M + Na]+ | C25H40O9 | MGMG(16:4) | 🗸 | |
| 5 | 274.2734 | 22.4747 | [M + H]+ | C16H35NO2 | Lauryldiethanolamine | 🗸 | 🗸 |
| 6 | 318.2998 | 22.6623 | [M + H]+ | C18H39NO3 | Phytosphingosine | 🗸 | 🗸 |
| 7 | 290.2684 | 22.8782 | [M + H]+ | C16H35NO3 | Hexadecaphytosphingosine | 🗸 | 🗸 |
| 8 | 530.3297 | 23.6846 | [M + H3N + H]+ | C27H44O9 | MGMG(18:4) | 🗸 | |
| 9 | 516.3079 | 23.7679 | [M + H]+ | C26H46NO7P | LPC(18:4) | 🗸 | 🗸 |
| Species | Extract | Fucoxanthin (mg/g DW) |
|---|---|---|
| C. ineffabilis IMA082A | Acetone 80% | 0.19 ± 0.04 ab |
| Methanol 50% | 0.25 ± 0.02 bc | |
| C. zucchelli IMA088A | Acetone 80% | 0.14 ± 0.03 a |
| Methanol 50% | 0.32 ± 0.05 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trentin, R.; Moschin, E.; Custódio, L.; Moro, I. Bioprospection of the Antarctic Diatoms Craspedostauros ineffabilis IMA082A and Craspedostauros zucchelli IMA088A. Mar. Drugs 2024, 22, 35. https://doi.org/10.3390/md22010035
Trentin R, Moschin E, Custódio L, Moro I. Bioprospection of the Antarctic Diatoms Craspedostauros ineffabilis IMA082A and Craspedostauros zucchelli IMA088A. Marine Drugs. 2024; 22(1):35. https://doi.org/10.3390/md22010035
Chicago/Turabian StyleTrentin, Riccardo, Emanuela Moschin, Luísa Custódio, and Isabella Moro. 2024. "Bioprospection of the Antarctic Diatoms Craspedostauros ineffabilis IMA082A and Craspedostauros zucchelli IMA088A" Marine Drugs 22, no. 1: 35. https://doi.org/10.3390/md22010035
APA StyleTrentin, R., Moschin, E., Custódio, L., & Moro, I. (2024). Bioprospection of the Antarctic Diatoms Craspedostauros ineffabilis IMA082A and Craspedostauros zucchelli IMA088A. Marine Drugs, 22(1), 35. https://doi.org/10.3390/md22010035