
Bioactivity Screening and Genomic Analysis Reveals Deep-Sea Fish Microbiome Isolates as Sources of Novel Antimicrobials

Abstract
1. Introduction
2. Results
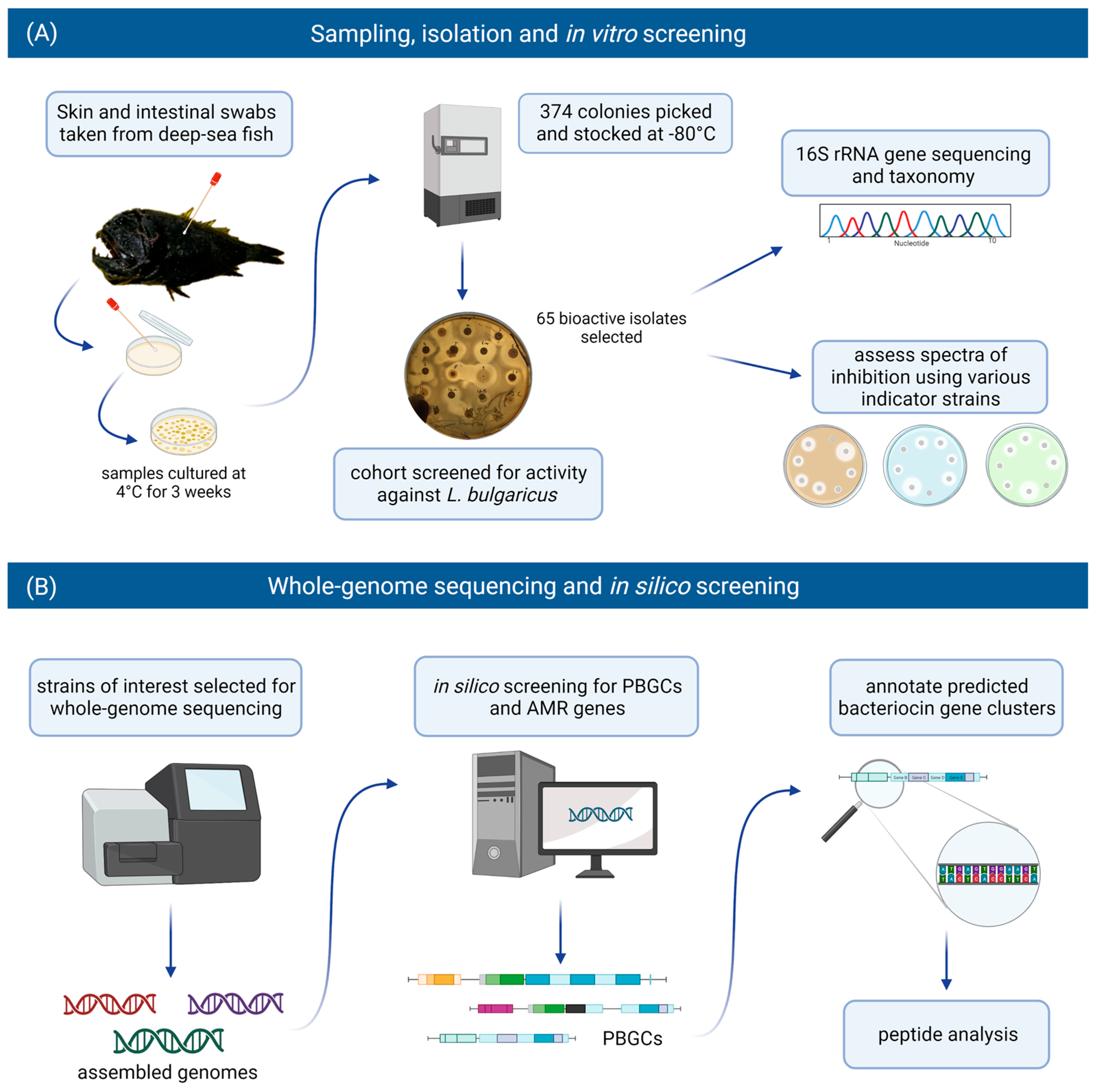
2.1. Recovery and Selection of Bioactive Isolates
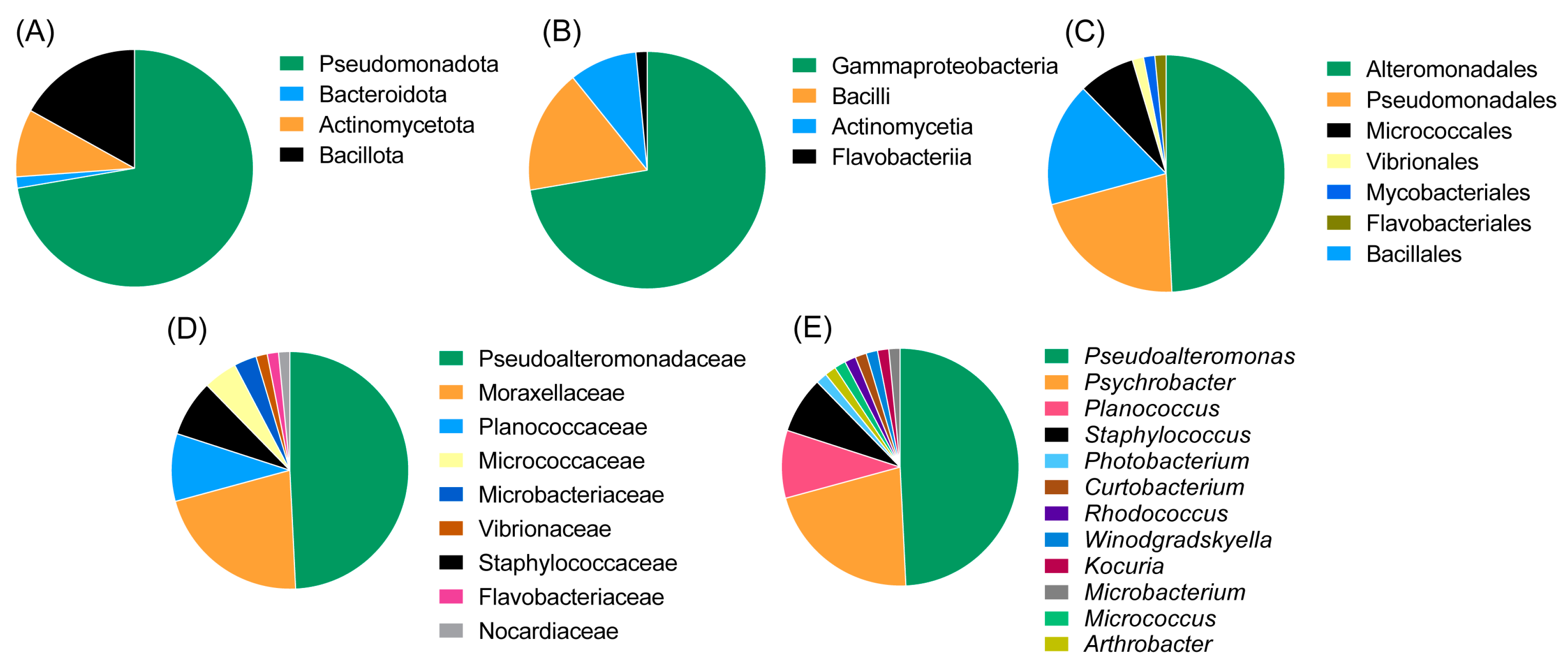
2.2. Taxonomic Diversity
2.3. Spectrum of Activity
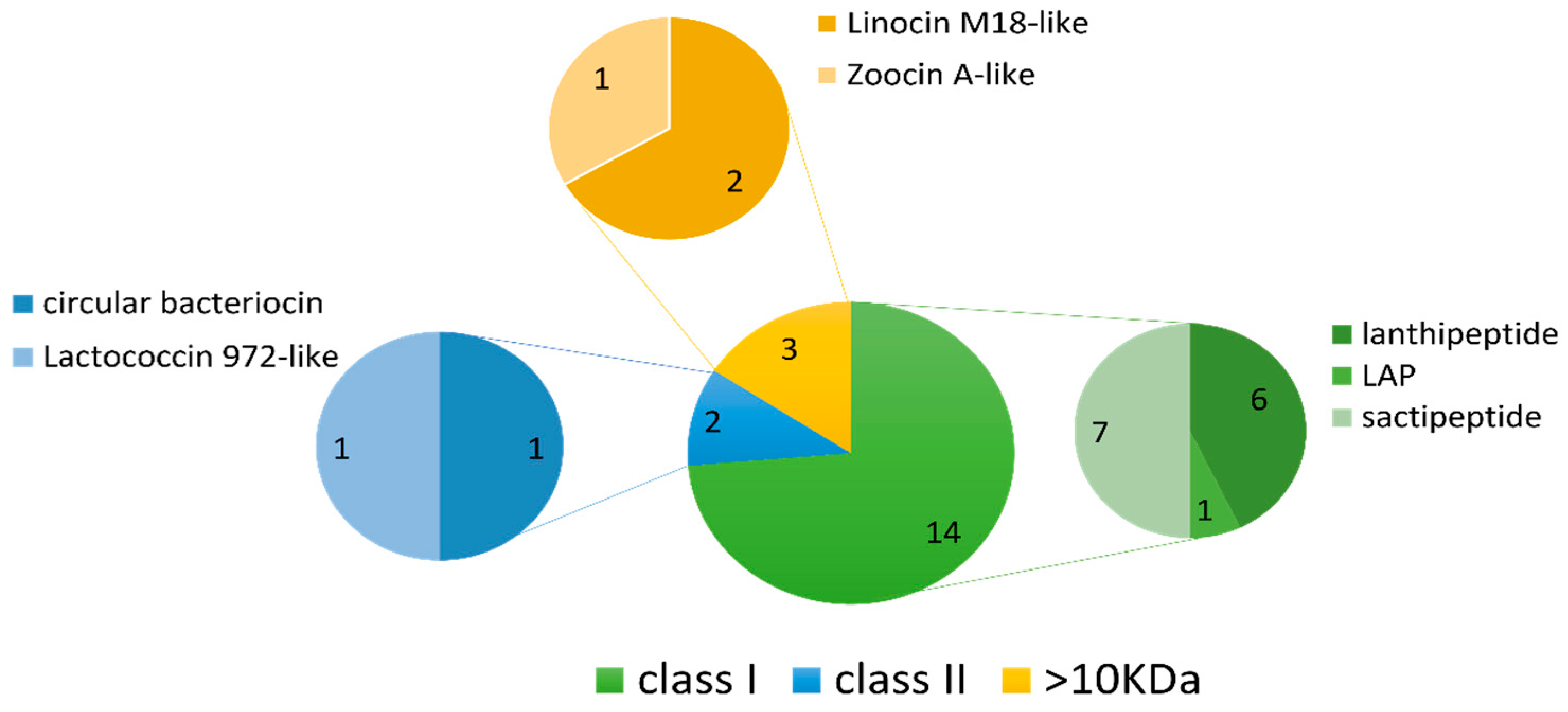
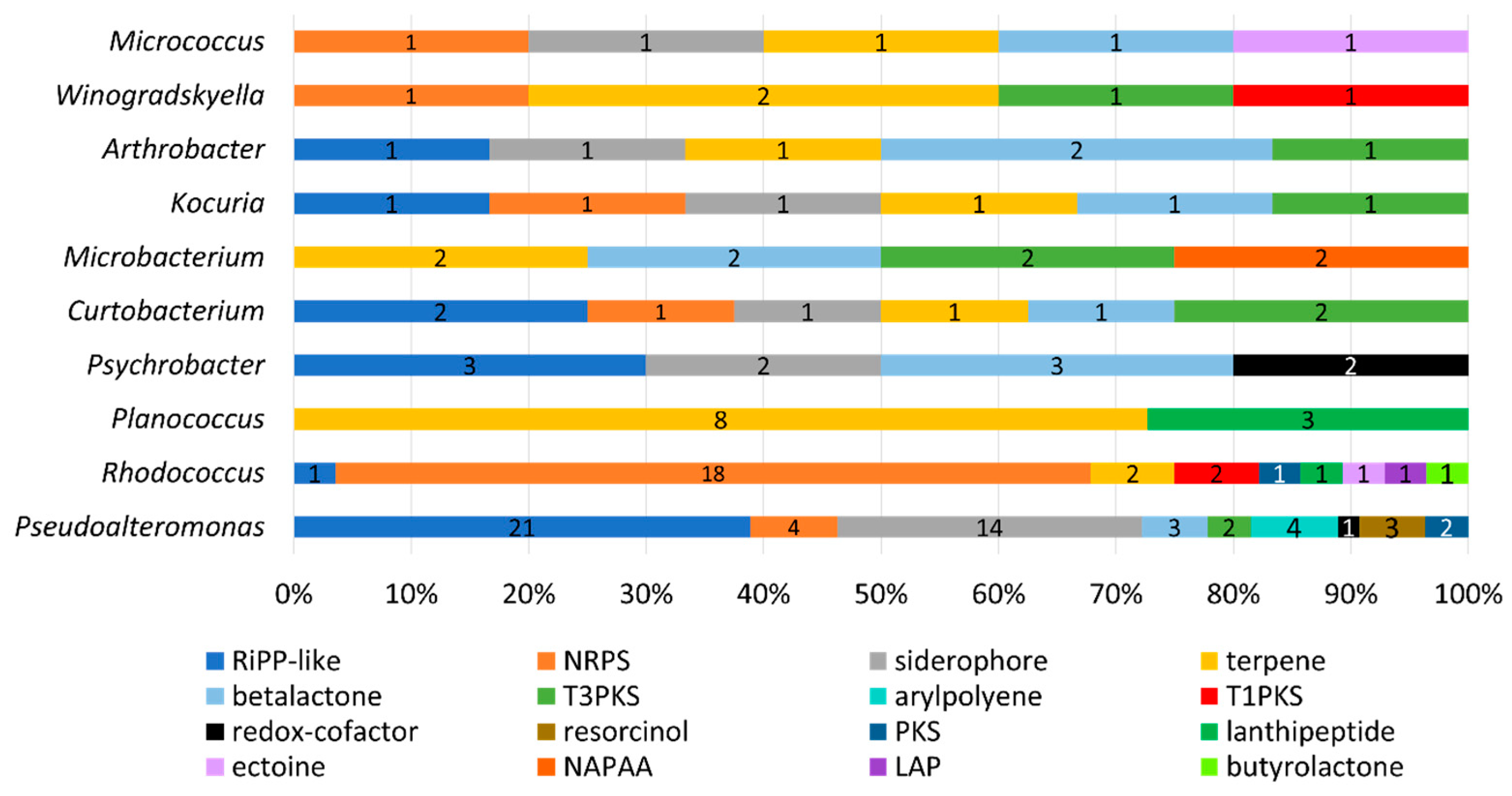
2.4. In Silico Antimicrobial Screening
2.4.1. Class I LanM-Group Bacteriocins (Lanthipeptides)
2.4.2. Class IIc (Circular) Bacteriocin
2.4.3. Class IId Bacteriocins
2.5. Antimicrobial Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Isolation and Cultivation of Antimicrobial-Producing Isolates
4.2. In Vitro Screening
4.3. 16S rRNA Gene Sequencing and Taxonomy
4.4. Genomic DNA Extraction and Sequencing
4.5. Genome Assembly and Annotation
4.6. In Silico Antimicrobial Screening
4.7. Putative Biosynthetic Gene Cluster (PBGC) Analysis
4.8. Antimicrobial Resistance Gene Screening
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Greenaway, S.F.; Sullivan, K.D.; Umfress, S.H.; Beittel, A.B.; Wagner, K.D. Revised depth of the Challenger Deep from submersible transects; including a general method for precise, pressure-derived depths in the ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2021, 178, 103644. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Snelgrove, P.V.R. The deep-sea under global change. Curr. Biol. 2017, 27, R461–R465. [Google Scholar] [CrossRef] [PubMed]
- Somero, G.N. Biochemical ecology of deep-sea animals. Experientia 1992, 48, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Pernice, M.C.; Giner, C.R.; Logares, R.; Perera-Bel, J.; Acinas, S.G.; Duarte, C.M.; Gasol, J.M.; Massana, R. Large variability of bathypelagic microbial eukaryotic communities across the world’s oceans. ISME J. 2016, 10, 945–958. (In English) [Google Scholar] [CrossRef]
- Zhang, J.-W.; Zeng, R.-Y. Molecular Cloning and Expression of a Cold-Adapted Lipase Gene from an Antarctic Deep Sea Psychrotrophic Bacterium Pseudomonas sp. 7323. Mar. Biotechnol. 2008, 10, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Liu, G.; Zheng, R.; Sun, C.; Wu, S. Structural and Functional Insights into Iturin W, a Novel Lipopeptide Produced by the Deep-Sea Bacterium Bacillus sp. Strain wsm-1. Appl. Environ. Microbiol. 2020, 86, e01597-20. [Google Scholar] [CrossRef]
- Penhallurick, R.W.; Ichiye, T. Pressure Adaptations in Deep-Sea Moritella Dihydrofolate Reductases: Compressibility versus Stability. Biology 2021, 10, 11. (In English) [Google Scholar] [CrossRef]
- Yan, B.-Q.; Chen, X.-L.; Hou, X.-Y.; He, H.; Zhou, B.-C.; Zhang, Y.-Z. Molecular analysis of the gene encoding a cold-adapted halophilic subtilase from deep-sea psychrotolerant bacterium Pseudoalteromonas sp. SM9913: Cloning, expression, characterization and function analysis of the C-terminal PPC domains. Extremophiles 2009, 13, 725–733. [Google Scholar] [CrossRef]
- Zhou, X.; Huang, H.; Chen, Y.; Tan, J.; Song, Y.; Zou, J.; Tian, X.; Hua, Y.; Ju, J. Marthiapeptide A, an Anti-infective and Cytotoxic Polythiazole Cyclopeptide from a 60 L Scale Fermentation of the Deep Sea-Derived Marinactinospora thermotolerans SCSIO 00652. J. Nat. Prod. 2012, 75, 2251–2255. [Google Scholar] [CrossRef]
- Niu, S.; Li, S.; Chen, Y.; Tian, X.; Zhang, H.; Zhang, G.; Zhang, W.; Yang, X.; Zhang, S.; Ju, J.; et al. Lobophorins E and F, new spirotetronate antibiotics from a South China Sea-derived Streptomyces sp. SCSIO 01127. J. Antibiot. 2011, 64, 711–716. [Google Scholar] [CrossRef]
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Osset, M.; Pérez-Victoria, I.; Martín, J.; De Pedro, N.; De la Cruz, M.; Díaz, C.; Vicente, F.; Reyes, F.; et al. Lobophorin K, a New Natural Product with Cytotoxic Activity Produced by Streptomyces sp. M-207 Associated with the Deep-Sea Coral Lophelia pertusa. Marine Drugs 2017, 15, 144. [Google Scholar] [CrossRef]
- Jang, K.H.; Nam, S.J.; Locke, J.B.; Kauffman, C.A.; Beatty, D.S.; Paul, L.A.; Fenical, W. Anthracimycin, a Potent Anthrax Antibiotic from a Marine-Derived Actinomycete. Angew. Chem. Int. Ed. 2013, 30, 7822–7824. [Google Scholar] [CrossRef]
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Pérez-Victoria, I.; Otero, L.; Fernández, J.; Palacios, J.J.; Martín, J.; de la Cruz, M.; Díaz, C.; Vicente, F.; et al. Branimycins B and C, Antibiotics Produced by the Abyssal Actinobacterium Pseudonocardia carboxydivorans M-227. J. Nat. Prod. 2017, 80, 569–573. [Google Scholar] [CrossRef]
- Back, C.R.; Stennett, H.L.; Williams, S.E.; Wang, L.; Ojeda Gomez, J.; Abdulle, O.M.; Duffy, T.; Neal, C.; Mantell, J.; Jepson, M.A.; et al. A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge. Mar. Drugs 2021, 19, 105. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Montalbán-López, M.; Scott, T.A.; Ramesh, S.; Rahman, I.R.; Van Heel, A.J.; Viel, J.H.; Bandarian, V.; Dittmann, E.; Genilloud, O.; Goto, Y.; et al. New developments in RiPP discovery, enzymology and engineering. Nat. Prod. Rep. 2021, 38, 130–239. [Google Scholar] [CrossRef] [PubMed]
- Nissen-Meyer, J.; Rogne, P.; Oppegard, C.; Haugen, S.H.; Kristiansen, E.P. Structure-Function Relationships of the Non-Lanthionine-Containing Peptide (class II) Bacteriocins Produced by Gram-Positive Bacteria. Curr. Pharm. Biotechnol. 2009, 10, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019 ed.; U.S. Department of Health and Human Services, CDC: Washington, DC, USA, 2019.
- Bindiya, E.S.; Tina, K.J.; Sasidharan, R.S.; Bhat, S.G. BaCf3: Highly thermostable bacteriocin from Bacillus amyloliquefaciens BTSS3 antagonistic on food-borne pathogens. 3 Biotech 2019, 9, 136. [Google Scholar] [CrossRef]
- Bindiya, E.S.; Tina, K.J.; Raghul, S.S.; Bhat, S.G. Characterization of Deep Sea Fish Gut Bacteria with Antagonistic Potential, from Centroscyllium fabricii (Deep Sea Shark). Probiotics Antimicrob. Proteins 2015, 7, 157–163. (In English) [Google Scholar] [CrossRef] [PubMed]
- Collins, F.W.; Walsh, C.J.; Gomez-Sala, B.; Guijarro-García, E.; Stokes, D.; Jakobsdóttir, K.B.; Kristjánsson, K.; Burns, F.; Cotter, P.D.; Rea, M.C.; et al. The microbiome of deep-sea fish reveals new microbial species and a sparsity of antibiotic resistance genes. Gut Microbes 2021, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.N.; O’Connor, P.M.; Rea, M.C.; O’Sullivan, O.; Walsh, C.J.; Healy, B.; Mathur, H.; Field, D.; Hill, C.; Ross, R.P. Nisin J, a Novel Natural Nisin Variant, Is Produced by Staphylococcus capitis Sourced from the Human Skin Microbiota. J. Bacteriol. 2020, 202, e00639-19. [Google Scholar] [CrossRef] [PubMed]
- Sugrue, I.; O’Connor, P.M.; Hill, C.; Stanton, C.; Ross, R.P. Actinomyces Produces Defensin-Like Bacteriocins (Actifensins) with a Highly Degenerate Structure and Broad Antimicrobial Activity. J. Bacteriol. 2020, 202, e00529-19. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, E.F.; Gardiner, G.E.; O’Connor, P.M.; Mills, S.; Ross, R.P.; Hill, C. Characterization of enterocin- and salivaricin-producing lactic acid bacteria from the mammalian gastrointestinal tract. FEMS Microbiol. Lett. 2009, 291, 24–34. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Teng, K.; Sun, S.; Sun, Z.; Zhong, J. Cerecidins, novel lantibiotics from Bacillus cereus with potent antimicrobial activity. Appl. Environ. Microbiol. 2014, 80, 2633–2643. (In English) [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- Balo, A.R.; Tao, L.; Britt, R.D. Characterizing SPASM/twitch Domain-Containing Radical SAM Enzymes by EPR Spectroscopy. Appl. Magn. Reson. 2022, 53, 809–820. [Google Scholar] [CrossRef]
- Grell, T.A.; Kincannon, W.M.; Bruender, N.A.; Blaesi, E.J.; Krebs, C.; Bandarian, V.; Drennan, C.L. Structural and spectroscopic analyses of the sporulation killing factor biosynthetic enzyme SkfB, a bacterial AdoMet radical sactisynthase. J. Biol. Chem. 2018, 293, 17349–17361. [Google Scholar] [CrossRef]
- Booth, M.C.; Bogie, C.P.; Sahl, H.-G.; Siezen, R.J.; Hatter, K.L.; Gilmore, M.S. Structural analysis and proteolytic activation of Enterococcus faecalis cytolysin, a novel lantibiotic. Mol. Microbiol. 1996, 21, 1175–1184. [Google Scholar] [CrossRef]
- Lohans, C.T.; Li, J.L.; Vederas, J.C. Structure and Biosynthesis of Carnolysin, a Homologue of Enterococcal Cytolysin with d-Amino Acids. J. Am. Chem. Soc. 2014, 136, 13150–13153. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Bactericidal and bacteriolytic action of peptide antibiotic AS-48 against gram-positive and gram-negative bacteria and other organisms. Res. Microbiol. 1989, 140, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, F.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Valdivia, E. Antilisterial activity of peptide AS-48 and study of changes induced in the cell envelope properties of an AS-48-adapted strain of Listeria monocytogenes. Appl. Environ. Microbiol. 1999, 65, 618–625. (In English) [Google Scholar] [CrossRef]
- Grande Burgos, M.J.; Pulido, R.P.; Del Carmen López Aguayo, M.; Gálvez, A.; Lucas, R. The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications. Int. J. Mol. Sci. 2014, 15, 22706–22727. [Google Scholar] [CrossRef]
- Mu, F.; Masuda, Y.; Zendo, T.; Ono, H.; Kitagawa, H.; Ito, H.; Nakayama, J.; Sonomoto, K. Biological function of a DUF95 superfamily protein involved in the biosynthesis of a circular bacteriocin, leucocyclicin Q. J. Biosci. Bioeng. 2014, 117, 158–164. [Google Scholar] [CrossRef]
- Cebrián, R.; Maqueda, M.; Neira José, L.; Valdivia, E.; Martínez-Bueno, M.; Montalbán-López, M. Insights into the Functionality of the Putative Residues Involved in Enterocin AS-48 Maturation. Appl. Environ. Microbiol. 2010, 76, 7268–7276. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cebrián, R.; Martínez-Bueno, M.; Valdivia, E.; Albert, A.; Maqueda, M.; Sánchez-Barrena, M.J. The bacteriocin AS-48 requires dimer dissociation followed by hydrophobic interactions with the membrane for antibacterial activity. J. Struct. Biol. 2015, 190, 162–172. [Google Scholar] [CrossRef]
- Martínez, B.; Suárez, J.E.; Rodríguez, A. Lactococcin 972: A homodimeric lactococcal bacteriocin whose primary target is not the plasma membrane. Microbiology 1996, 142, 2393–2398. (In English) [Google Scholar] [CrossRef]
- Chikindas, M.L.; Novák, J.; Driessen, A.J.; Konings, W.N.; Schilling, K.M.; Caufield, P.W. Mutacin II, a bactericidal antibiotic from Streptococcus mutans. Antimicrob. Agents Chemother. 1995, 39, 2656–2660. (In English) [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Teng, K.; Wang, J.; Zhang, Z.; Zhang, J.; Sun, S.; Li, L.; Yang, X.; Zhong, J. CerR, a Single-Domain Regulatory Protein of the LuxR Family, Promotes Cerecidin Production and Immunity in Bacillus cereus. Appl. Environ. Microbiol. 2018, 84, e02245-17. [Google Scholar] [CrossRef]
- Ishikawa, J.; Chiba, K.; Kurita, H.; Satoh, H. Contribution of rpoB2 RNA polymerase beta subunit gene to rifampin resistance in Nocardia species. Antimicrob. Agents Chemother. 2006, 50, 1342–1346. (In English) [Google Scholar] [CrossRef] [PubMed]
- Talà, A.; Wang, G.; Zemanova, M.; Okamoto, S.; Ochi, K.; Alifano, P. Activation of Dormant Bacterial Genes by Nonomuraea sp. Strain ATCC 39727 Mutant-Type RNA Polymerase. J. Bacteriol. 2009, 191, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Yazawa, K.; Dabbs, E.R.; Nishikawa, K.; Komaki, H.; Mikami, Y.; Miyaji, M.; Morisaki, N.; Iwasaki, S. Different Rifampicin Inactivation Mechanisms in Nocardia and Related Taxa. Microbiol. Immunol. 1996, 40, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.P. Resistance to rifampicin: A review. J. Antibiot. 2014, 67, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, Z.; Wang, Y.; Liao, M.; Rong, X.; Li, B.; Wang, K.; Chen, J.; Zhang, H. Complete Genome Sequence of Photobacterium damselae Subsp. damselae Strain SSPD1601 Isolated from Deep-Sea Cage-Cultured Sebastes schlegelii with Septic Skin Ulcer. Int. J. Genom. 2019, 4242653, 2019. [Google Scholar] [CrossRef]
- Chen, H.; Bai, X.; Jing, L.; Chen, R.; Teng, Y. Characterization of antibiotic resistance genes in the sediments of an urban river revealed by comparative metagenomics analysis. Sci. Total Environ. 2019, 653, 1513–1521. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Liu, P.; Sun, Y.; Dong, X.; Hu, X. Unveiling the occurrence, hosts and mobility potential of antibiotic resistance genes in the deep ocean. Sci. Total Environ. 2022, 816, 151539. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The Ocean as a Global Reservoir of Antibiotic Resistance Genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. (In English) [Google Scholar]
- Rather, I.A.; Galope, R.; Bajpai, V.K.; Lim, J.; Paek, W.K.; Park, Y.H. Diversity of Marine Bacteria and Their Bacteriocins: Applications in Aquaculture. Rev. Fish. Sci. Aquac. 2017, 25, 257–269. (In English) [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. (In English) [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. (In English) [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. (In English) [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res 2016, 44, 6614–6624. (In English) [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Agrawal, P.; Khater, S.; Gupta, M.; Sain, N.; Mohanty, D. RiPPMiner: A bioinformatics resource for deciphering chemical structures of RiPPs based on prediction of cleavage and cross-links. Nucleic Acids Res 2017, 45, W80–W88. (In English) [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. (In English) [Google Scholar] [CrossRef]
- Harrison, K.J.; Crécy-Lagard, V.; Zallot, R. Gene Graphics: A genomic neighborhood data visualization web application. Bioinformatics 2018, 34, 1406–1408. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. (In English) [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res 2017, 45, D566–D573. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ingle, D.J.; Valcanis, M.; Kuzevski, A.; Tauschek, M.; Inouye, M.; Stinear, T.; Levine, M.M.; Robins-Browne, R.M.; Holt, K.E. In silico serotyping of E. coli from short read data identifies limited novel O-loci but extensive diversity of O:H serotype combinations within and between pathogenic lineages. Microb. Genom. 2016, 2, e000064. (In English) [Google Scholar] [CrossRef] [PubMed]
- Doster, E.; Lakin, S.M.; Dean, C.J.; Wolfe, C.; Young, J.G.; Boucher, C.; Belk, K.E.; Noyes, N.R.; Morley, P.S. MEGARes 2.0: A database for classification of antimicrobial drug, biocide and metal resistance determinants in metagenomic sequence data. Nucleic Acids Res. 2020, 48, D561–D569. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, 11. (In English) [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother 2014, 58, 3895–3903. (In English) [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. (In English) [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res 2016, 44, D694–D697. (In English) [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity vs. L. bulgaricus LMG 6901 | ||||
|---|---|---|---|---|
| Isolate | MA | BHI | mTSA | Source (Sample, Host) |
| Arthrobacter sp. APC 3897 | − | + | + | skin, 6 |
| Curtobacterium sp. APC 4022 | − | − | + | intestine, 4 |
| Photobacterium sp. APC 3280 | − | + | + | intestine, 4 |
| Planococcus sp. 26D.a_F | − | nd | +++ | skin, 6 |
| Pseudoalteromonas spp. | ||||
| APC 3495 | + | + | + | intestine, 1 |
| APC 3213 | + | + | ++ | intestine, 2 |
| APC 3238 | + | + | + | intestine, 3 |
| APC 3419 | + | ng | + | intestine, 3 |
| APC 3695 | − | + | +++ | intestine, 3 |
| APC 4023 | + | + | + | intestine, 3 |
| APC 4024 | + | + | +++ | intestine, 3 |
| APC 3284 | + | ng | ng | intestine, 4 |
| APC 3355 | ++ | +++ | ng | intestine, 4 |
| APC 3356 | ++ | nd | − | intestine, 4 |
| APC 3358 | ++ | +++ | ++ | intestine, 4 |
| APC 3412 | ++ | ng | ng | intestine, 5 |
| APC 3224 | + | ng | ng | intestine, 6 |
| APC 3391 | + | ++ | + | skin, 7 |
| APC 3502 | + | ng | + | skin, 1 |
| APC 4025 | + | + | + | skin, 2 |
| APC 4026 | + | + | + | skin, 2 |
| APC 3893 | ++ | ng | ng | skin, 3 |
| APC 3895 | + | ng | − | skin, 3 |
| APC 3896 | − | +++ | + | skin, 3 |
| APC 4017 | + | ng | ng | skin, 3 |
| APC 3274 | − | +++ | + | skin, 4 |
| APC 3691 | + | ng | ng | skin, 4 |
| APC 3407 | + | + | + | skin, 5 |
| APC 3904 | − | + | + | skin, 5 |
| APC 3221 | ++ | ng | + | skin, 6 |
| APC 3250 | + | + | + | skin, 6 |
| APC 4019 | + | nd | + | skin, 6 |
| APC 4020 | + | nd | ++ | skin, 6 |
| Psychrobacter spp. | ||||
| 5A.1 | − | ++ | + | skin, 4 |
| 5A.2 | − | +++ | + | skin, 4 |
| APC 3272 | − | ++ | − | skin, 6 |
| APC 3275 | − | + | + | intestine, 6 |
| APC 3276 | − | + | + | skin, 4 |
| APC 3277 | + | + | + | skin, 4 |
| APC 3350 | + | ++ | − | skin, 4 |
| APC 3426 | − | ++ | +++ | skin, 3 |
| APC 3692 | ++ | nd | + | skin, 6 |
| APC 4028 | − | ++ | + | skin, 4 |
| Strain | BAGEL4 | ||
|---|---|---|---|
| Prediction | Annotation | % Sim. | |
| Arthrobacter sp. APC 3897 | circularin_A | bacteriocin class cyclical uberolysin-like | 67% |
| sactipeptide | GTP 3’,8-cyclase (moaA) | 73.70% | |
| Curtobacterium sp. APC 4022 | lactococcin_972 | bacteriocin (Lactococcin_972) | 39.70% |
| sactipeptide | GTP 3’,8-cyclase (moaA) | 59.50% | |
| Kocuria sp. APC 4018 | linocin-M18 | encapsulating protein for peroxidase | 58.90% |
| Planococcus sp. APC 3900 | bacteriocin mersacidin/cerecidin (×2) | bacteriocin class II with double-glycine leader (×2) | 50–55.6% |
| Planococcus sp. APC 4015 | bacteriocin mersacidin/cerecidin (×2) | bacteriocin class II with double-glycine leader peptide (×2) | 50–55.6% |
| sactipeptide | undefined | - | |
| Planococcus sp. APC 4016 | bacteriocin mersacidin/cerecidin (×2) | bacteriocin class II with double-glycine leader (×2) | 50–55.6% |
| Pseudoalteromonas sp. APC 3224 | zoocin_A | undefined | 36.40% |
| Pseudoalteromonas sp. APC 3426 | sactipeptide | undefined | - |
| Pseudoalteromonas sp. APC 3893 | lanthipeptide class III | undefined | - |
| Pseudoalteromonas sp. APC 4017 | lanthipeptide class III | undefined | - |
| Psychrobacter sp. APC 3350 | sactipeptide | probable GTP 3’,8-cyclase | - |
| Rhodococcus sp. APC 3903 | lanthipeptide class III | undefined | - |
| LAPs | undefined | - | |
| linocin_M18/putative bacteriocin family protein | encapsulating protein for peroxidase | 66.70% | |
| sactipeptide | undefined | - | |
| Winogradskyella sp. APC 3343 | sactipeptide | GTP 3’,8-cyclase (moaA) | 41.40% |
| Strain | AntiSMASH | |||
|---|---|---|---|---|
| Bacteriocin Prediction | Similar Cluster | % Sim. | Other PBGCs Present | |
| Arthrobacter sp. APC 3897 | RiPP-like | head-to-tail cyclized peptide | 71 | betalactone (×2), siderophore, T3PKS, terpene |
| Curtobacterium sp. APC 4022 | RiPP-like (×2) | lactococcin_972 (×2) | nd | betalactone, NRPS-like, siderophore, T3PKS (×2), terpene |
| Kocuria sp. APC 4018 | RiPP-like | linocin_M18 | nd | betalactone, NRPS-like, siderophore, T3PKS, terpene |
| Microbacterium sp. APC 3898 | none | - | - | betalactone, NAPAA, T3PKS, terpene |
| Microbacterium sp. APC 3901 | none | - | - | NAPPA, T3PKS, betalactone, terpene |
| Micrococcus sp. APC 4021 | none | - | - | betalactone, ectoine, NRPS-like, siderophore, terpene |
| Planococcus sp. APC 3900 | lanthipeptide class II | cerecidin | 70 | terpene (×2) |
| Planococcus sp. APC 3906 | none | - | - | terpene (×2) |
| Planococcus sp. APC 4015 | lanthipeptide | cerecidin | 70 | terpene (×2) |
| Planococcus sp. APC 4016 | lanthipeptide class II | cerecidin | 70 | terpene (×2) |
| Pseudoalteromonas sp. APC 3213 | RiPP-like | burkholderic acid | 15 | siderophore |
| Pseudoalteromonas sp. APC 3215 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3218 | RiPP-like | none (DUF692-family protein) | nd | siderophore |
| Pseudoalteromonas sp. APC 3224 | RiPP-like (×2) | none (DUF692-family protein, ×2) | nd | NRPS, siderophore |
| Pseudoalteromonas sp. APC 3227 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3250 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3350 | RiPP-like | none (DUF692-family protein) | nd | betalactone, redox-cofactor, siderophore |
| Pseudoalteromonas sp. APC 3356 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3358 | RiPP-like | nucleocidin | 17 | arylpolyene |
| Pseudoalteromonas sp. APC 3426 | none | - | - | betalactone |
| Pseudoalteromonas sp. APC 3495 | RiPP-like | none (DUF692-family protein) | nd | siderophore |
| Pseudoalteromonas sp. APC 3691 | RiPP-like | none (DUF692-family protein) | nd | arylpolyene, betalactone |
| Pseudoalteromonas sp. APC 3694 | RiPP-like | none (DUF692-family protein) | nd | none |
| Pseudoalteromonas sp. APC 3893 | RiPP-like | none | - | arylpolyene, resorincol, NRPS (×2), PKS, T3PKS |
| Pseudoalteromonas sp. APC 3894 | RiPP-like | none (DUF692-family protein) | nd | siderophore |
| Pseudoalteromonas sp. APC 3895 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3896 | RiPP-like | none | - | siderophore |
| Pseudoalteromonas sp. APC 3907 | RiPP-like | none | - | siderophore, betalactone |
| Pseudoalteromonas sp. APC 4017 | RiPP-like | none | - | arylpolyene, resorcinol (×2), PKS, NRPS, T3PKS |
| Pseudoalteromonas sp. APC 4026 | RiPP-like | none (DUF692-family protein) | nd | siderophore |
| Psychrobacter sp. 5A.1 | RiPP-like | none (DUF692-family protein) | nd | betalactone, redox-cofactor, siderophore |
| Psychrobacter sp. APC 3279 | none | - | - | betalactone |
| Psychrobacter sp. APC 3281 | none | - | - | betalactone, redox-cofactor |
| Psychrobacter sp. APC 3355 | RiPP-like (×2) | none (DUF692-family protein, ×2) | nd | siderophore |
| Rhodococcus sp. APC 3903 | LAP | dissonitrile antibiotic SF2768 | 11 | butyrolactone, ectoine, NRPS (×15), NRPS-like (×3), terpene (×2), PKS-like, T1PKS (×2), |
| RiPP-like | branched-chain fatty acids | 75 | ||
| lanthipeptide class III | none | - | ||
| Winogradskyella sp. APC 3343 | none | - | - | NRPS, T1pks, T3PKS, Terpene (×2) |
| Contig | Gene | Resistance | %ID | Database | Product | Accession |
|---|---|---|---|---|---|---|
| 6 | rpoB2 | rifamycin/rifampin | 80 | card | beta-subunit of RNA polymerase (RpoB2) | AP006618.1:4835199-4838688 |
| 8 | (rif)iri | rifamycin/rifampin | 97.81 | argannot | (Rif)iri | U56415:280-1719 |
| 8 | iri | rifamycin/rifampin | 97.81 | ncbi | rifampin monooxygenase Iri | NG_047911.1 |
| 8 | iri | rifamycin/rifampin | 97.81 | card | rifampin monooxygenase Iri | U56415:279-1719 |
| 8 | iri | rifamycin/rifampin | 97.81 | megares | rifampin monooxygenase Iri | MEG_3434 |
| 9 | mtrA | macrolide;penam | 80.41 | card | transcriptional activator of multidrug efflux pump (MtrCDE) | AL123456.3:3627349-3626662 |
| 9 | mtrAD | multidrug | 80.41 | megares | Multi-drug RND efflux regulator MTRAD | MEG_4078 |
| Species | Strain ID | Temperature | Atmosphere | Growth Media | Notes |
|---|---|---|---|---|---|
| Arthrobacter sp. | APC 3897 | 20 °C | aerobic | MA | |
| Bacillus cereus | DPC 6087 | 37 °C | aerobic | BHI | |
| Enterococcus faecalis | OG1RF | 37 °C | aerobic | BHI | |
| Enterococcus faecium | DSM 25644 | 37 °C | aerobic | BHI | |
| Escherichia coli | MG1655 | 37 °C | aerobic | LB | |
| Kocuria sp. | APC 4018 | 20 °C | aerobic | MA | |
| Lactobacillus acidophilus | EM066-BC-T3-3 | 37 °C | anaerobic | MRS | +0.5 g/L cysteine |
| Lactobacillus delbrueckii subsp. bulgaricus | LMG 6901 | 37 °C | aerobic | MRS | +0.5 g/L cysteine |
| Lactococcus lactis subsp. cremoris | HP | 37 °C | aerobic | GM17 | |
| Listeria innocua | ATCC 33090 | 30 °C | aerobic | BHI | |
| Listeria monocytogenes | EDG-e | 37 °C | aerobic | BHI | |
| Microbacterium sp. | APC 3901 | 20 °C | aerobic | MA | |
| Micrococcus luteus | DSM 1790 | 30 °C | aerobic | BHI | |
| Planococcus sp. | APC 3906 | 20 °C | aerobic | MA | |
| Pseudomonas aeruginosa | PA01 | 37 °C | aerobic | BHI | |
| Psychrobacter sp. | APC 3276 | 20 °C | aerobic | MA | |
| Rhodococcus sp. | APC 3903 | 20 °C | aerobic | MA | |
| Salmonella enterica ser. Typhimurium | DPC 6046 | 37 °C | aerobic | BHI | |
| Staphylococcus aureus | RN4220 | 37 °C | aerobic | BHI | |
| Staphylococcus intermedius | DSM 20373 | 37 °C | aerobic | BHI | |
| Streptococcus pyogenes | DPC 6992 | 37 °C | aerobic | BHI | |
| Vibrio fischeri | n/a | 20 °C | aerobic | MA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uniacke-Lowe, S.; Collins, F.W.J.; Hill, C.; Ross, R.P. Bioactivity Screening and Genomic Analysis Reveals Deep-Sea Fish Microbiome Isolates as Sources of Novel Antimicrobials. Mar. Drugs 2023, 21, 444. https://doi.org/10.3390/md21080444
Uniacke-Lowe S, Collins FWJ, Hill C, Ross RP. Bioactivity Screening and Genomic Analysis Reveals Deep-Sea Fish Microbiome Isolates as Sources of Novel Antimicrobials. Marine Drugs. 2023; 21(8):444. https://doi.org/10.3390/md21080444
Chicago/Turabian StyleUniacke-Lowe, Shona, Fergus W. J. Collins, Colin Hill, and R. Paul Ross. 2023. "Bioactivity Screening and Genomic Analysis Reveals Deep-Sea Fish Microbiome Isolates as Sources of Novel Antimicrobials" Marine Drugs 21, no. 8: 444. https://doi.org/10.3390/md21080444
APA StyleUniacke-Lowe, S., Collins, F. W. J., Hill, C., & Ross, R. P. (2023). Bioactivity Screening and Genomic Analysis Reveals Deep-Sea Fish Microbiome Isolates as Sources of Novel Antimicrobials. Marine Drugs, 21(8), 444. https://doi.org/10.3390/md21080444