
A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys molitrix)

Abstract
1. Introduction
2. Results
2.1. Extraction Yields of Collagens
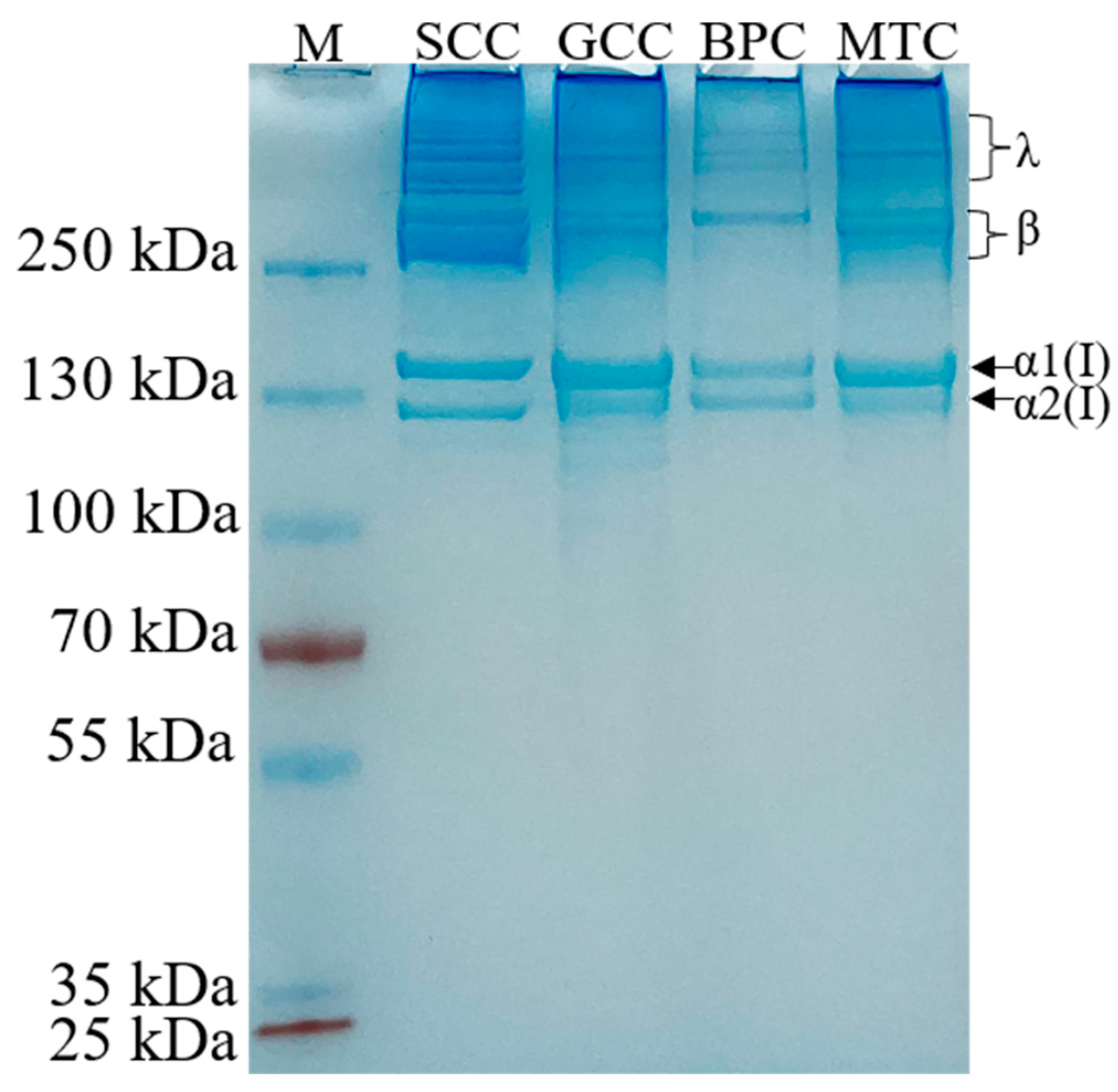
2.2. SDS-PAGE
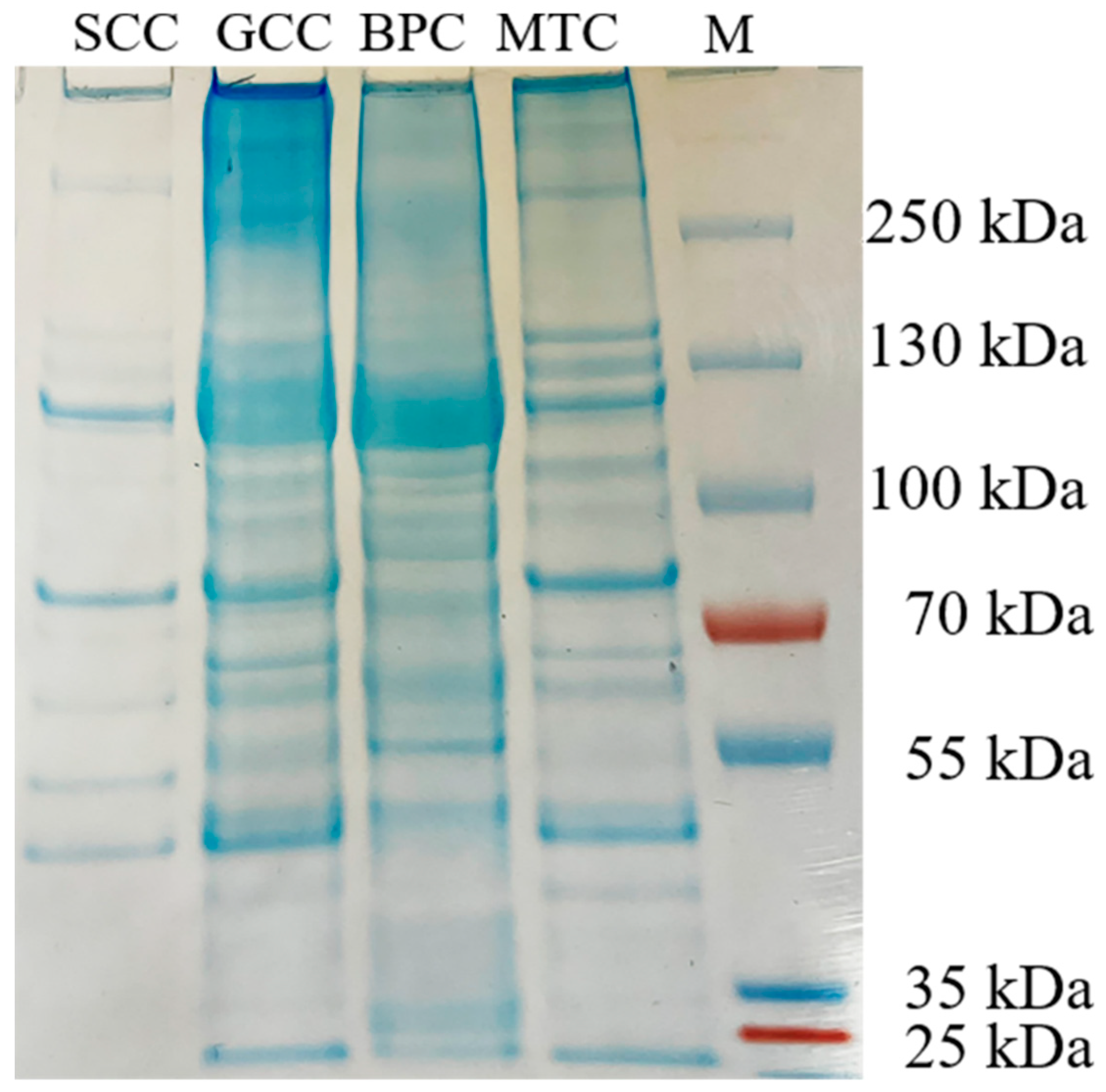
2.3. Peptide Maps
2.4. Amino Acid Composition
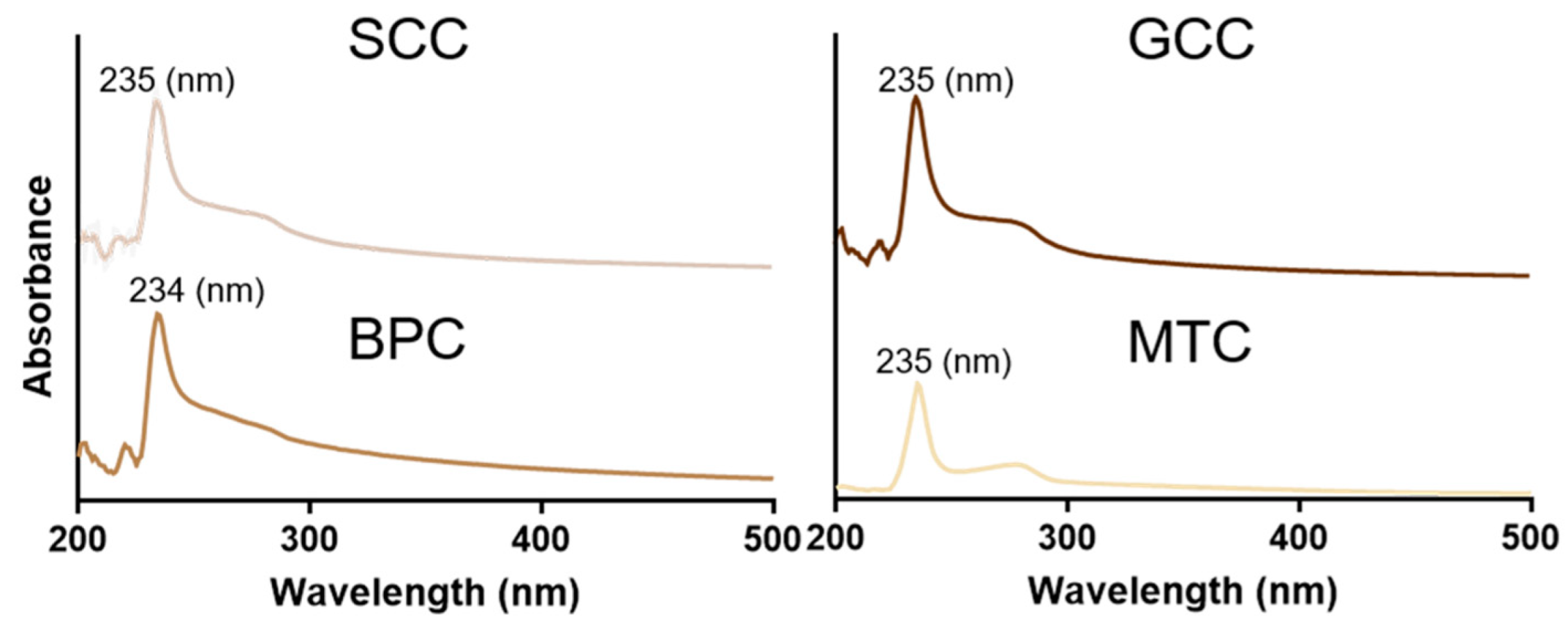
2.5. UV Absorption Spectrum
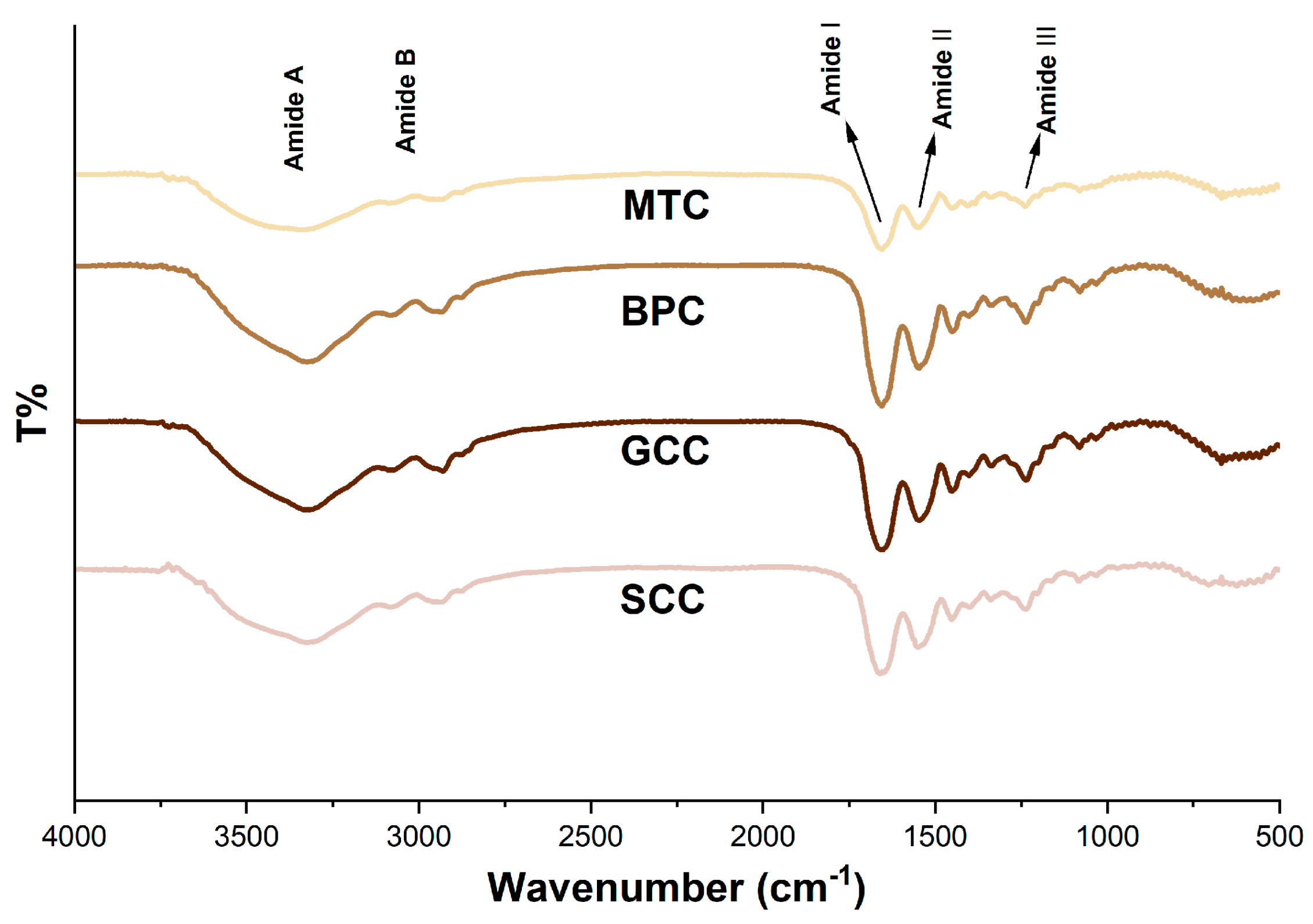
2.6. FTIR Spectroscopy
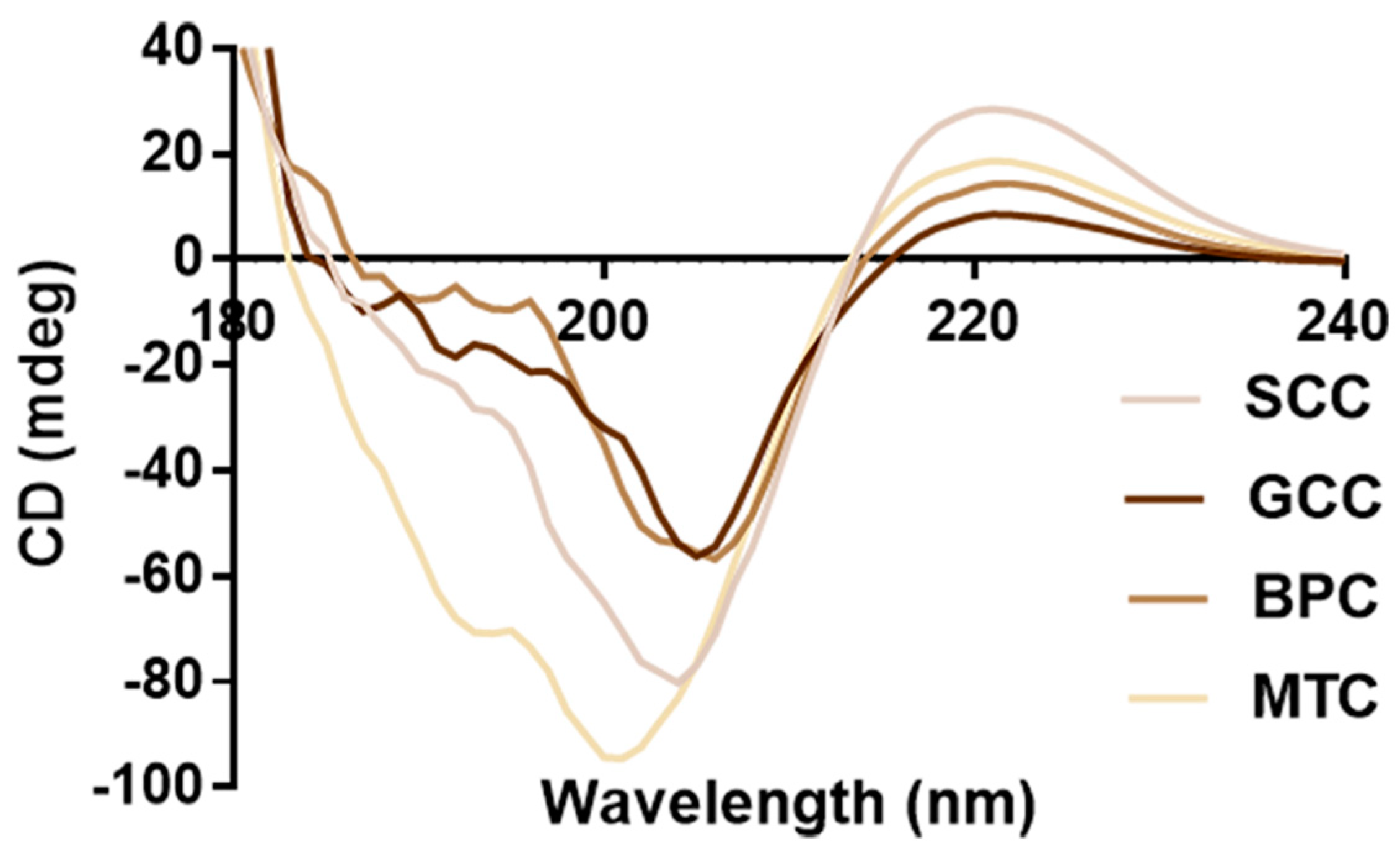
2.7. CD Spectrum
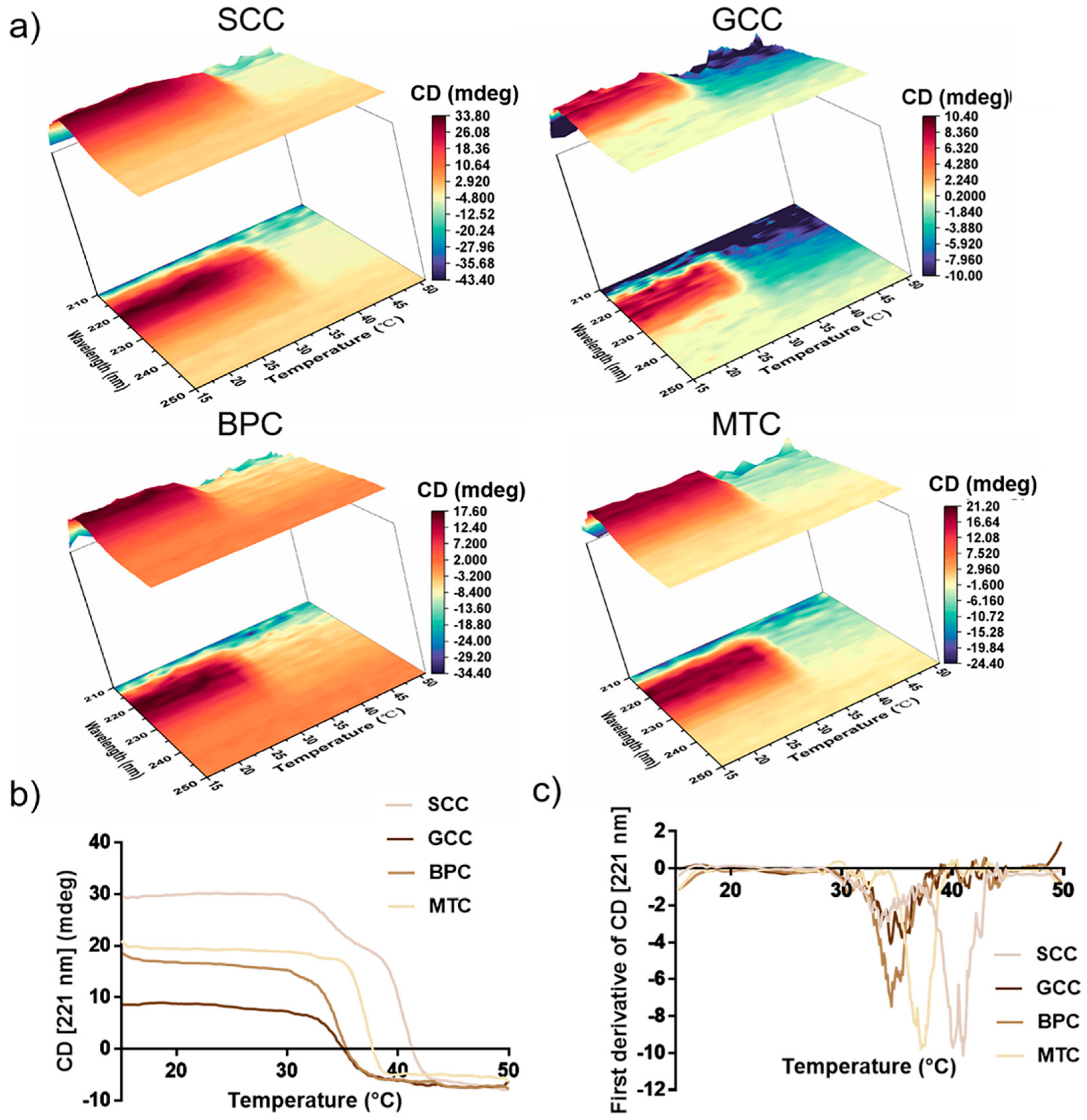
2.8. Thermal Stability
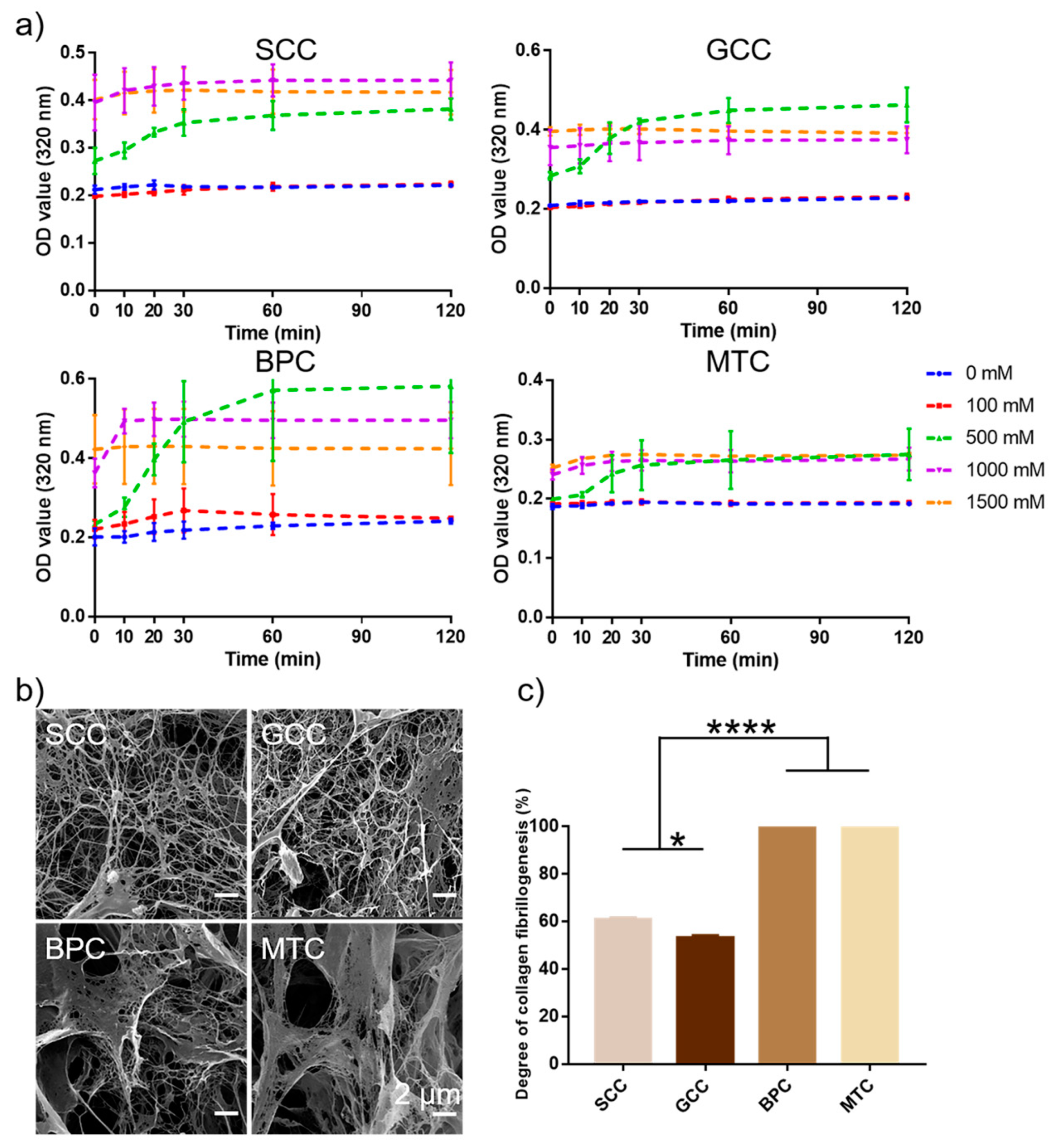
2.9. Collagen Fibrillogenesis
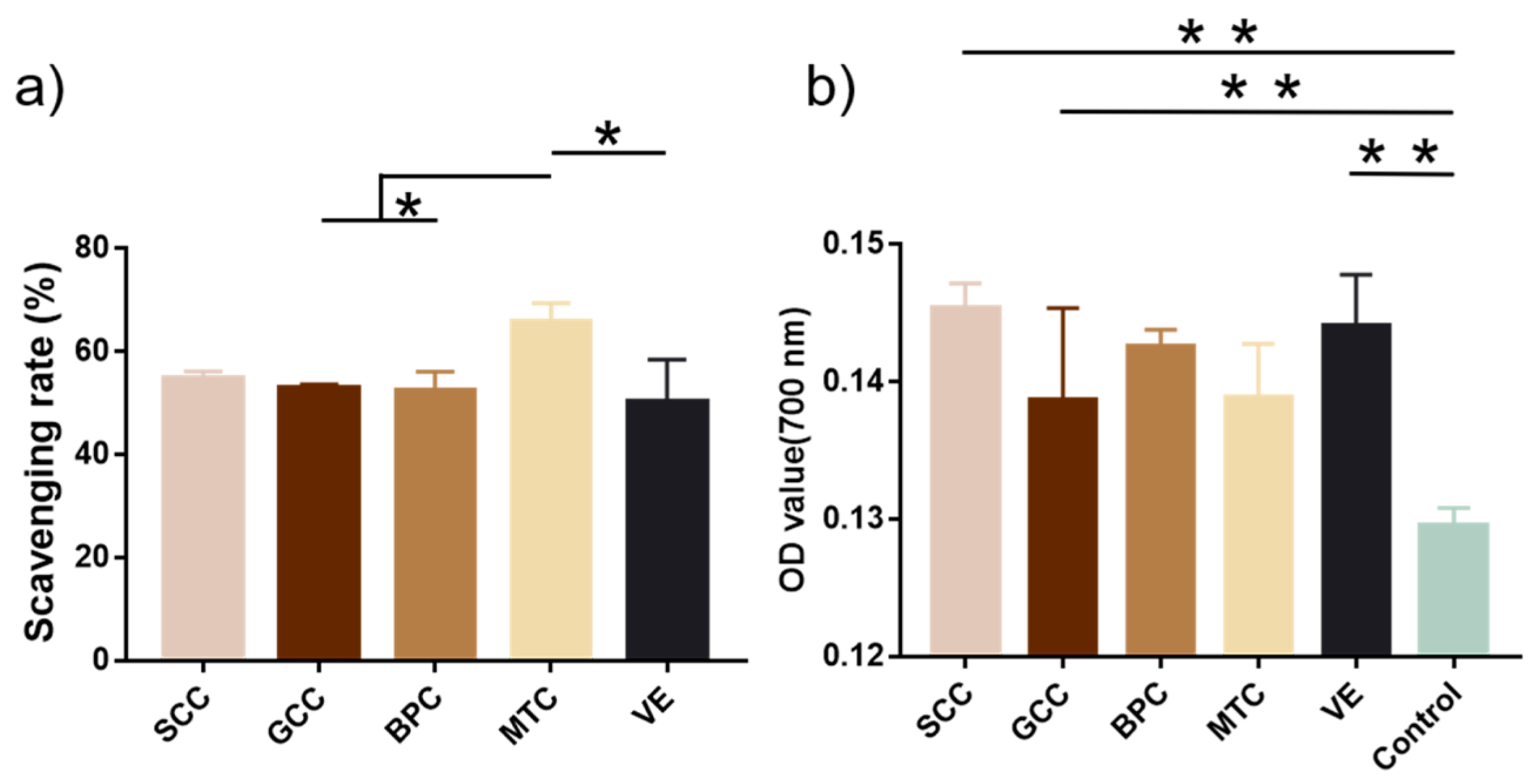
2.10. Antioxidant Activity
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Isolation and Purification of Collagen
4.3. SDS–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4. Peptide Maps
4.5. Amino Acid Analysis
4.6. Ultraviolet-Visible Spectroscopic Analysis
4.7. Fourier Transform Infrared Spectroscopic Analysis
4.8. Circular Dichroism Measurement
4.9. Collagen Fibrillogenesis In Vitro
4.10. Measurement of Degree of Collagen Fibrillogenesis
4.11. Scanning Electron Microscopy (SEM) Observation
4.12. Antioxidant Activity
4.12.1. DPPH(2,2-diphenyl-1-picrylhydrazyl) Radical Scavenging Activities
4.12.2. Reducing Power
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, L.; Ren, Y.; Emmert, S.; Vučković, I.; Stojanovic, S.; Najman, S.; Schnettler, R.; Barbeck, M.; Schenke-Layland, K.; Xiong, X. The Use of Collagen-Based Materials in Bone Tissue Engineering. Int. J. Mol. Sci. 2023, 24, 3744. [Google Scholar] [CrossRef]
- Nurilmala, M.; Suryamarevita, H.; Husein Hizbullah, H.; Jacoeb, A.M.; Ochiai, Y. Fish skin as a biomaterial for halal collagen and gelatin. Saudi J. Biol. Sci. 2022, 29, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, J.; Jiang, J.; Wang, L.; Roth, J.; Mcguinness, K.N.; Baum, J.; Dai, W.; Sun, Y.; Nanda, V.; et al. Design of synthetic collagens that assemble into supramolecular banded fibers as a functional biomaterial testbed. Nat. Commun. 2022, 13, 6761. [Google Scholar] [CrossRef]
- Jafari, H.; Lista, A.; Siekapen, M.M.; Ghaffari-Bohlouli, P.; Nie, L.; Alimoradi, H.; Shavandi, A. Fish Collagen: Extraction, Characterization, and Applications for Biomaterials Engineering. Polymers 2020, 12, 2230. [Google Scholar] [CrossRef]
- Rajabimashhadi, Z.; Gallo, N.; Salvatore, L.; Lionetto, F. Collagen Derived from Fish Industry Waste: Progresses and Challenges. Polymers 2023, 15, 544. [Google Scholar] [CrossRef]
- Moroi, S.; Miura, T.; Tamura, T.; Zhang, X.; Ura, K.; Takagi, Y. Self-assembled collagen fibrils from the swim bladder of Bester sturgeon enable alignment of MC3T3-E1 cells and enhance osteogenic differentiation. Mater. Sci. Eng. C-Mater. Biol. Appl. 2019, 104, 109925. [Google Scholar] [CrossRef] [PubMed]
- Rastian, Z.; Putz, S.; Wang, Y.; Kumar, S.; Fleissner, F.; Weidner, T.; Parekh, S.H. Type I Collagen from Jellyfish Catostylus mosaicus for Biomaterial Applications. ACS Biomater. Sci. Eng. 2018, 4, 2115–2125. [Google Scholar] [CrossRef]
- Ahmed, R.; Haq, M.; Chun, B.S. Characterization of marine derived collagen extracted from the by-products of bigeye tuna (Thunnus obesus). Int. J. Biol. Macromol. 2019, 135, 668–676. [Google Scholar] [CrossRef]
- Guan, Y.; He, J.; Chen, J.; Li, Y.; Zhang, X.; Zheng, Y.; Jia, L. Valorization of Fish Processing By-Products: Microstructural, Rheological, Functional, and Properties of Silver Carp Skin Type I Collagen. Foods 2022, 11, 2985. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, S.; Wang, Z.; Liu, S.; Li, R.; Jia, X.; Chen, J.; Liu, X.; Song, B.; Zhong, S. Anticoagulant and anti-inflammatory effects of a degraded sulfate glycosaminoglycan from swimming bladder. Food Res. Int. 2022, 157, 111444. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Wang, W.Y.; Wu, M.F.; Wang, Y.M.; Zhu, W.Y.; Chi, C.F.; Wang, B. Eighteen Novel Bioactive Peptides from Monkfish (Lophius litulon) Swim Bladders: Production, Identification, Antioxidant Activity, and Stability. Mar. Drugs 2023, 21, 169. [Google Scholar] [CrossRef]
- Cai, S.Y.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H(2)O(2)-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury. Int. J. Mol. Sci. 2019, 20, 5425. [Google Scholar] [CrossRef]
- Zheng, J.; Tian, X.; Xu, B.; Yuan, F.; Gong, J.; Yang, Z. Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H(2)O(2)-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells. Mar. Drugs 2020, 18, 430. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Q.; Zeng, L.; Yang, Z.S.; Huang, F.F.; Ding, G.F.; Wang, B. Anti-Fatigue Effect by Peptide Fraction from Protein Hydrolysate of Croceine Croaker (Pseudosciaena crocea) Swim Bladder through Inhibiting the Oxidative Reactions including DNA Damage. Mar. Drugs 2016, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, K.P.; Nidheesh, T.; Govindaraju, K.; Jyoti; Suresh, P.V. Enzymatic extraction and characterisation of a thermostable collagen from swim bladder of rohu (Labeo rohita). J. Sci. Food Agric. 2017, 97, 1451–1458. [Google Scholar] [CrossRef]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and characterisation of collagen extracted from the skin of striped catfish (Pangasianodon hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, R.; Ye, C.; Konno, K. Isolation and characterization of collagens from scale of silver carp (Hypophthalmichthys molitrix). J. Food Biochem. 2010, 34, 1343–1354. [Google Scholar] [CrossRef]
- Zhang, X.; Adachi, S.; Ura, K.; Takagi, Y. Properties of collagen extracted from Amur sturgeon Acipenser schrenckii and assessment of collagen fibrils in vitro. Int. J. Biol. Macromol. 2019, 137, 809–820. [Google Scholar] [CrossRef]
- Pal, G.K.; Suresh, P.V. Physico-chemical characteristics and fibril-forming capacity of carp swim bladder collagens and exploration of their potential bioactive peptides by in silico approaches. Int. J. Biol. Macromol. 2017, 101, 304–313. [Google Scholar] [CrossRef]
- Gao, L.; Orth, P.; Cucchiarini, M.; Madry, H. Effects of solid acellular type-I/III collagen biomaterials on in vitro and in vivo chondrogenesis of mesenchymal stem cells. Expert Rev. Med. Devices 2017, 14, 717–732. [Google Scholar] [CrossRef]
- Kaewdang, O.; Benjakul, S.; Kaewmanee, T.; Kishimura, H. Characteristics of collagens from the swim bladders of yellowfin tuna (Thunnus albacares). Food Chem. 2014, 155, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (Hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Appel, W. Chymotrypsin: Molecular and catalytic properties. Clin. Biochem. 1986, 19, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Kishimura, H.; Shahidi, F. Isolation and Characterisation of collagen from the skin of brownbanded bamboo shark (Chiloscyllium punctatum). Food Chem. 2010, 119, 1519–1526. [Google Scholar] [CrossRef]
- Burjanadze, T.V. New analysis of the phylogenetic change of collagen thermostability. Biopolymers 2000, 53, 523–528. [Google Scholar] [CrossRef]
- Rigby, B.J. Amino-acid composition and thermal stability of the skin collagen of the Antarctic ice-fish. Nature 1968, 219, 166–167. [Google Scholar] [CrossRef]
- Pal, G.K.; Suresh, P.V. Comparative assessment of physico-chemical characteristics and fibril formation capacity of thermostable carp scales collagen. Mater. Sci. Eng. C-Mater. Biol. Appl. 2017, 70, 32–40. [Google Scholar] [CrossRef]
- Liao, W.; Guanghua, X.; Li, Y.; Shen, X.R.; Li, C. Comparison of characteristics and fibril-forming ability of skin collagen from barramundi (Lates calcarifer) and tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2018, 107, 549–559. [Google Scholar] [CrossRef]
- Zhao, W.H.; Chi, C.F.; Zhao, Y.Q.; Wang, B. Preparation, Physicochemical and Antioxidant Properties of Acid- and Pepsin-Soluble Collagens from the Swim Bladders of Miiuy Croaker (Miichthys miiuy). Mar. Drugs 2018, 16, 161. [Google Scholar] [CrossRef]
- Jeevithan, E.; Jingyi, Z.; Wang, N.; He, L.; Bao, B.; Wu, W. Physico-chemical, antioxidant and intestinal absorption properties of whale shark type-II collagen based on its solubility with acid and pepsin. Process Biochem. 2015, 50, 463–472. [Google Scholar] [CrossRef]
- Doyle, B.B.; Bendit, E.G.; Blout, E.R. Infrared spectroscopy of collagen and collagen-like polypeptides. Biopolymers 1975, 14, 937–957. [Google Scholar] [CrossRef] [PubMed]
- Riaz, T.; Zeeshan, R.; Zarif, F.; Ilyas, K.; Muhammad, N.; Safi, S.Z.; Rahim, A.; Rizvi, S.A.A.; Rehman, I.U. FTIR analysis of natural and synthetic collagen. Appl. Spectrosc. Rev. 2018, 53, 703–746. [Google Scholar] [CrossRef]
- Gauza-Włodarczyk, M.; Kubisz, L.; Mielcarek, S.; Włodarczyk, D. Comparison of thermal properties of fish collagen and bovine collagen in the temperature range 298-670K. Mater. Sci. Eng. C-Mater. Biol. Appl. 2017, 80, 468–471. [Google Scholar] [CrossRef]
- Murphy, B.M.; D’Antonio, J.; Manning, M.C.; Al-Azzam, W. Use of the amide II infrared band of proteins for secondary structure determination and comparability of higher order structure. Curr. Pharm. Biotechnol. 2014, 15, 880–889. [Google Scholar] [CrossRef]
- Cai, S.; Singh, B.R. A distinct utility of the amide III infrared band for secondary structure estimation of aqueous protein solutions using partial least squares methods. Biochemistry 2004, 43, 2541–2549. [Google Scholar] [CrossRef]
- Oliveira, V.D.M.; Assis, C.R.D.; Costa, B.D.A.M.; Neri, R.C.D.A.; Monte, F.T.D.; Freitas, H.M.S.D.; França, R.C.P.; Santos, J.F.; Bezerra, R.D.S.; Porto, A.L.F. Physical, biochemical, densitometric and spectroscopic techniques for characterization collagen from alternative sources: A review based on the sustainable valorization of aquatic by-products. J. Mol. Struct. 2021, 1224, 129023. [Google Scholar] [CrossRef]
- Sun, L.; Hou, H.; Li, B.; Zhang, Y. Characterization of acid- and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2017, 99, 8–14. [Google Scholar] [CrossRef]
- Mu, C.; Li, D.; Lin, W.; Ding, Y.; Zhang, G. Temperature induced denaturation of collagen in acidic solution. Biopolymers 2007, 86, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Kaneniwa, M.; Wang, X.; Chen, S.; Cheng, Y.; Qu, Y.; Fukuda, Y.; Konno, K. Seasonal Expression of 2 Types of Myosin with Different Thermostability in Silver Carp Muscle (Hypophthalmichthys molitrix). J. Food Sci. 2006, 71, C39–C43. [Google Scholar] [CrossRef]
- Pal, G.K.; Nidheesh, T.; Suresh, P.V. Comparative study on characteristics and in vitro fibril formation ability of acid and pepsin soluble collagen from the skin of catla (Catla catla) and rohu (Labeo rohita). Food Res. Int. 2015, 76, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (Rhopilema esculentum) on mice skin photoaging induced by UV irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Sinthusamran, S.; Benjakul, S.; Kishimura, H. Comparative study on molecular characteristics of acid soluble collagens from skin and swim bladder of seabass (Lates calcarifer). Food Chem. 2013, 138, 2435–2441. [Google Scholar] [CrossRef] [PubMed]
- Matmaroh, K.; Benjakul, S.; Prodpran, T.; Encarnacion, A.B.; Kishimura, H. Characteristics of acid soluble collagen and pepsin soluble collagen from scale of spotted golden goatfish (Parupeneus heptacanthus). Food Chem. 2011, 129, 1179–1186. [Google Scholar] [CrossRef]
- Wei, B.; Zenaidee, M.A.; Lantz, C.; Williams, B.J.; Totten, S.; Ogorzalek, L.R.; Loo, J.A. Top-down mass spectrometry and assigning internal fragments for determining disulfide bond positions in proteins. Analyst 2022, 148, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Akita, M.; Nishikawa, Y.; Shigenobu, Y.; Ambe, D.; Morita, T.; Morioka, K.; Adachi, K. Correlation of proline, hydroxyproline and serine content, denaturation temperature and circular dichroism analysis of type I collagen with the physiological temperature of marine teleosts. Food Chem. 2020, 329, 126775. [Google Scholar] [CrossRef]
- Cisneros, D.A.; Hung, C.; Franz, C.M.; Muller, D.J. Observing growth steps of collagen self-assembly by time-lapse high-resolution atomic force microscopy. J. Struct. Biol. 2006, 154, 232–245. [Google Scholar] [CrossRef]
- Venkatesan, J.; Anil, S.; Kim, S.K.; Shim, M.S. Marine Fish Proteins and Peptides for Cosmeceuticals: A Review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Jeevithan, E.; Bao, B.; Bu, Y.; Zhou, Y.; Zhao, Q.; Wu, W. Type II collagen and gelatin from silvertip shark (Carcharhinus albimarginatus) cartilage: Isolation, purification, physicochemical and antioxidant properties. Mar. Drugs 2014, 12, 3852–3873. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, B.; Jing, H.; Wu, Y.; Kong, D.; Leng, X.; Wang, Z. Swim Bladder as a Novel Biomaterial for Cardiovascular Materials with Anti-Calcification Properties. Adv. Healthc. Mater. 2020, 9, e1901154. [Google Scholar] [CrossRef]
- Sheng, Y.; Qiu, Y.T.; Wang, Y.M.; Chi, C.F.; Wang, B. Novel Antioxidant Collagen Peptides of Siberian Sturgeon (Acipenserbaerii) Cartilages: The Preparation, Characterization, and Cytoprotection of H(2)O(2)-Damaged Human Umbilical Vein Endothelial Cells (HUVECs). Mar. Drugs 2022, 20, 325. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Wang, J.; He, Y.; Chi, C.; Wang, B. Antioxidant peptides from protein hydrolysate of skipjack tuna milt: Purification, identification, and cytoprotection on H2O2 damaged human umbilical vein endothelial cells. Process Biochem. 2022, 113, 258–269. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Collagens | dECMs | ||||
|---|---|---|---|---|---|---|
| SC | GC | BP | SC | GC | BP | |
| Histidine | 4.06 | 4.22 | 4.10 | 5.93 | 5.53 | 3.89 |
| Hydroxyproline | 65.81 | 68.96 | 72.77 | 47.60 | 33.08 | 66.64 |
| Arginine | 89.23 | 95.38 | 94.62 | 83.80 | 75.22 | 85.29 |
| Serine | 33.35 | 37.03 | 32.23 | 34.92 | 36.43 | 29.80 |
| Glycine | 196.44 | 204.50 | 198.35 | 173.50 | 173.12 | 180.46 |
| Aspartic acid | 76.61 | 76.89 | 72.57 | 83.61 | 85.28 | 65.70 |
| Glutamic acid | 108.81 | 110.83 | 108.05 | 104.54 | 94.06 | 96.20 |
| Threonine | 28.86 | 29.60 | 18.61 | 34.05 | 40.48 | 16.58 |
| Alanine | 97.75 | 103.20 | 101.48 | 88.76 | 84.67 | 90.75 |
| Proline | 142.87 | 149.21 | 155.08 | 124.37 | 113.37 | 136.70 |
| Lysine | 16.92 | 18.39 | 17.04 | 14.62 | 13.33 | 12.72 |
| Cystine | 1.33 | 1.13 | 0.00 | 5.45 | 5.08 | 0.00 |
| Methionine | 15.24 | 11.37 | 5.01 | 17.00 | 12.97 | 4.90 |
| Tyrosine | 5.78 | 7.31 | 4.09 | 27.81 | 49.44 | 5.64 |
| Valine | 16.77 | 19.52 | 19.41 | 23.60 | 28.96 | 17.73 |
| Isoleucine | 14.06 | 15.93 | 13.16 | 18.92 | 21.90 | 11.76 |
| Leucine | 29.36 | 32.53 | 27.22 | 41.50 | 55.53 | 25.29 |
| Phenylalanine | 23.78 | 24.37 | 19.78 | 25.94 | 23.88 | 18.91 |
| Tryptophan | 0.45 | 0.07 | 0.04 | 0.02 | 0.02 | 0.07 |
| CD [221 nm] (mdeg) | Rpn | |
|---|---|---|
| SCC | 28.6594 | 0.3566 |
| GCC | 8.75611 | 0.1530 |
| BPC | 14.4292 | 0.2533 |
| MTC | 18.7925 | 0.1958 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Kong, Y.; Song, L.; Liu, J.; Wang, Z. A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys molitrix). Mar. Drugs 2023, 21, 280. https://doi.org/10.3390/md21050280
Jiang H, Kong Y, Song L, Liu J, Wang Z. A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys molitrix). Marine Drugs. 2023; 21(5):280. https://doi.org/10.3390/md21050280
Chicago/Turabian StyleJiang, Honghui, Yuanyuan Kong, Lili Song, Jing Liu, and Zhihong Wang. 2023. "A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys molitrix)" Marine Drugs 21, no. 5: 280. https://doi.org/10.3390/md21050280
APA StyleJiang, H., Kong, Y., Song, L., Liu, J., & Wang, Z. (2023). A Thermostable Type I Collagen from Swim Bladder of Silver Carp (Hypophthalmichthys molitrix). Marine Drugs, 21(5), 280. https://doi.org/10.3390/md21050280