
Efficient Degradation of Alginate and Preparation of Alginate Oligosaccharides by a Novel Biofunctional Alginate Lyase with High Activity and Excellent Thermophilic Features



Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis
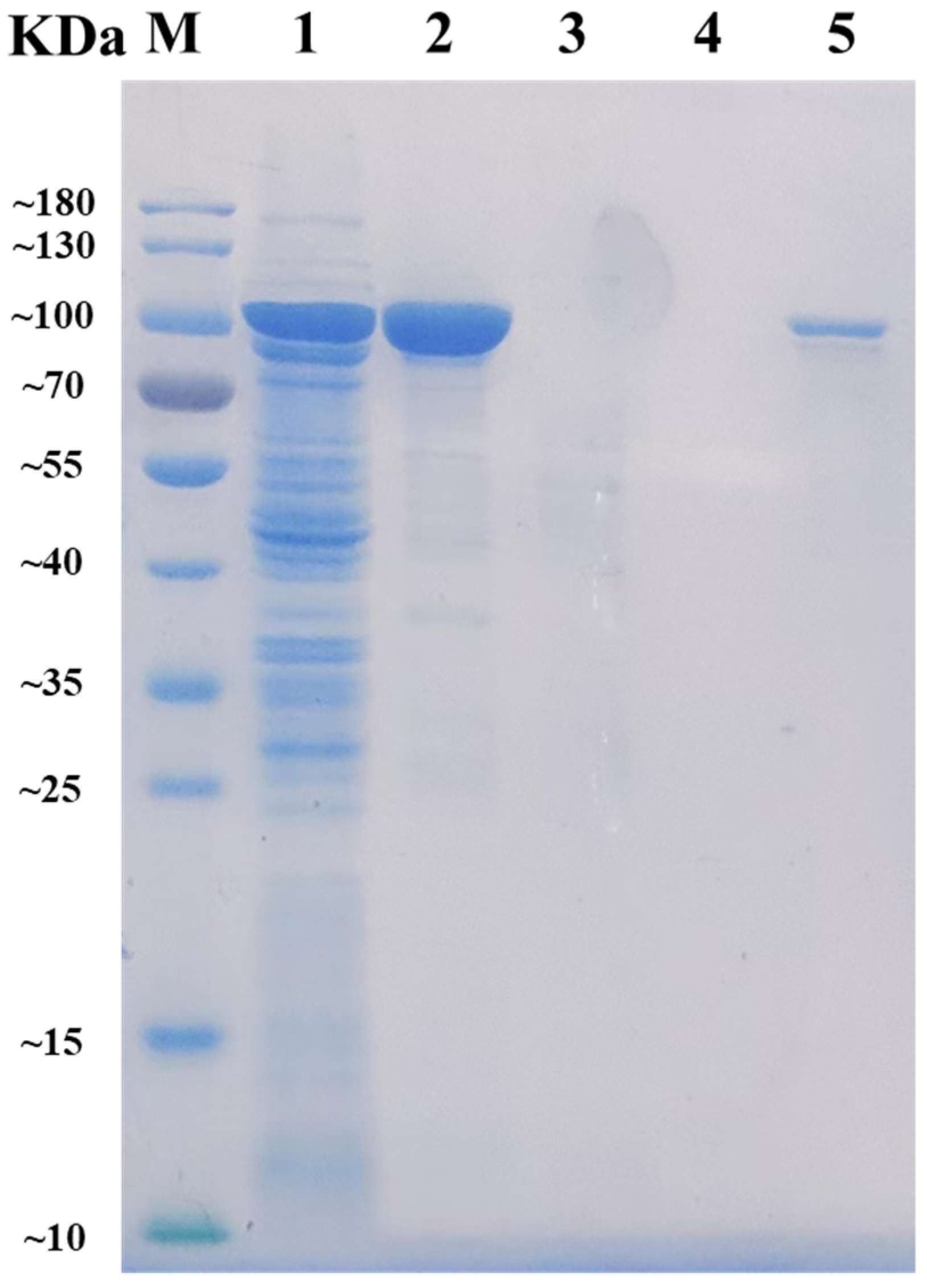
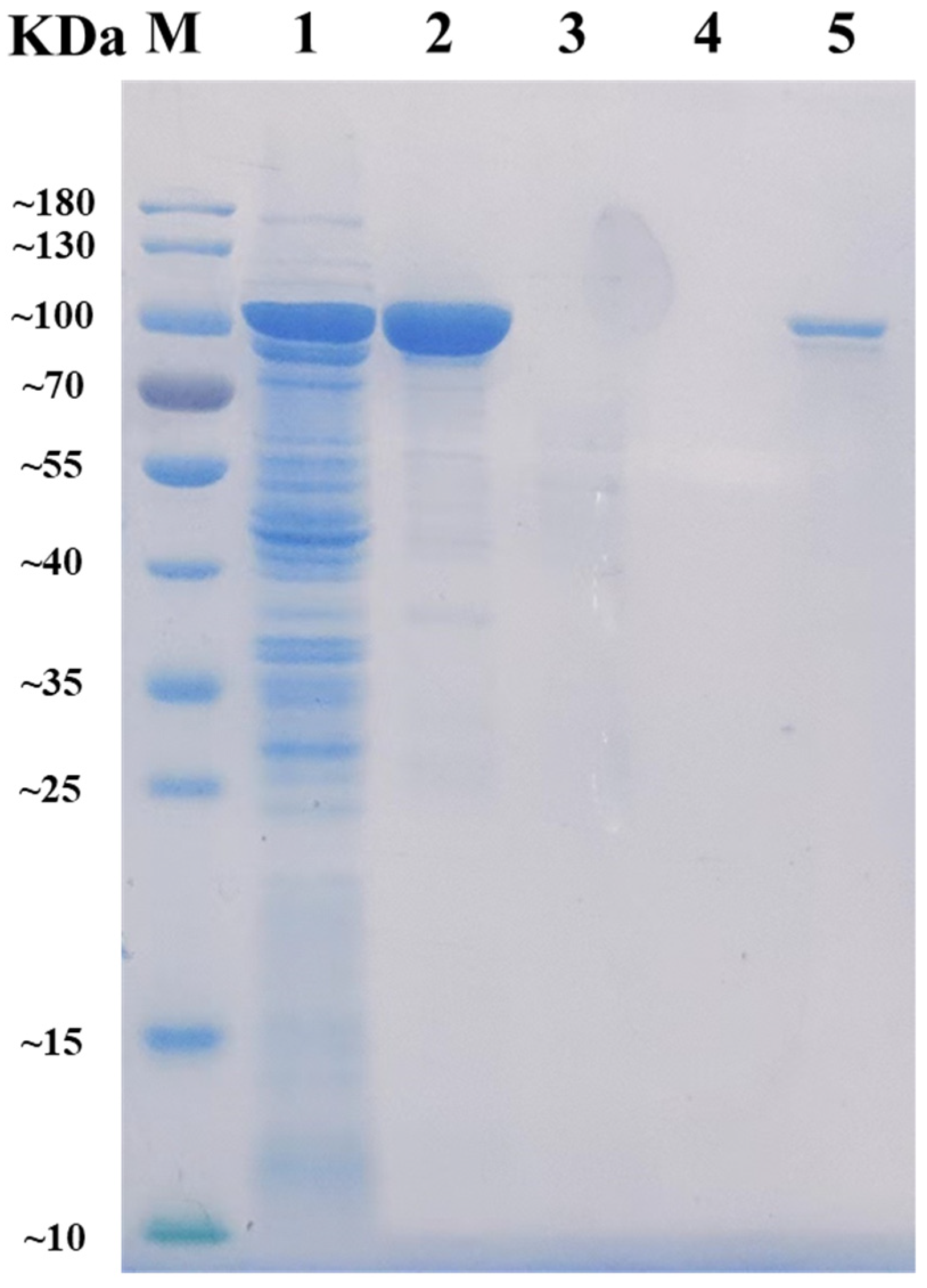
2.2. Expression and Purification of AlyRm3
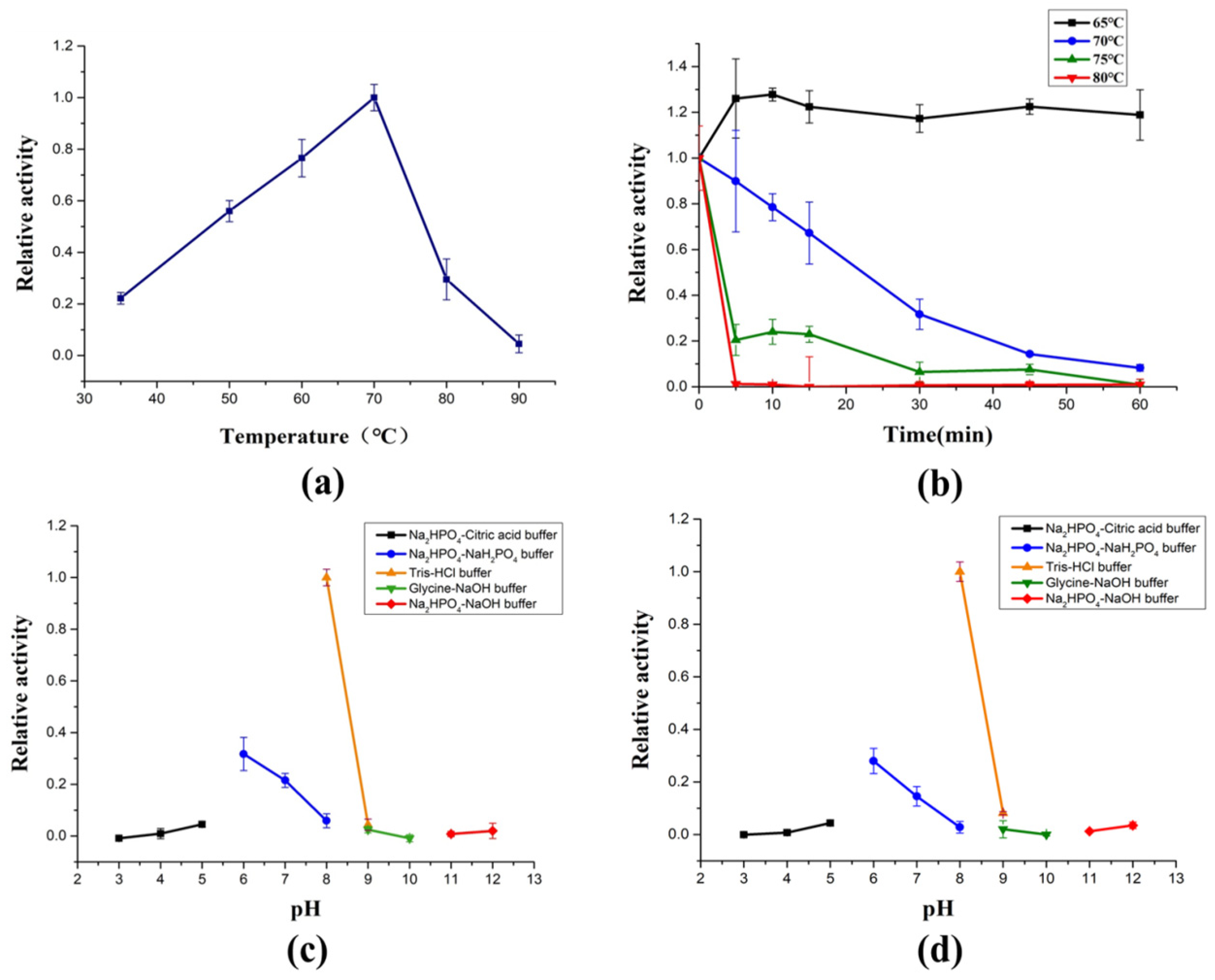
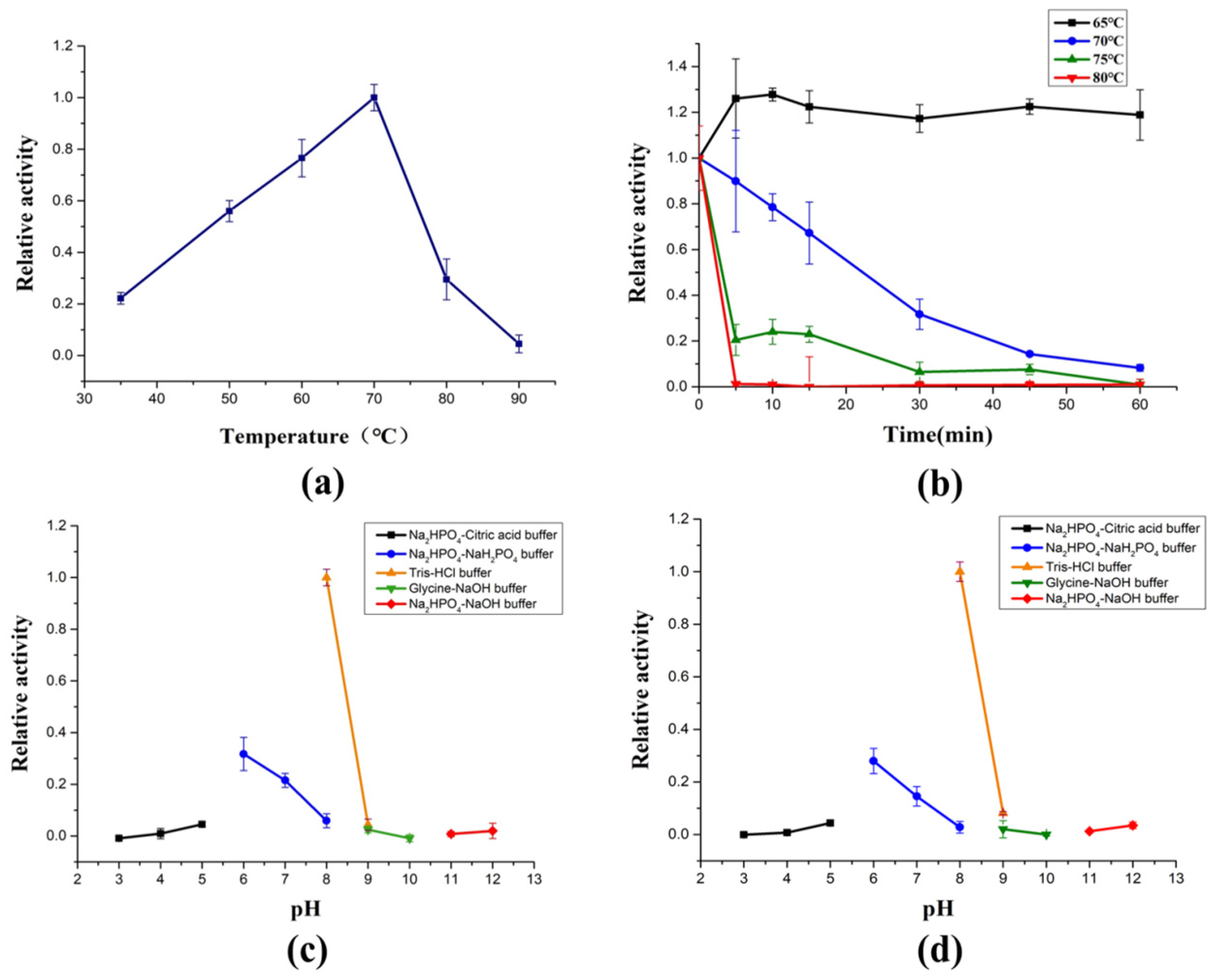
2.3. Biochemical Characterization of AlyRm3
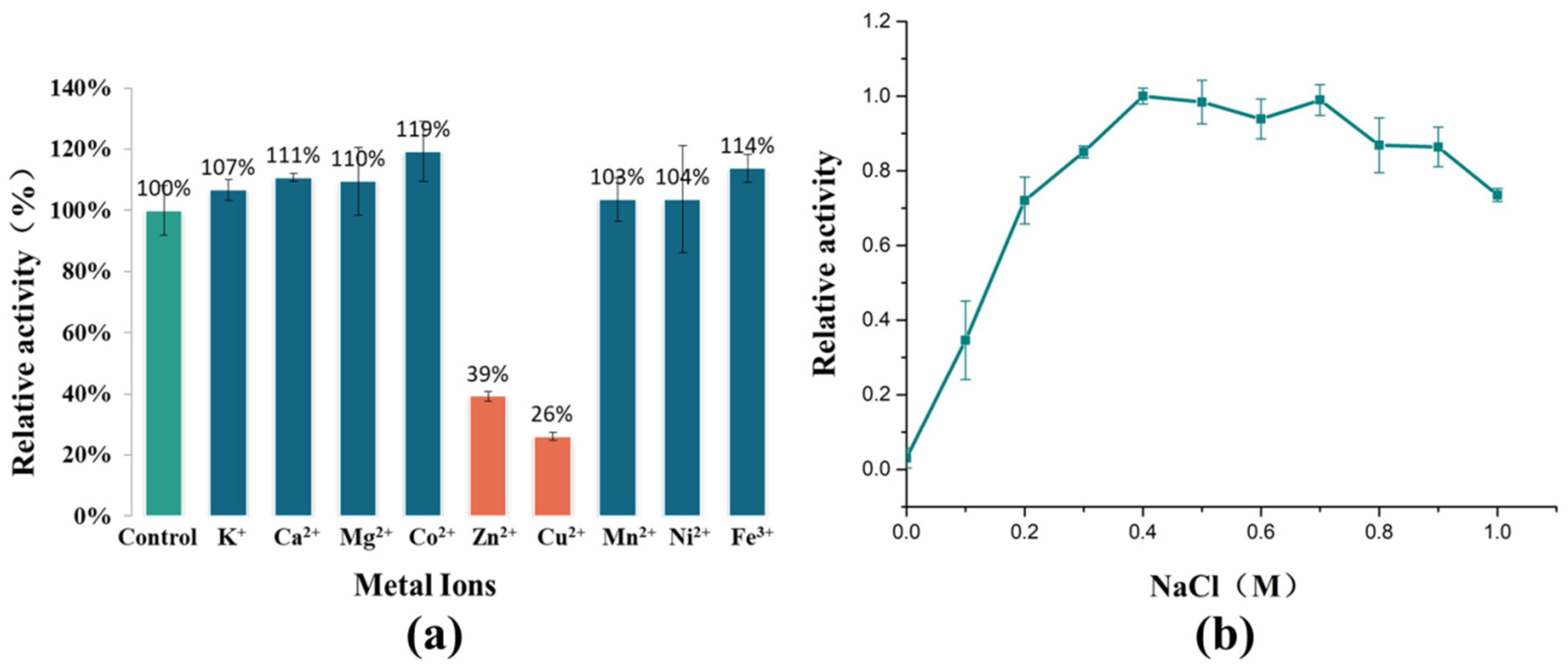
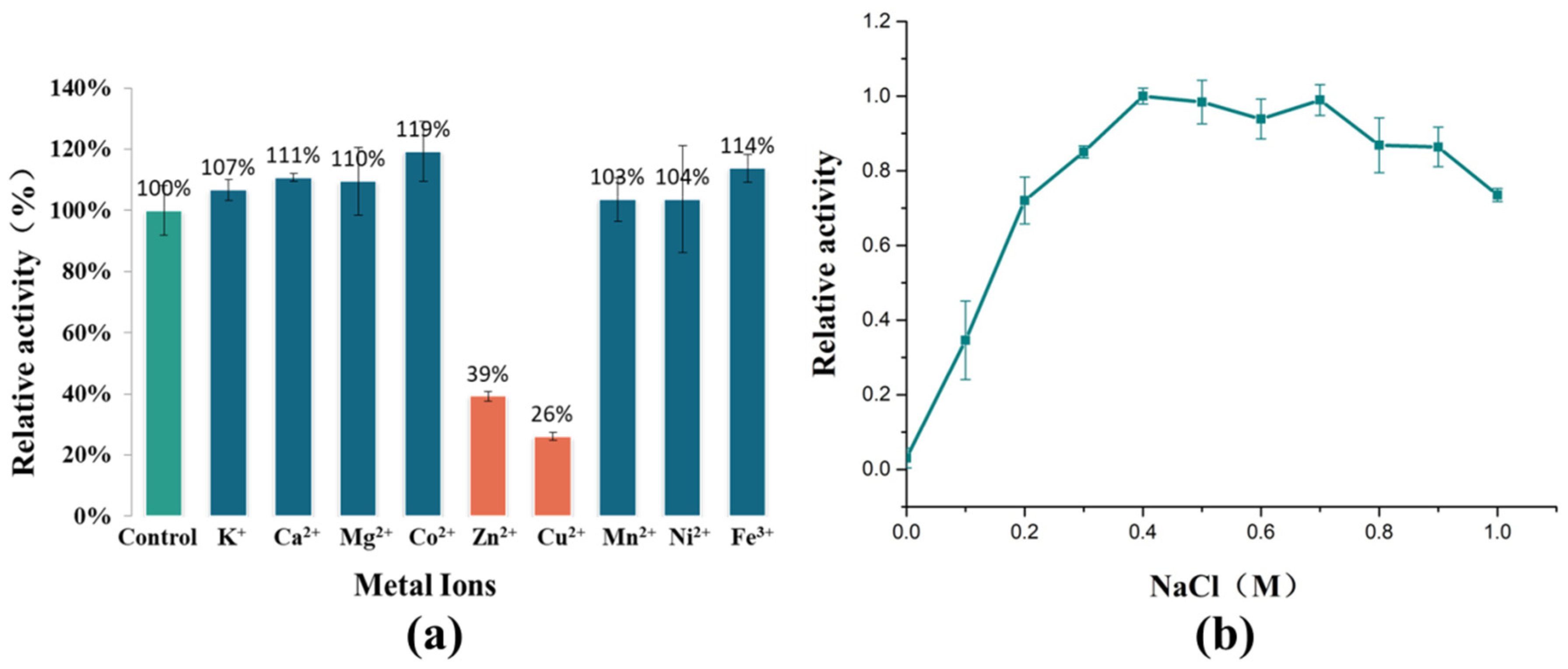
2.4. Effect of Metal Ions on AlyRm3
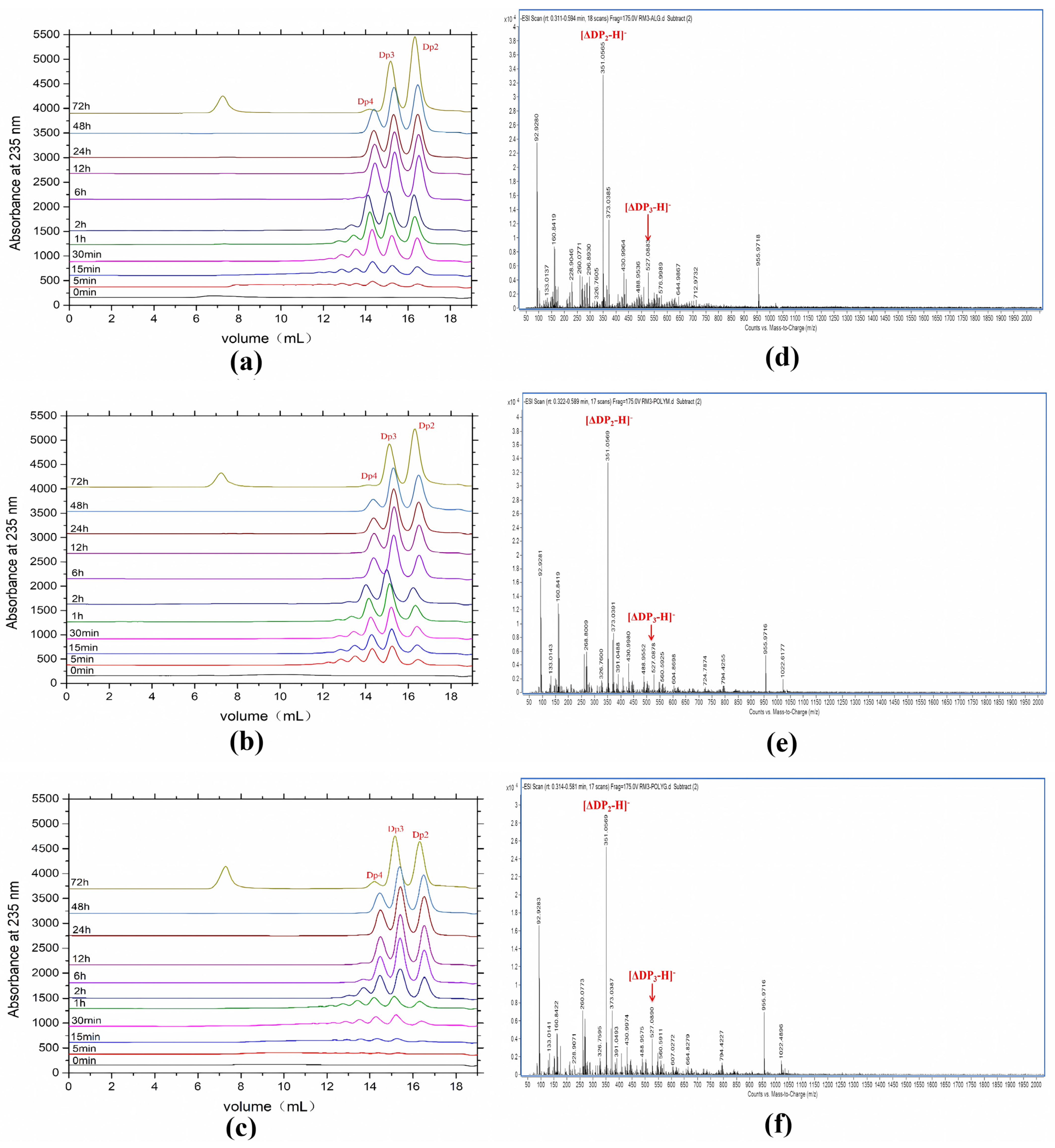
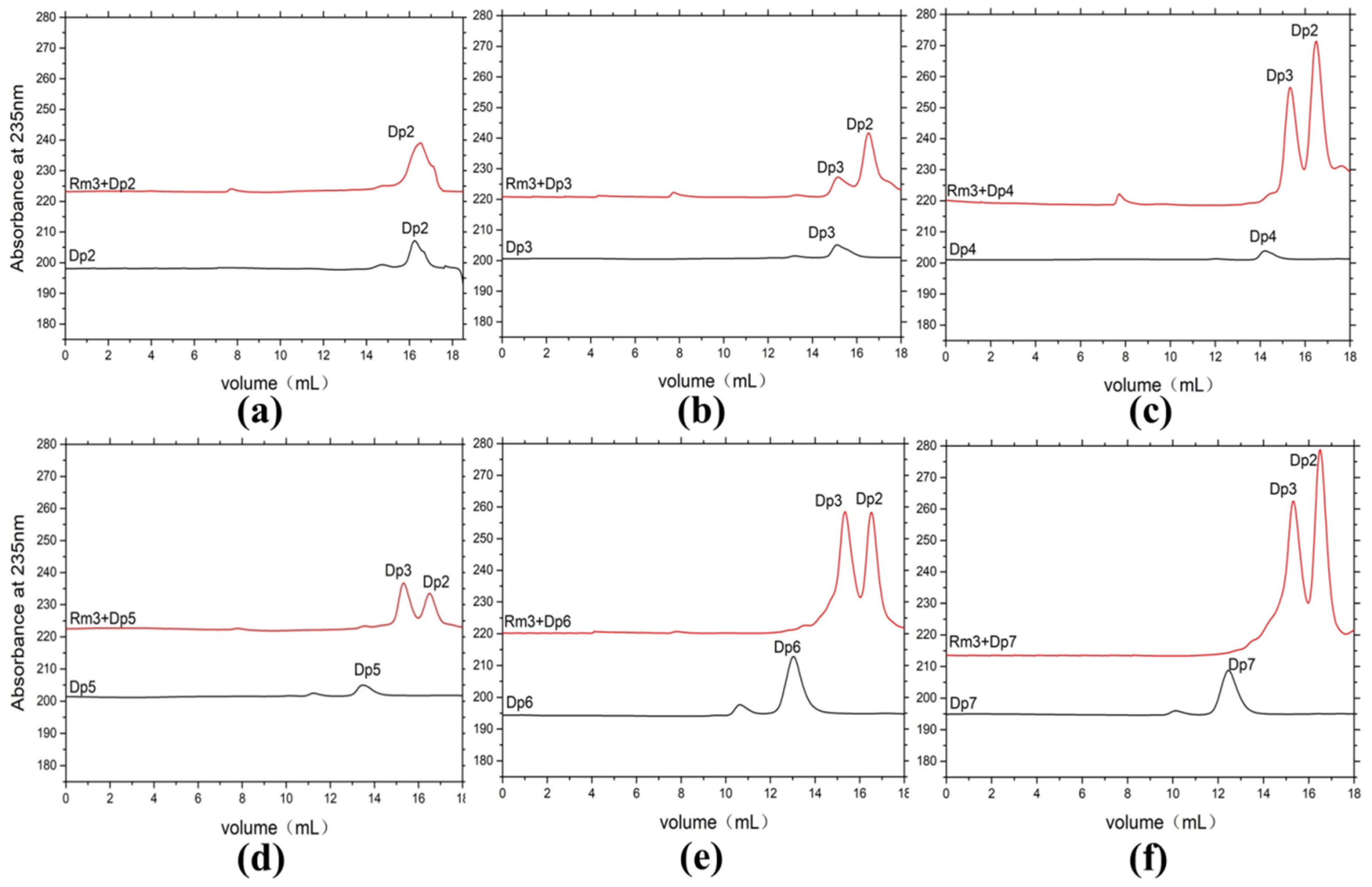
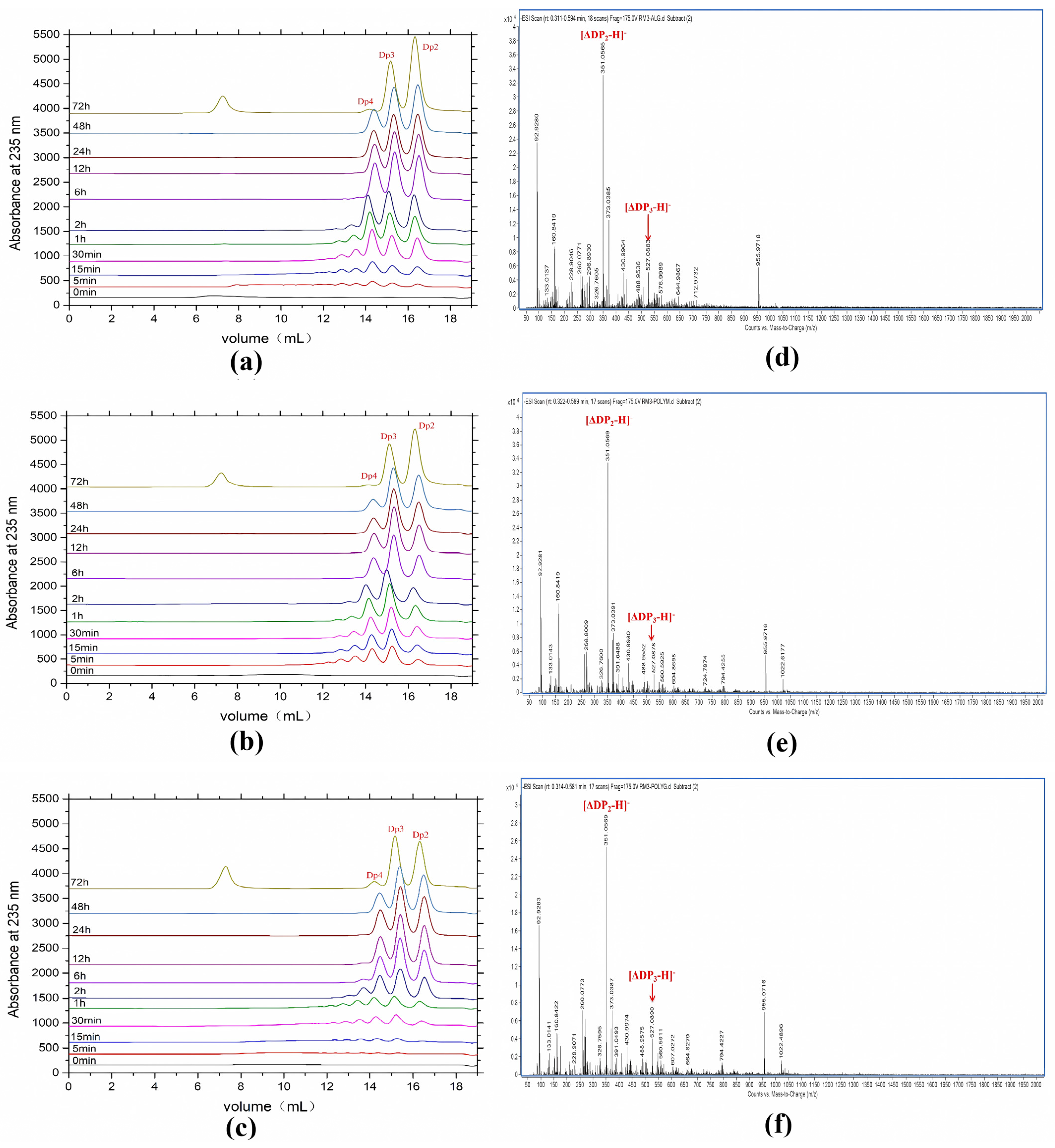
2.5. Products Distribution and Action Pattern of AlyRm3
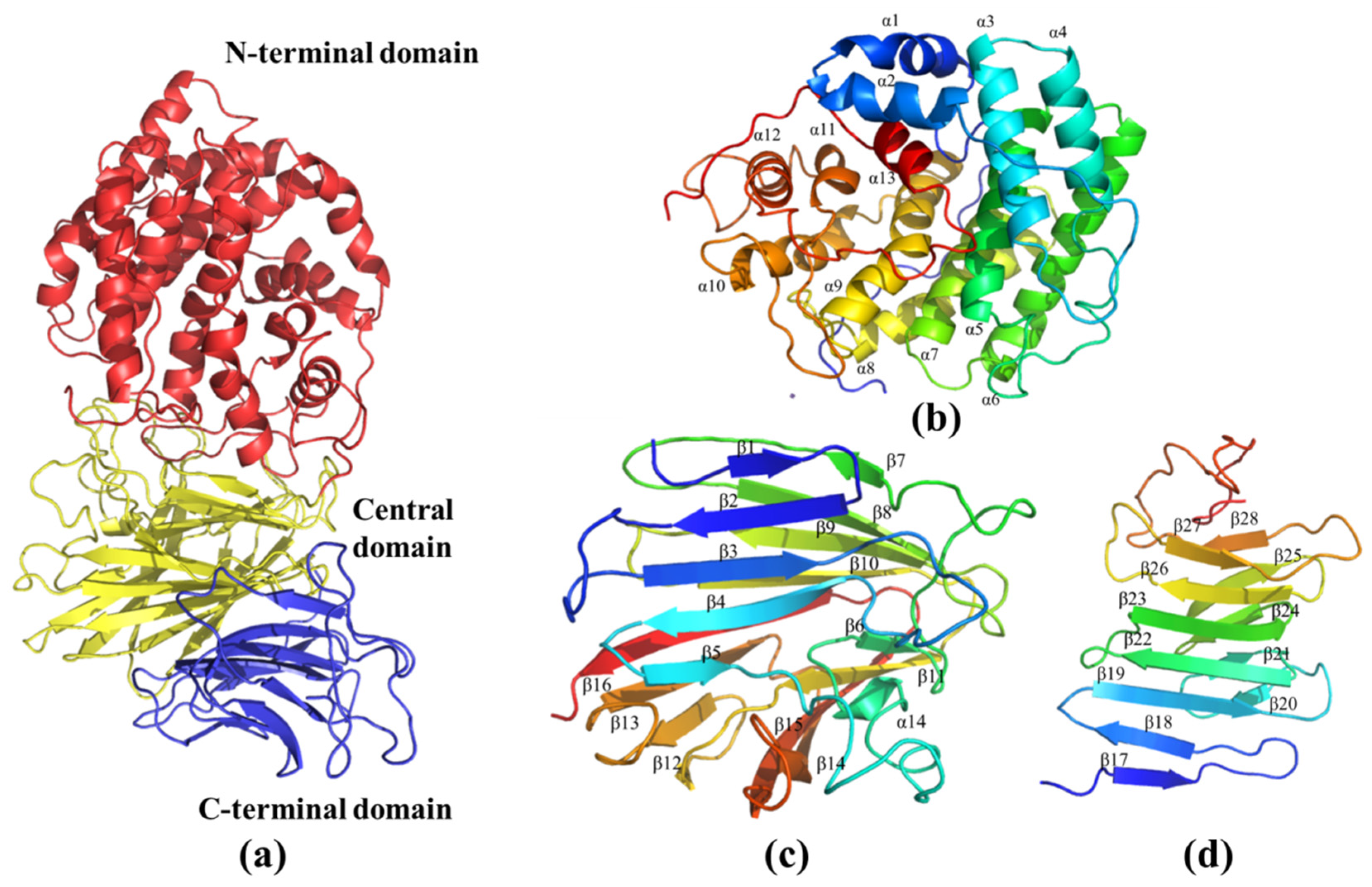
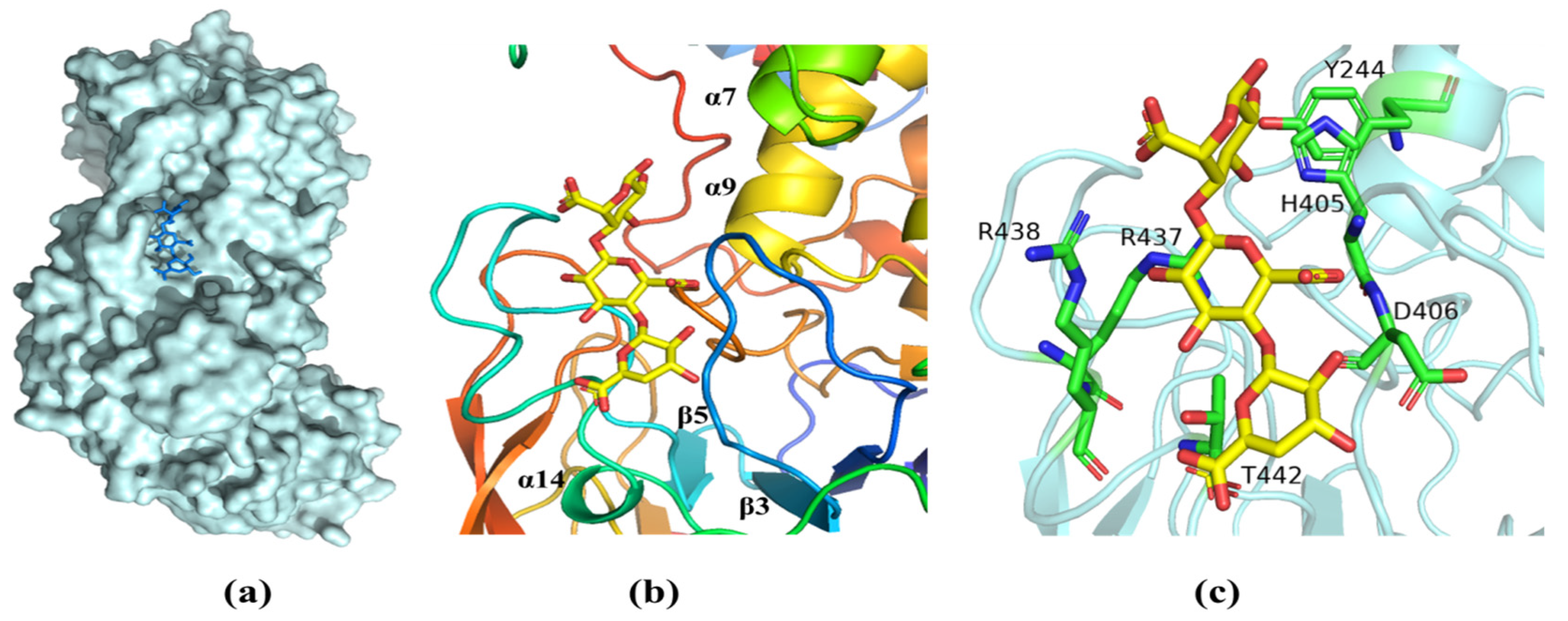
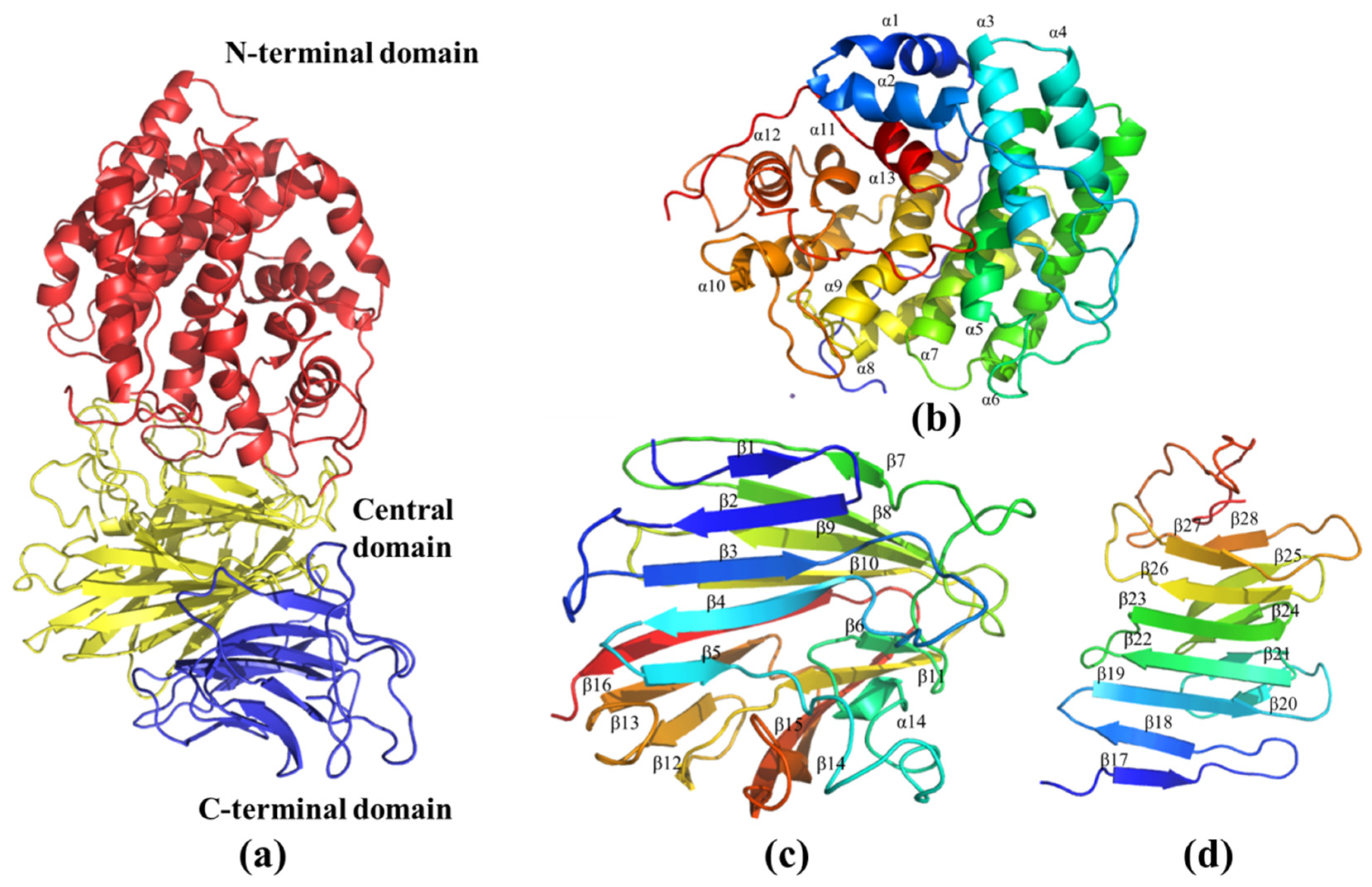
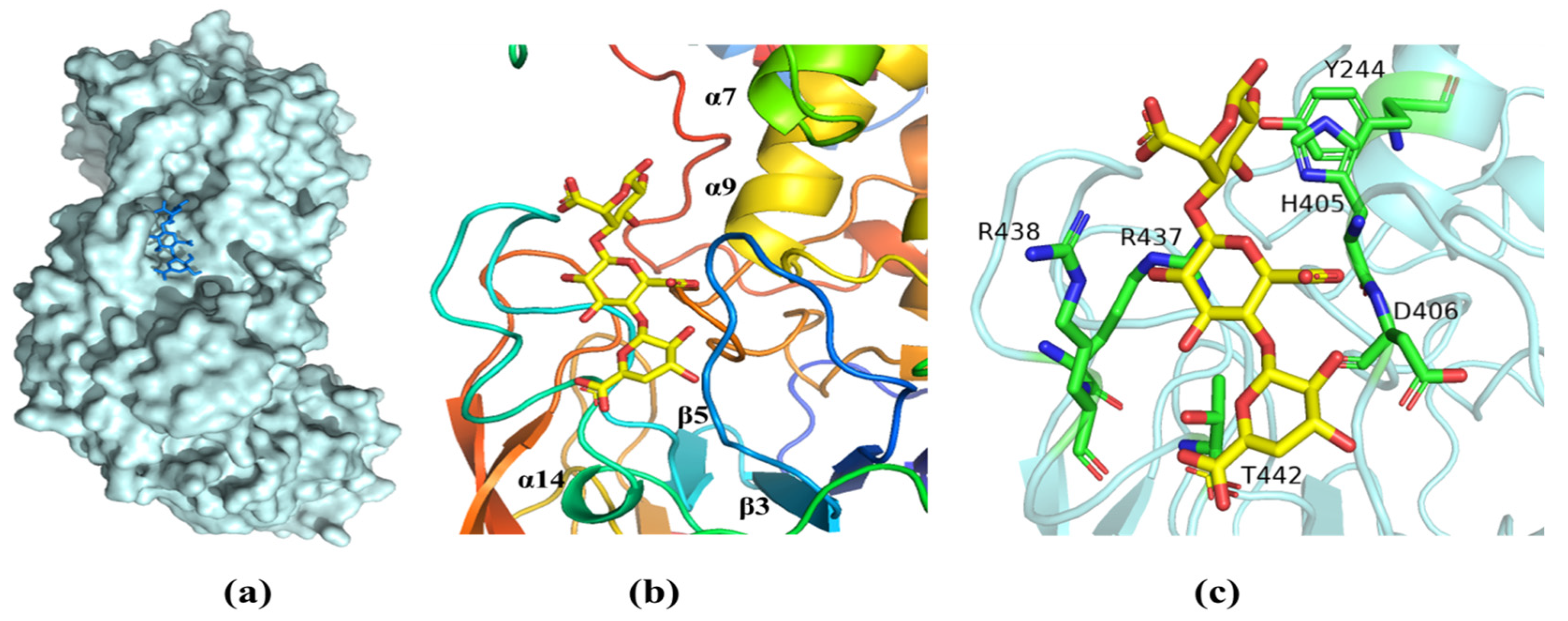
2.6. Molecular Modeling
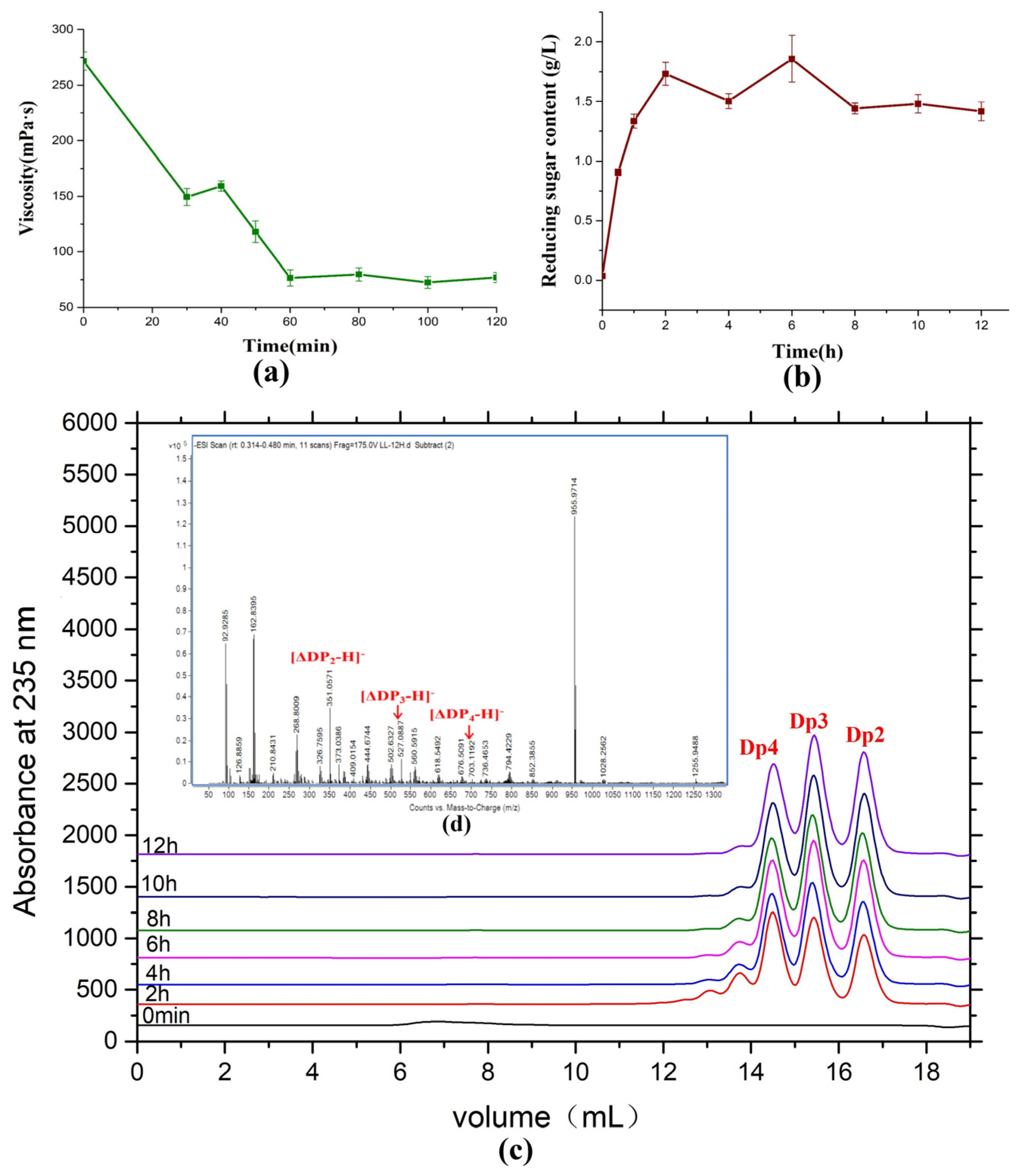
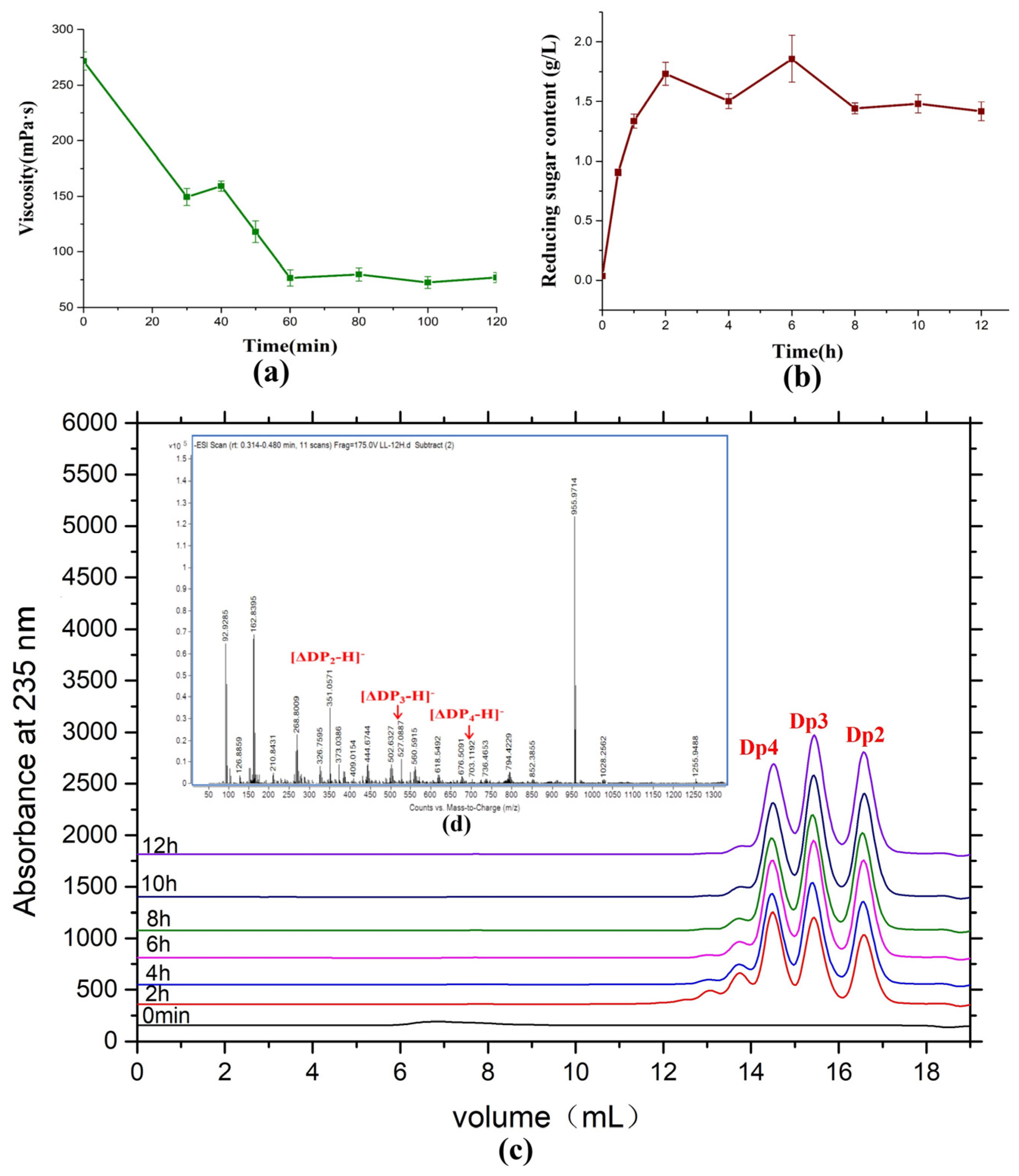
2.7. Saccharification of Alginate by AlyRm3
3. Discussion
4. Experimental Materials and Methods
4.1. Sequence Analysis
4.2. Cloning, Heterologous Expression, and Purification of AlyRm3
4.3. Substrate Specificity and Enzyme Kinetic Constants
4.4. Biochemical Characterization of AlyRm3
4.5. Products Distribution and Action Pattern Analysis of AlyRm3
4.6. Saccharification of Alginate by AlyRm3
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vasudevan, U.M.; Lee, O.K.; Lee, E.Y. Alginate derived functional oligosaccharides: Recent developments, barriers, and future outlooks. Carbohydr. Polym. 2021, 267, 118–158. [Google Scholar]
- Cheng, D.Y.; Jiang, C.C.; Xu, J.C.; Liu, Z.; Mao, X.Z. Characteristics and applications of alginate lyases: A review. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Li, L.; Li, Q.; Jiang, L.; Zhu, B.; Yao, Z. A novel alginate lyase and its domain functions for the preparation of unsaturated monosaccharides. Appl. Microbiol. Biotechnol. 2023, 107, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Castellano, M.; Alloisio, M.; Darawish, R.; Dodero, A.; Vicini, S. Electrospun composite mats of alginate with embedded silver nanoparticles: Synthesis and characterization. J. Therm. Anal. Calorim. 2019, 137, 767–778. [Google Scholar] [CrossRef]
- Dodero, A.; Alloisio, M.; Vicini, S.; Castellano, M. Preparation of composite alginate-based electrospun membranes loaded with ZnO nanoparticles. Carbohydr. Polym. 2020, 227, 115371. [Google Scholar] [CrossRef]
- Dodero, A.; Scarfi, S.; Pozzolini, M.; Vicini, S.; Alloisio, M.; Castellano, M. Alginate-Based Electrospun Membranes Containing ZnO Nanoparticles as Potential Wound Healing Patches: Biological, Mechanical, and Physicochemical Characterization. Acs Appl. Mater. Interfaces 2020, 12, 3371–3381. [Google Scholar] [CrossRef]
- Wang, B.; Wan, Y.S.; Zheng, Y.L.; Lee, X.Q.; Liu, T.Z.; Yu, Z.B.; Huang, J.; Ok, Y.S.; Chen, J.; Gao, B. Alginate-based composites for environmental applications: A critical review. Crit. Rev. Environ. Sci. Technol. 2019, 49, 318–356. [Google Scholar] [CrossRef]
- Pham, L.H.P.; Ly, K.L.; Colon-Ascanio, M.; Ou, J.; Wang, H.; Lee, S.W.; Wang, Y.; Choy, J.S.; Phillips, K.S.; Luo, X. Dissolvable alginate hydrogel-based biofilm microreactors for antibiotic susceptibility assays. Biofilm 2023, 5, 100103. [Google Scholar] [CrossRef]
- Zhu, B.W.; Huang, L.S.X.; Tan, H.D.; Qin, Y.Q.; Du, Y.G.; Yin, H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol. Lett. 2015, 37, 409–415. [Google Scholar] [CrossRef]
- Barzkar, N.; Sheng, R.; Sohail, M.; Jahromi, S.T.; Babich, O.; Sukhikh, S.; Nahavandi, R. Alginate lyases from marine bacteria: An enzyme ocean for sustainable future. Molecules 2022, 27, 3375. [Google Scholar] [CrossRef]
- Inoue, A.; Ojima, T. Functional identification of alginate lyase from the brown alga Saccharina japonica. Sci. Rep. 2019, 9, 4937. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Dai, A.; Liu, Y.; Li, T. Sustainable alginate lyases catalyzed degradation of bio-based carbohydrates. Front. Chem. 2022, 10, 1008010. [Google Scholar] [CrossRef]
- Sasaki, Y.; Yoshikuni, Y. Metabolic engineering for valorization of macroalgae biomass. Metab. Eng. 2022, 71, 42–61. [Google Scholar] [CrossRef]
- Di Donato, P.; Buono, A.; Poli, A.; Finore, I.; Abbamondi, G.R.; Nicolaus, B.; Lama, L. Exploring marine environments for the identification of extremophiles and their enzymes for sustainable and green bioprocesses. Sustainability 2019, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, X.; Zhang, X.; Li, Y.; Wang, L. Bacterial alginate metabolism: An important pathway for bioconversion of brown algae. Biotechnol. Biofuels 2021, 14, 158. [Google Scholar] [CrossRef]
- Amtzen, M.O.; Pedersen, B.; Klau, L.J.; Stokke, R.; Oftebro, M.; Antonsen, S.G.; Fredriksen, L.; Sletta, H.; Aarstad, O.A.; Aachmann, F.L.; et al. Alginate degradation: Insights obtained through characterization of a thermophilic exolytic alginate lyase. Appl. Environ. Microbiol. 2021, 87, e02399-20. [Google Scholar]
- Ji, S.Q.; Dix, S.R.; Aziz, A.A.; Sedelnikova, S.E.; Baker, P.J.; Rafferty, J.B.; Bullough, P.; Tzokov, S.; Agirre, J.; Li, F.-L.; et al. The molecular basis of endolytic activity of a multidomain alginate lyase from Defluviitalea phaphyphila, a representative of a new lyase family, PL39. J. Biol. Chem. 2019, 294, 18077–18091. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor sp SB155-2 Thriving at Deep-sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [Green Version]
- Yagi, H.; Fujise, A.; Itabashi, N.; Ohshiro, T. Purification and characterization of a novel alginate lyase from the marine bacterium Cobetia sp NAP1 isolated from brown algae. Biosci. Biotechnol. Biochem. 2016, 80, 2338–2346. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Li, N.N.; Yang, S.X.; Yu, Y.; Han, Z.L.; Li, L.; Mou, H. Study on expression and action mode of recombinant alginate lyases based on conserved domains reconstruction. Appl. Microbiol. Biotechnol. 2019, 103, 807–817. [Google Scholar] [CrossRef]
- Yin, C.; Sun, J.; Wang, H.; Yu, W.; Han, F. Identification and Characterization of a New Cold-Adapted and Alkaline Alginate Lyase TsAly7A from Thalassomonas sp. LD5 Produces Alginate Oligosaccharides with High Degree of Polymerization. Mar. Drugs 2022, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Dong, S.; Xu, F.; Dong, F.; Li, P.Y.; Zhang, X.Y.; Zhou, B.-C.; Zhang, Y.-Z.; Xie, B.-B. Characterization of a New Cold-Adapted and Salt-Activated Polysaccharide Lyase Family 7 Alginate Lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Wang, Q.; Lu, M.; Xu, C.; Li, F.; Zhang, R.; Liao, W.; Huang, S. AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity. Mar. Drugs 2018, 16, 120. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Li, Q.; Zhu, B.; Ni, F.; Sun, Y.; Yao, Z. Effects of module truncation on biochemical characteristics and products distribution of a new alginate lyase with two catalytic modules. Glycobiology 2019, 29, 876–884. [Google Scholar] [CrossRef]
- Cao, S.S.; Li, L.; Zhu, B.W.; Yao, Z. Biochemical Characterization and Elucidation of the Hybrid Action Mode of a New Psychrophilic and Cold-Tolerant Alginate Lyase for Efficient Preparation of Alginate Oligosaccharides. Mar. Drugs 2022, 20, 506. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Ni, F.; Sun, Y.; Ning, L.; Yao, Z. Elucidation of degrading pattern and substrate recognition of a novel bifunctional alginate lyase from Flammeovirga sp. NJ-04 and its use for preparation alginate oligosaccharides. Biotechnol. Biofuels 2019, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Hu, F.; Wang, M.; Zhu, B.; Ni, F.; Yao, Z. Elucidation of degradation pattern and immobilization of a novel alginate lyase for preparation of alginate oligosaccharides. Int. J. Biol. Macromol. 2020, 146, 579–587. [Google Scholar] [CrossRef]
- Yang, J.; Cui, D.D.; Chen, D.W.; Chen, W.K.; Ma, S.; Shen, H. Purification and Characterization of a Novel Endolytic Alginate Lyase from Microbulbifer sp. SH-1 and Its Agricultural Application. Mar. Drugs 2020, 18, 184. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Zhu, B.; Li, Q.; Yin, H.; Sun, Y.; Yao, Z.; Ming, D. Elucidation of a Unique Pattern and the Role of Carbohydrate Binding Module of an Alginate Lyase. Mar. Drugs 2019, 18, 32. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Yi, Y.J.; Chen, Z.; Wang, B.X.; Li, X.H.; Zhou, Y.X. Characterization of a New Biofunctional, Exolytic Alginate Lyase from Tamlana sp. s12 with High Catalytic Activity and Cold-Adapted Features. Mar. Drugs 2021, 19, 191. [Google Scholar] [CrossRef]
- Zhu, B.; Sun, Y.; Ni, F.; Ning, L.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef]
- Chen, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Sun, Y. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Hu, F.; Yuan, H.; Sun, Y.; Yao, Z. Biochemical Characterization and Degradation Pattern of a Unique pH-Stable PolyM-Specific Alginate Lyase from Newly Isolated Serratia marcescens NJ-07. Mar. Drugs 2018, 16, 129. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, L.; Hao, J.; Xing, M.; Sun, J.; Sun, M. Purification and Characterization of a New Alginate Lyase from Marine Bacterium Vibrio sp. SY08. Mar. Drugs 2016, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; An, D.; Jiao, C.; Xiao, Q.; Weng, H.F.; Yang, Q.M.; Xiao, A.F. Cloning, expression, and characterization of a new pH- and heat-stable alginate lyase from Pseudoalteromonas carrageenovora ASY5. J. Food Biochem. 2019, 43, e12886. [Google Scholar] [CrossRef]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and Characterization of a Novel Oligoalginate Lyase from a Newly Isolated Bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef]
- Itoh, T.; Nakagawa, E.; Yoda, M.; Nakaichi, A.; Hibi, T.; Kimoto, H. Structural and biochemical characterisation of a novel alginate lyase from Paenibacillus sp. str. FPU-7. Sci. Rep. 2019, 9, 14870. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Yu, Y.; Yang, S.; Shi, X.; Mou, H.; Li, L. Expression and Characterization of a New PolyG-Specific Alginate Lyase From Marine Bacterium Microbulbifer sp. Q7. Front. Microbiol. 2018, 9, 2894. [Google Scholar] [CrossRef]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an alginate lyase, FlAlyA, from Flavobacterium sp. strain UMI-01 and its expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Sodium Alginate | PolyM | PolyG |
|---|---|---|---|
| Activity (U/mg) | 37,315.08 | 28,814.31 | 21,329.21 |
| Km(mM) | 15.6738 | 7.3264 | 5.3961 |
| Vmax (mol/s) | 0.3285 | 0.5446 | 0.0497 |
| kcat (s−1) | 141.69 | 2323.78 | 212.07 |
| kcat/Km (s−1/mM) | 89.4288 | 317.1783 | 39.3006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Cao, S.; Zhu, B.; Yao, Z.; Zhu, B.; Qin, Y.; Jiang, J. Efficient Degradation of Alginate and Preparation of Alginate Oligosaccharides by a Novel Biofunctional Alginate Lyase with High Activity and Excellent Thermophilic Features. Mar. Drugs 2023, 21, 180. https://doi.org/10.3390/md21030180
Li L, Cao S, Zhu B, Yao Z, Zhu B, Qin Y, Jiang J. Efficient Degradation of Alginate and Preparation of Alginate Oligosaccharides by a Novel Biofunctional Alginate Lyase with High Activity and Excellent Thermophilic Features. Marine Drugs. 2023; 21(3):180. https://doi.org/10.3390/md21030180
Chicago/Turabian StyleLi, Li, Shengsheng Cao, Benwei Zhu, Zhong Yao, Bo Zhu, Yimin Qin, and Jinju Jiang. 2023. "Efficient Degradation of Alginate and Preparation of Alginate Oligosaccharides by a Novel Biofunctional Alginate Lyase with High Activity and Excellent Thermophilic Features" Marine Drugs 21, no. 3: 180. https://doi.org/10.3390/md21030180
APA StyleLi, L., Cao, S., Zhu, B., Yao, Z., Zhu, B., Qin, Y., & Jiang, J. (2023). Efficient Degradation of Alginate and Preparation of Alginate Oligosaccharides by a Novel Biofunctional Alginate Lyase with High Activity and Excellent Thermophilic Features. Marine Drugs, 21(3), 180. https://doi.org/10.3390/md21030180