

Anti-Inflammatory Activity of Fucan from Spatoglossum schröederi in a Murine Model of Generalized Inflammation Induced by Zymosan

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Fucan from S. schröederi Can Inhibit Leukocyte Migration to Peritoneal Cavity in Zymosan-Induced Peritonits Murine Model
2.2. Fucan from S. schröederi Can Attenuate Systemic Toxicity Signs in ZIGI Murine Model
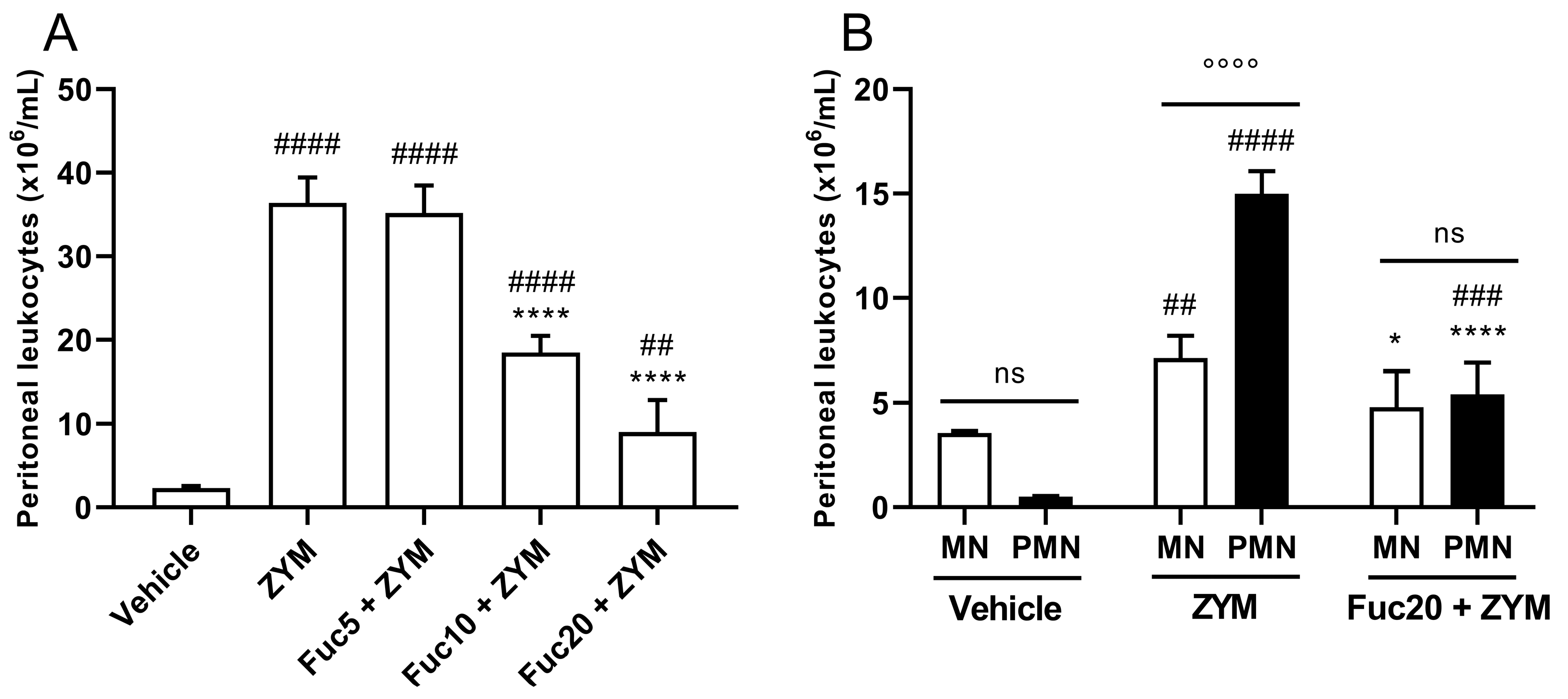
2.3. Fucan from S. schröederi Can Inhibit Peritoneal Leukocyte Migration in ZIGI Murine Model
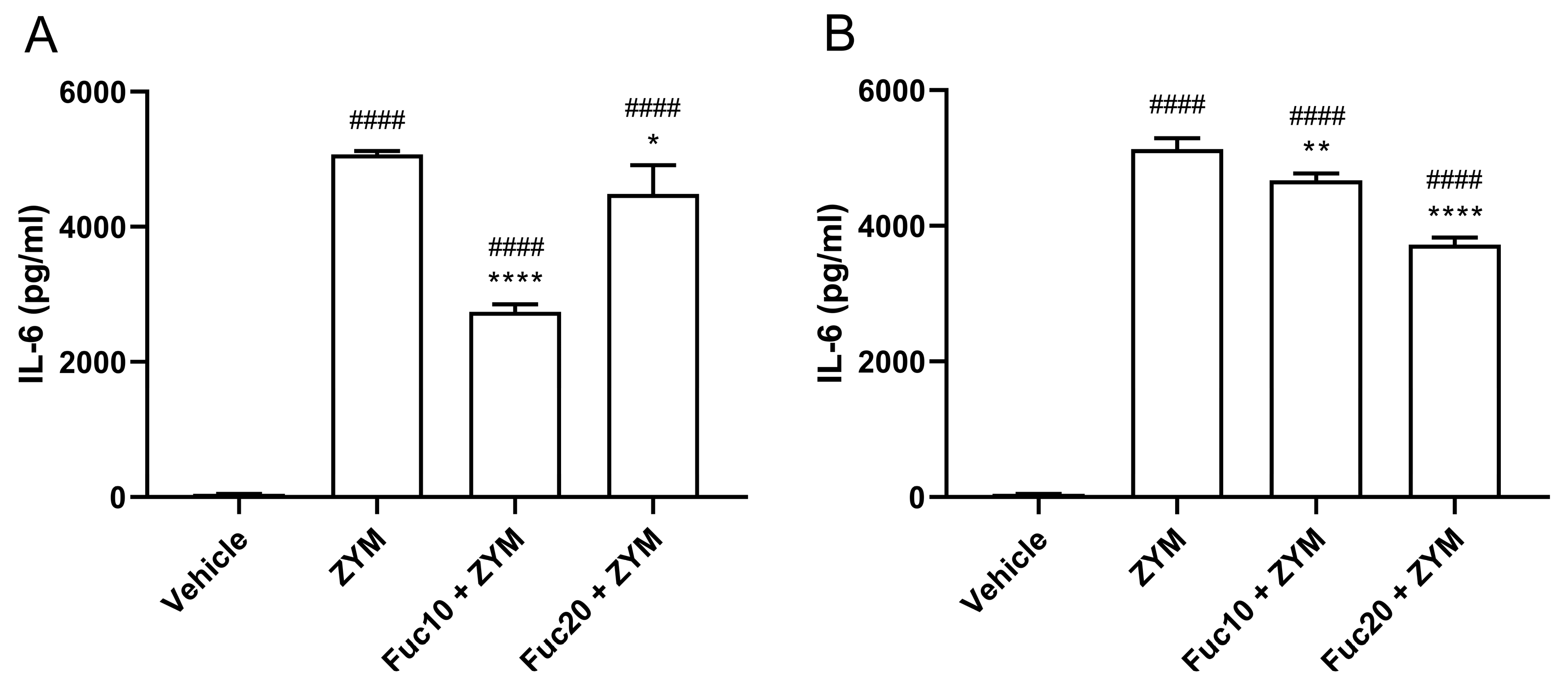
2.4. Fucan from S. schröederi Can Reduce IL-6 Levels in ZIGI Murine Model
2.5. Fucan of S. schröederi Show Protective Hepatic Effect, Attenuating Liver Damage in ZIGI Murine Model
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Fucan Obtaining
4.3. Zymosan-Induced Peritonitis
4.4. Zymosan-Induced Generalized Inflammation Model (ZIGI)
4.5. IL-6 and Hepatic Transaminase (ALT/AST) Determination
4.6. ZIGI Systemic Toxicity Evaluation
4.7. Histopathological Analysis
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kloareg, B.; Quatrano, R.S. Structure of the Cell Walls of Marine Algae and Ecophysiological Functions of the Matrix Polysaccharides. Oceanogr. Mar. Biol. Annu. Rev. 1988, 26, 259–315. [Google Scholar]
- Mazepa, E.; Biscaia, S.M.P.; Bellan, D.d.L.; Trindade, E.d.S.; Simas, F.F. Structural Characteristics of Native and Chemically Sulfated Polysaccharides from Seaweed and Their Antimelanoma Effects. Carbohydr. Polym. 2022, 289, 119436. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. The Fucoidans from Brown Algae of Far-Eastern Seas: Anti-Tumor Activity and Structure-Function Relationship. Food Chem. 2013, 141, 1211–1217. [Google Scholar] [CrossRef]
- Lima, A.G.F.; Mignone, V.W.; Vardiero, F.; Kozlowski, E.O.; Fernandes, L.R.; Motta, J.M.; Pavão, M.S.G.; Figueiredo, C.C.; Mourão, P.A.S.; Morandi, V. Direct Anti-Tumoral Effects of Sulfated Fucans Isolated from Echinoderms: A Possible Role of Neuropilin-1/Β1 Integrin Endocytosis and Focal Adhesion Kinase (FAK) Degradation. Glycobiology 2023, cwad044. [Google Scholar] [CrossRef]
- Dore, C.M.P.G.; Alves, M.G.d.C.F.; Will, L.S.E.P.; Costa, T.G.; Sabry, D.A.; de Souza Rêgo, L.A.R.; Accardo, C.M.; Rocha, H.A.; Filgueira, L.G.A.; Leite, E.L. A Sulfated Polysaccharide, Fucans, Isolated from Brown Algae Sargassum Vulgare with Anticoagulant, Antithrombotic, Antioxidant and Anti-Inflammatory Effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Liu, J.; Lin, L.; Sun, H.; Yang, W.; Cai, Y.; Gao, N.; Zhou, L.; Qin, H.; et al. A Regular Fucan Sulfate from Stichopus Herrmanni and Its Peroxide Depolymerization: Structure and Anticoagulant Activity. Carbohydr. Polym. 2021, 256, 117513. [Google Scholar] [CrossRef]
- Zhu, Q.; Lin, L.; Zhao, M. Sulfated Fucan/Fucosylated Chondroitin Sulfate-Dominated Polysaccharide Fraction from Low-Edible-Value Sea Cucumber Ameliorates Type 2 Diabetes in Rats: New Prospects for Sea Cucumber Polysaccharide Based-Hypoglycemic Functional Food. Int. J. Biol. Macromol. 2020, 159, 34–45. [Google Scholar] [CrossRef]
- Ahmadi, A.; Moghadamtousi, S.Z.; Abubakar, S.; Zandi, K. Antiviral Potential of Algae Polysaccharides Isolated from Marine Sources: A Review. BioMed Res. Int. 2015, 2015, 825203. [Google Scholar] [CrossRef]
- Leal, D.; Mansilla, A.; Matsuhiro, B.; Moncada-Basualto, M.; Lapier, M.; Maya, J.D.; Olea-Azar, C.; De Borggraeve, W.M. Chemical Structure and Biological Properties of Sulfated Fucan from the Sequential Extraction of SubAntarctic Lessonia Sp (Phaeophyceae). Carbohydr. Polym. 2018, 199, 304–313. [Google Scholar] [CrossRef]
- Gao, L.; Xu, C.; Tao, X.; Zuo, Z.; Ning, Z.; Wang, L.; Gao, N.; Zhao, J. Structure Elucidation of Fucan Sulfate from Sea Cucumber Holothuria Fuscopunctata through a Bottom-Up Strategy and the Antioxidant Activity Analysis. Int. J. Mol. Sci. 2022, 23, 4488. [Google Scholar] [CrossRef]
- Feng, G.; Laijin, S.; Chen, S.; Teng, W.; Dejian, Z.; Yin, C.; Shoudong, G. In Vitro and in Vivo Immunoregulatory Activity of Sulfated Fucan from the Sea Cucumber A. Leucoprocta. Int. J. Biol. Macromol. 2021, 187, 931–938. [Google Scholar] [CrossRef]
- Chen, X.; Ni, L.; Fu, X.; Wang, L.; Duan, D.; Huang, L.; Xu, J.; Gao, X. Molecular Mechanism of Anti-Inflammatory Activities of a Novel Sulfated Galactofucan from Saccharina Japonica. Mar. Drugs 2021, 19, 430. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Shikov, A.N. In Vitro Anti-Inflammatory Activities of Fucoidans from Five Species of Brown Seaweeds. Mar. Drugs 2022, 20, 606. [Google Scholar] [CrossRef]
- Wang, S.; Ni, L.; Fu, X.; Duan, D.; Xu, J.; Gao, X. A Sulfated Polysaccharide from Saccharina Japonica Suppresses LPS-Induced Inflammation Both in a Macrophage Cell Model via Blocking MAPK/NF-ΚB Signal Pathways In Vitro and a Zebrafish Model of Embryos and Larvae In Vivo. Mar. Drugs 2020, 18, 593. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, A.; Subramanian, P.; Manoharan, V.; Muthu, T.; Periyannan, R.; Thangapandi, M.; Ponnuchamy, K.; Pandi, B.; Marimuthu, P.N. Phyto-Mediated Synthesis of Silver Nanoparticles Using Fucoidan Isolated from Spatoglossum Asperum and Assessment of Antibacterial Activities. J. Photochem. Photobiol. B 2018, 185, 117–125. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. In Vitro Antioxidant and Antibacterial Activity of Sulfated Polysaccharides Isolated from Spatoglossum Asperum. Carbohydr. Polym. 2017, 170, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Tanna, B.; Choudhary, B.; Mishra, A.; Chauhan, O.P.; Patel, M.K.; Shokralla, S.; El-Abedin, T.K.Z.; Elansary, H.O.; Mahmoud, E.A. Antioxidant, Scavenging, Reducing, and Anti-Proliferative Activities of Selected Tropical Brown Seaweeds Confirm the Nutraceutical Potential of Spatoglossum Asperum. Foods 2021, 10, 2482. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Souza, I.; Pessatti, J.B.K.; da Silva, L.R.; Bellan, D.d.L.; de Souza, I.R.; Cestari, M.M.; de Assis, H.C.S.; Rocha, H.A.O.; Simas, F.F.; Trindade, E.d.S.; et al. Protective Potential of Sulfated Polysaccharides from Tropical Seaweeds against Alkylating- and Oxidizing-Induced Genotoxicity. Int. J. Biol. Macromol. 2022, 211, 524–534. [Google Scholar] [CrossRef]
- Bellan, D.L.; Bini, I.H.; Santi, F.C.; Rossi, G.R.; Biscaia, S.M.P.; Maximo, A.I.; Reis, M.B.; Simas, F.F.; Oliveira, C.C.; Winnischofer, S.M.B.; et al. Fucoidan from Spatoglossum Schröederi Promotes B16-F10 Malignancy Features Modulation and Antimelanoma in Vivo Activities. Algal Res. 2023, 72, 103134. [Google Scholar] [CrossRef]
- Rocha, H.A.O.; Franco, C.R.C.; Trindade, E.S.; Carvalho, L.C.M.; Veiga, S.S.; Leite, E.L.; Dietrich, C.P.; Nader, H.B. A Fucan from the Brown Seaweed Spatoglossum Schröederi Inhibits Chinese Hamster Ovary Cell Adhesion to Several Extracellular Matrix Proteins. Braz. J. Med. Biol. Res. 2001, 34, 621–626. [Google Scholar] [CrossRef]
- Rocha, H.A.d.O.; Bezerra, L.C.d.M.; de Albuquerque, I.R.; Costa, L.S.; Guerra, C.M.; de Abreu, L.D.; Nader, H.B.; Leite, E.L. A Xylogalactofucan from the Brown Seaweed Spatoglossum schröederi Stimulates the Synthesis of an Antithrombotic Heparan Sulfate from Endothelial Cells. Planta Med. 2005, 71, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Farias, W.R.L.; Lima, P.C.W.C.; Rodrigues, N.V.F.C.; Siqueira, R.C.L.; Amorim, R.M.F.; Pereira, M.G.; Assreuy, A.M.S. A Novel Antinociceptive Sulphated Polysaccharide of the Brown Marine Alga Spatoglossum schroederi. Nat. Prod. Commun. 2011, 6, 1934578X1100600. [Google Scholar] [CrossRef]
- Menezes, M.M.; Nobre, L.T.D.B.; Rossi, G.R.; Almeida-Lima, J.; Melo-Silveira, R.F.; Franco, C.R.C.; Trindade, E.S.; Nader, H.B.; Rocha, H.A.O. A Low-Molecular-Weight Galactofucan from the Seaweed, Spatoglossum schröederi, Binds Fibronectin and Inhibits Capillary-like Tube Formation in Vitro. Int. J. Biol. Macromol. 2018, 111, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.O.; Oliveira, J.W.d.F.; Moreno, C.J.G.; de Medeiros, M.J.C.; Fernandes-Negreiros, M.M.; Souza, F.R.M.; Pontes, D.L.; Silva, M.S.; Rocha, H.A.O. Silver Nanoparticles Containing Fucoidan Synthesized by Green Method Have Anti-Trypanosoma Cruzi Activity. Nanomaterials 2022, 12, 2059. [Google Scholar] [CrossRef] [PubMed]
- Mandalka, A.; Cavalcanti, M.I.L.G.; Harb, T.B.; Toyota Fujii, M.; Eisner, P.; Schweiggert-Weisz, U.; Chow, F. Nutritional Composition of Beach-Cast Marine Algae from the Brazilian Coast: Added Value for Algal Biomass Considered as Waste. Foods 2022, 11, 1201. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801. [Google Scholar] [CrossRef]
- van der Poll, T.; Shankar-Hari, M.; Wiersinga, W.J. The Immunology of Sepsis. Immunity 2021, 54, 2450–2464. [Google Scholar] [CrossRef]
- Arina, P.; Singer, M. Pathophysiology of Sepsis. Curr. Opin. Anaesthesiol. 2021, 34, 77–84. [Google Scholar] [CrossRef]
- Font, M.D.; Thyagarajan, B.; Khanna, A.K. Sepsis and Septic Shock—Basics of Diagnosis, Pathophysiology and Clinical Decision Making. Med. Clin. N. Am. 2020, 104, 573–585. [Google Scholar] [CrossRef]
- Pool, R.; Gomez, H.; Kellum, J.A. Mechanisms of Organ Dysfunction in Sepsis. Crit. Care Clin. 2018, 34, 63–80. [Google Scholar] [CrossRef]
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and Septic Shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Gavelli, F.; Castello, L.M.; Avanzi, G.C. Management of Sepsis and Septic Shock in the Emergency Department. Intern. Emerg. Med. 2021, 16, 1649–1661. [Google Scholar] [CrossRef]
- Perner, A.; Gordon, A.C.; Angus, D.C.; Lamontagne, F.; Machado, F.; Russell, J.A.; Timsit, J.-F.; Marshall, J.C.; Myburgh, J.; Shankar-Hari, M.; et al. The Intensive Care Medicine Research Agenda on Septic Shock. Intensiv. Care Med. 2017, 43, 1294–1305. [Google Scholar] [CrossRef]
- Thompson, K.; Venkatesh, B.; Finfer, S. Sepsis and Septic Shock: Current Approaches to Management. Intern. Med. J. 2019, 49, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, Regional, and National Sepsis Incidence and Mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Goris, R.J.A.; Boekholtz, W.K.F.; van Bebber, I.P.T.; Nuytinck, J.K.S.; Schillings, P.H.M. Multiple-Organ Failure and Sepsis Without Bacteria: An Experimental Model. Arch. Surg. 1986, 121, 897. [Google Scholar] [CrossRef]
- Volman, T.J.H.; Hendriks, T.; Goris, R.J.A. Zymosan-Induced Generalized Inflammation: Experimental Studies into Mechanisms Leading to Multiple Organ Dysfunction Syndrome. Shock 2005, 23, 291–297. [Google Scholar] [CrossRef]
- Paola, R.; Mazzon, E.; Muià, C.; Crisafulli, C.; Genovese, T.; Bella, P.; Esposito, E.; Menegazzi, M.; Meli, R.; Suzuki, H.; et al. Green Tea Polyphenol Extract Attenuates Zymosan-Induced Non-Septic Shock in Mice. Shock 2006, 26, 402–409. [Google Scholar] [CrossRef]
- Malleo, G.; Mazzon, E.; Genovese, T.; Di Paola, R.; Caminiti, R.; Esposito, E.; Bramanti, P.; Cuzzocrea, S. Absence of Endogenous Interleukin-10 Enhanced Organ Dysfunction and Mortality Associated to Zymosan-Induced Multiple Organ Dysfunction Syndrome. Cytokine 2008, 41, 136–143. [Google Scholar] [CrossRef]
- Galuppo, M.; Di Paola, R.; Mazzon, E.; Genovese, T.; Crisafulli, C.; Paterniti, I.; Cuzzocrea, E.; Bramanti, P.; Kapoor, A.; Thiemermann, C.; et al. Role of PPAR-δ in the Development of Zymosan-Induced Multiple Organ Failure: An Experiment Mice Study. J. Inflamm. 2010, 7, 12. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, J.; Pang, Q.; Song, S.; Miao, R.; Chen, W.; Zhou, Y.; Liu, C. The Protective Role of Curcumin in Zymosan-Induced Multiple Organ Dysfunction Syndrome in Mice. Shock 2016, 45, 209–219. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, Y.; Wang, W.; Jia, L.; Zhang, J. Characterization and Hepatoprotections of Ganoderma Lucidum Polysaccharides against Multiple Organ Dysfunction Syndrome in Mice. Oxidative Med. Cell. Longev. 2021, 2021, 9703682. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Feng, Y.; Zhang, M.; Song, X.; Jia, L. Protective Effects of Sulfated Polysaccharides from Lentinula edodes on the Lung and Liver of MODS Mice. Food Funct. 2021, 12, 6389–6402. [Google Scholar] [CrossRef]
- Fitton, J.H.; Stringer, D.N.; Park, A.Y.; Karpiniec, S.S. Therapies from Fucoidan: New Developments. Mar. Drugs 2019, 17, 571. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Y.; Zhang, Y.; Yang, Y.; Wang, P.; Imre, B.; Wong, A.C.Y.; Hsieh, Y.S.Y.; Wang, D. Brown Algae Carbohydrates: Structures, Pharmaceutical Properties, and Research Challenges. Mar. Drugs 2021, 19, 620. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Herath, K.H.I.N.M.; Yang, H.-W.; Choi, C.S.; Jeon, Y.-J. Anti-Inflammatory Mechanisms of Fucoidans to Treat Inflammatory Diseases: A Review. Mar. Drugs 2021, 19, 678. [Google Scholar] [CrossRef]
- Mensah, E.O.; Kanwugu, O.N.; Panda, P.K.; Adadi, P. Marine Fucoidans: Structural, Extraction, Biological Activities and Their Applications in the Food Industry. Food Hydrocoll. 2023, 142, 108784. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Manikandan, R.; Parimalanandhini, D.; Mahalakshmi, K.; Beulaja, M.; Arumugam, M.; Janarthanan, S.; Palanisamy, S.; You, S.; Prabhu, N.M. Studies on Isolation, Characterization of Fucoidan from Brown Algae Turbinaria Decurrens and Evaluation of It’s in Vivo and in Vitro Anti-Inflammatory Activities. Int. J. Biol. Macromol. 2020, 160, 1263–1276. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Delattre, C.; Molinié, R.; Petit, E.; Elboutachfaiti, R.; Nikolova, M.; Iliev, I.; Murdjeva, M.; et al. Structural Characterization and In Vivo Anti-Inflammatory Activity of Fucoidan from Cystoseira Crinita (Desf.) Borry. Mar. Drugs 2022, 20, 714. [Google Scholar] [CrossRef]
- Phull, A.R.; Kim, S.J. Fucoidan as Bio-Functional Molecule: Insights into the Anti-Inflammatory Potential and Associated Molecular Mechanisms. J. Funct. Foods 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, J.; Ge, K.; Tian, Q.; Zhao, P.; Guo, Y. Anti-Inflammatory Effect of Low Molecular Weight Fucoidan from Saccharina Japonica on Atherosclerosis in ApoE-Knockout Mice. Int. J. Biol. Macromol. 2018, 118, 365–374. [Google Scholar] [CrossRef]
- Fine, N.; Tasevski, N.; McCulloch, C.A.; Tenenbaum, H.C.; Glogauer, M. The Neutrophil: Constant Defender and First Responder. Front Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Marques, C.T.; Azevedo, T.C.G.d.; Nascimento, M.S.; Medeiros, V.P.; Alves, L.G.; Benevides, N.M.B.; Rocha, H.A.O.; Leite, E.L. Sulfated Fucans Extracted from Algae Padina Gymnospora Have Anti-Inflammatory Effect. Rev. Bras. Farm. 2012, 22, 115–122. [Google Scholar] [CrossRef]
- Yu, H.-H.; Chengchuan KO, E.; Chang, C.-L.; Yuan, K.; Wu, A.; Shan, Y.-S.; Wu, S.-Y. Fucoidan Inhibits Radiation-Induced Pneumonitis and Lung Fibrosis by Reducing Inflammatory Cytokine Expression in Lung Tissues. Mar. Drugs 2018, 16, 392. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Wang, L.; Fu, X.; Duan, D.; Jeon, Y.-J.; Xu, J.; Gao, X. In Vitro and in Vivo Anti-Inflammatory Activities of a Fucose-Rich Fucoidan Isolated from Saccharina Japonica. Int. J. Biol. Macromol. 2020, 156, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; Vaamonde-García, C.; Torres, M.D.; Buján, M.; Muíños, A.; Muiños, A.; Lamas-Vázquez, M.J.; Meijide-Faílde, R.; Blanco, F.J.; Domínguez, H. Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans. Pharmaceutics 2023, 15, 808. [Google Scholar] [CrossRef]
- Ahmad, T.; Ishaq, M.; Karpiniec, S.; Park, A.; Stringer, D.; Singh, N.; Ratanpaul, V.; Wolfswinkel, K.; Fitton, H.; Caruso, V.; et al. Oral Macrocystis Pyrifera Fucoidan Administration Exhibits Anti-Inflammatory and Antioxidant Properties and Improves DSS-Induced Colitis in C57BL/6J Mice. Pharmaceutics 2022, 14, 2383. [Google Scholar] [CrossRef]
- Kuznetsova, T.; Besednova, N.; Somova, L.; Plekhova, N. Fucoidan Extracted from Fucus Evanescens Prevents Endotoxin-Induced Damage in a Mouse Model of Endotoxemia. Mar. Drugs 2014, 12, 886–898. [Google Scholar] [CrossRef]
- Uciechowski, P.; Dempke, W.C.M. Interleukin-6: A Masterplayer in the Cytokine Network. Oncology 2020, 98, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Rivera, G.L.; Madera-Sandoval, R.L.; León-Pedroza, J.I.; Ferat-Osorio, E.; Salazar-Rios, E.; Hernández-Aceves, J.A.; Guadarrama-Aranda, U.; López-Macías, C.; Wong-Baeza, I.; Arriaga-Pizano, L.A. Increased TNF-α Production in Response to IL-6 in Patients with Systemic Inflammation without Infection. Clin. Exp. Immunol. 2022, 209, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Chen, D.; Zhou, R.; Zhao, X.; Ye, C.; Tao, H.; Sheng, W.; Wu, Y. Combination of C-reactive Protein, Procalcitonin, IL-6, IL-8, and IL-10 for Early Diagnosis of Hyperinflammatory State and Organ Dysfunction in Pediatric Sepsis. J. Clin. Lab. Anal. 2022, 36. [Google Scholar] [CrossRef] [PubMed]
- Shimazui, T.; Nakada, T.; Yazaki, M.; Mayumi, T.; Takasu, O.; Matsuda, K.; Sasaki, J.; Otsubo, H.; Teshima, Y.; Nabeta, M.; et al. Blood Interleukin-6 Levels Predict Multiple Organ Dysfunction in Critically III Patients. Shock 2021, 55, 790–795. [Google Scholar] [CrossRef]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Mar. Drugs 2021, 19, 702. [Google Scholar] [CrossRef]
- Dörschmann, P.; Seeba, C.; Thalenhorst, T.; Roider, J.; Klettner, A. Anti-Inflammatory Properties of Antiangiogenic Fucoidan in Retinal Pigment Epithelium Cells. Heliyon 2023, 9, e15202. [Google Scholar] [CrossRef]
- Wang, L.; Cui, Y.R.; Wang, K.; Fu, X.; Xu, J.; Gao, X.; Jeon, Y.-J. Anti-Inflammatory Effect of Fucoidan Isolated from Fermented Sargassum Fusiforme in in Vitro and in Vivo Models. Int. J. Biol. Macromol. 2022, 222, 2065–2071. [Google Scholar] [CrossRef]
- Jayasinghe, A.M.K.; Kirindage, K.G.I.S.; Fernando, I.P.S.; Kim, K.-N.; Oh, J.-Y.; Ahn, G. The Anti-Inflammatory Effect of Low Molecular Weight Fucoidan from Sargassum Siliquastrum in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages via Inhibiting NF-ΚB/MAPK Signaling Pathways. Mar. Drugs 2023, 21, 347. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and Tumor Progression: Signaling Pathways and Targeted Intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Zhong, L.; Simard, M.J.; Huot, J. Endothelial MicroRNAs Regulating the NF-κB Pathway and Cell Adhesion Molecules during Inflammation. FASEB J. 2018, 32, 4070–4084. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB System. WIREs Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, N.M.; Lee, H.-G.; Nagahawatta, D.P.; Jayawardhana, H.H.A.C.K.; Song, K.-M.; Choi, Y.-S.; Jeon, Y.-J.; Kang, M.-C. Fucoidan from Sargassum Autumnale Inhibits Potential Inflammatory Responses via NF-ΚB and MAPK Pathway Suppression in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Mar. Drugs 2023, 21, 374. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Chen, D.; Ye, Z.; Huang, Y.; Zhang, N.; Lui, E.M.K.; Xue, C.; Xiao, M. Fucoidan Isolated from Saccharina Japonica Inhibits LPS-Induced Inflammation in Macrophages via Blocking NF-ΚB, MAPK and JAK-STAT Pathways. Mar. Drugs 2020, 18, 328. [Google Scholar] [CrossRef] [PubMed]
- Nagahawatta, D.P.; Liyanage, N.M.; Jayawardhana, H.H.A.C.K.; Lee, H.-G.; Jayawardena, T.; Jeon, Y.-J. Anti-Fine Dust Effect of Fucoidan Extracted from Ecklonia Maxima Leaves in Macrophages via Inhibiting Inflammatory Signaling Pathways. Mar. Drugs 2022, 20, 413. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Nagahawatta, D.P.; Lee, H.-G.; Lu, Y.-A.; Vaas, A.P.J.P.; Abeytunga, D.T.U.; Nanayakkara, C.M.; Lee, D.-S.; Jeon, Y.-J. Anti-Inflammatory Effects of Sulfated Polysaccharide from Sargassum Swartzii in Macrophages via Blocking TLR/NF-Κb Signal Transduction. Mar. Drugs 2020, 18, 601. [Google Scholar] [CrossRef] [PubMed]
- Kiselevskiy, M.V.; Anisimova, N.Y.; Bilan, M.I.; Usov, A.I.; Ustyuzhanina, N.E.; Petkevich, A.A.; Shubina, I.Z.; Morozevich, G.E.; Nifantiev, N.E. Prospects for the Use of Marine Sulfated Fucose-Rich Polysaccharides in Treatment and Prevention of COVID-19 and Post-COVID-19 Syndrome. Russ. J. Bioorg. Chem. 2022, 48, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Potential Beneficial Actions of Fucoidan in Brain and Liver Injury, Disease, and Intoxication—Potential Implication of Sirtuins. Mar. Drugs 2020, 18, 242. [Google Scholar] [CrossRef]
- Lim, J.; Lee, S.; Kim, T.; Jang, S.-A.; Kang, S.; Koo, H.; Sohn, E.; Bak, J.; Namkoong, S.; Kim, H.; et al. Fucoidan from Fucus Vesiculosus Protects against Alcohol-Induced Liver Damage by Modulating Inflammatory Mediators in Mice and HepG2 Cells. Mar. Drugs 2015, 13, 1051–1067. [Google Scholar] [CrossRef]
- Meenakshi, S.; Umayaparvathi, S.; Saravanan, R.; Manivasagam, T.; Balasubramanian, T. Hepatoprotective Effect of Fucoidan Isolated from the Seaweed Turbinaria Decurrens in Ethanol Intoxicated Rats. Int. J. Biol. Macromol. 2014, 67, 367–372. [Google Scholar] [CrossRef]
- Kim, T.S.; Choi, D.H. Liver Dysfunction in Sepsis. Korean J. Gastroenterol. 2020, 75, 182–187. [Google Scholar] [CrossRef]
- de Garibay, A.P.R.; Kortgen, A.; Leonhardt, J.; Zipprich, A.; Bauer, M. Critical Care Hepatology: Definitions, Incidence, Prognosis and Role of Liver Failure in Critically III Patients. Crit. Care 2022, 26, 289. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.K.A.; Souza, C.R.d.M.; Silva, H.M.D.; Jales, J.T.; Gomez, L.A.d.S.; da Silveira, E.J.D.; Rocha, H.A.O.; Souto, J.T. Anti-Inflammatory Activity of Fucan from Spatoglossum schröederi in a Murine Model of Generalized Inflammation Induced by Zymosan. Mar. Drugs 2023, 21, 557. https://doi.org/10.3390/md21110557
Silva AKA, Souza CRdM, Silva HMD, Jales JT, Gomez LAdS, da Silveira EJD, Rocha HAO, Souto JT. Anti-Inflammatory Activity of Fucan from Spatoglossum schröederi in a Murine Model of Generalized Inflammation Induced by Zymosan. Marine Drugs. 2023; 21(11):557. https://doi.org/10.3390/md21110557
Chicago/Turabian StyleSilva, Ana Katarina Andrade, Cássio Ricardo de Medeiros Souza, Hylarina Montenegro Diniz Silva, Jéssica Teixeira Jales, Lucas Alves de Souza Gomez, Ericka Janine Dantas da Silveira, Hugo Alexandre Oliveira Rocha, and Janeusa Trindade Souto. 2023. "Anti-Inflammatory Activity of Fucan from Spatoglossum schröederi in a Murine Model of Generalized Inflammation Induced by Zymosan" Marine Drugs 21, no. 11: 557. https://doi.org/10.3390/md21110557
APA StyleSilva, A. K. A., Souza, C. R. d. M., Silva, H. M. D., Jales, J. T., Gomez, L. A. d. S., da Silveira, E. J. D., Rocha, H. A. O., & Souto, J. T. (2023). Anti-Inflammatory Activity of Fucan from Spatoglossum schröederi in a Murine Model of Generalized Inflammation Induced by Zymosan. Marine Drugs, 21(11), 557. https://doi.org/10.3390/md21110557