
Effect of Temperature and Light Intensity on the Polar Lipidome of Endophytic Brown Algae Streblonema corymbiferum and Streblonema sp. In Vitro


Abstract
:1. Introduction
2. Results
2.1. The Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
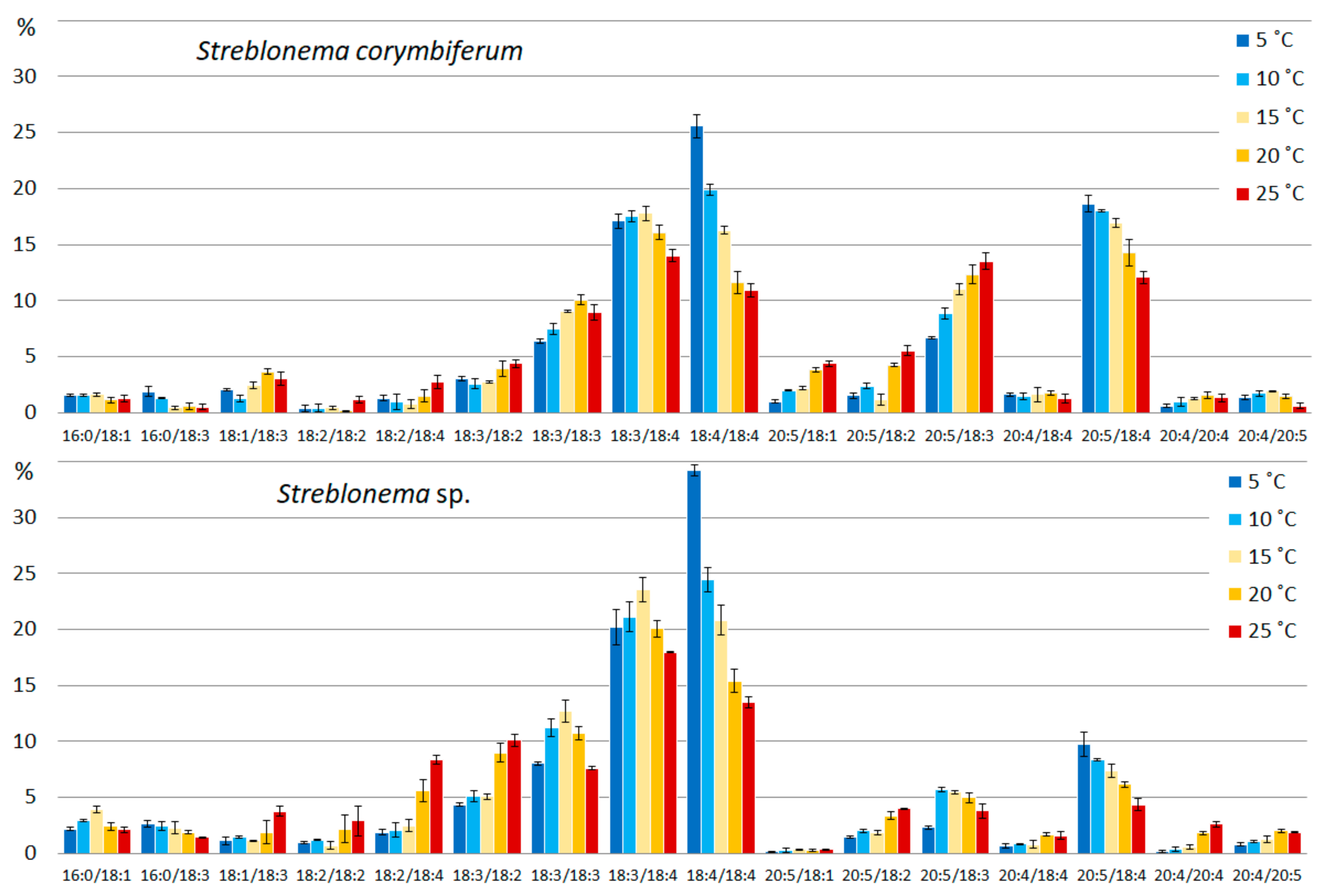
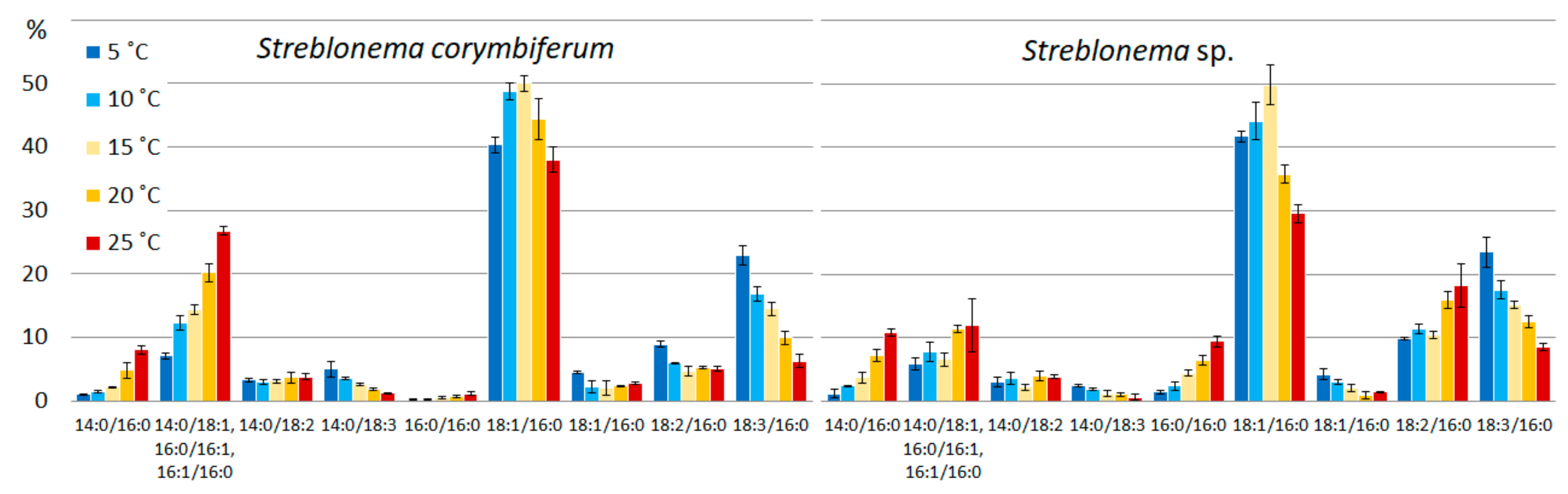
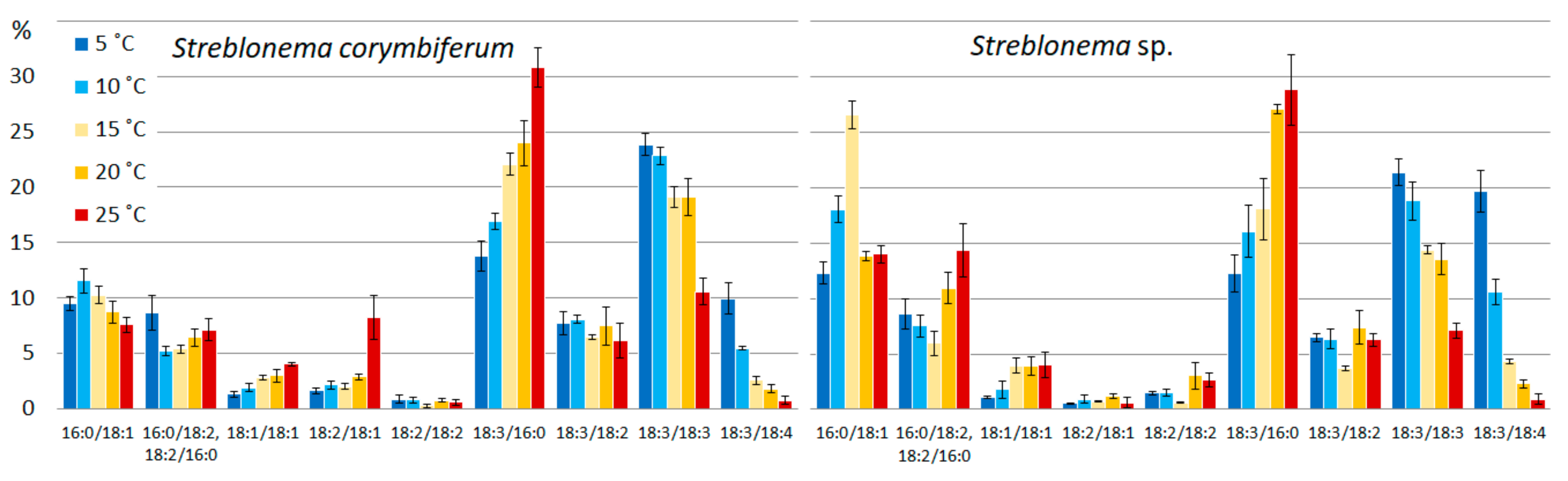
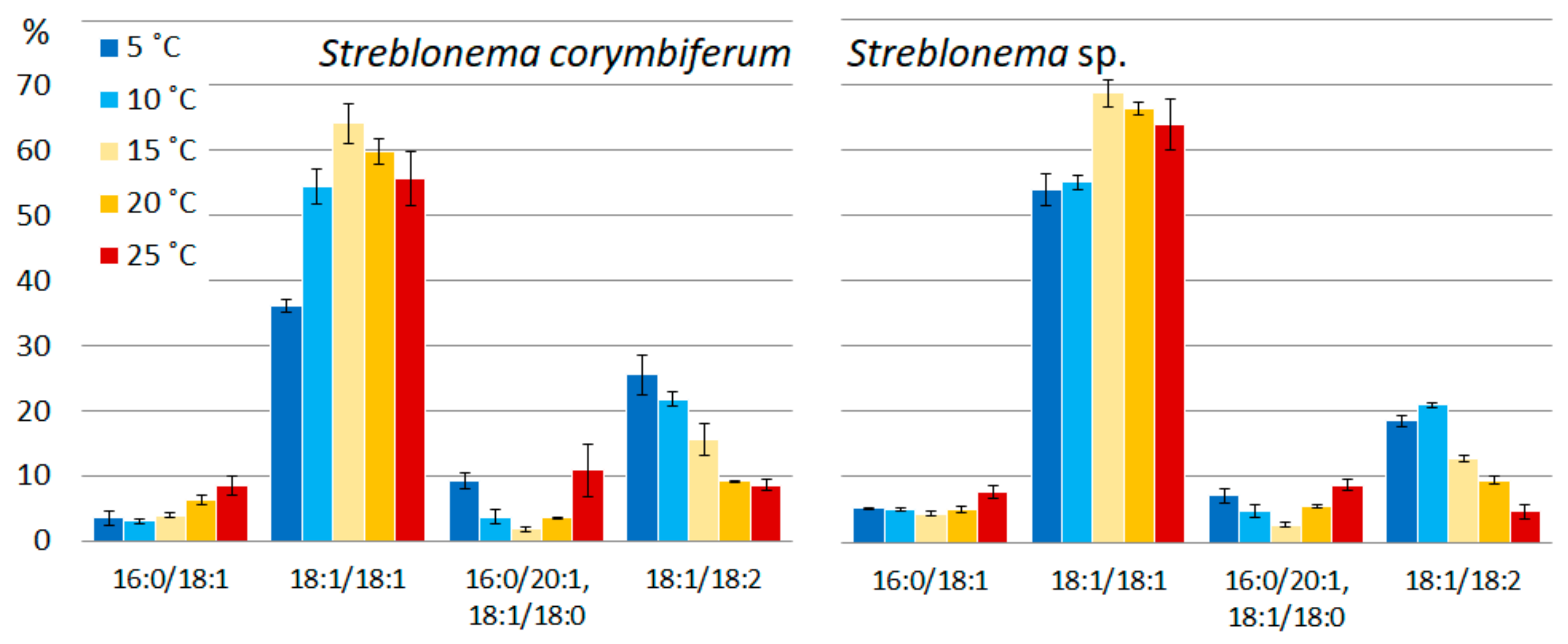
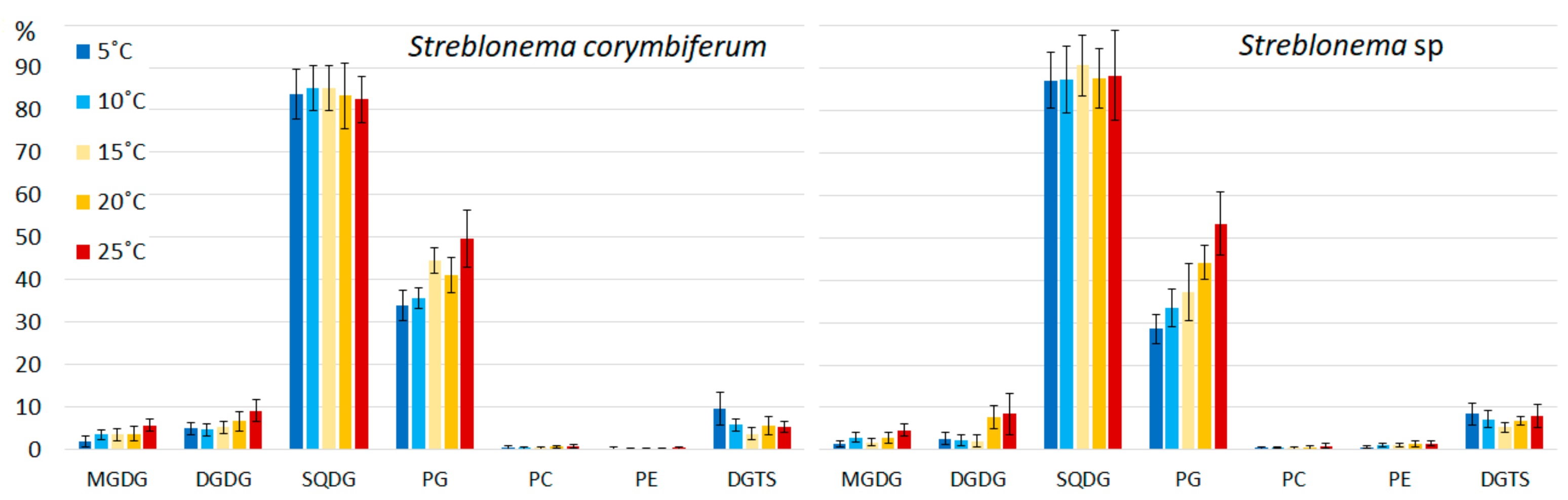
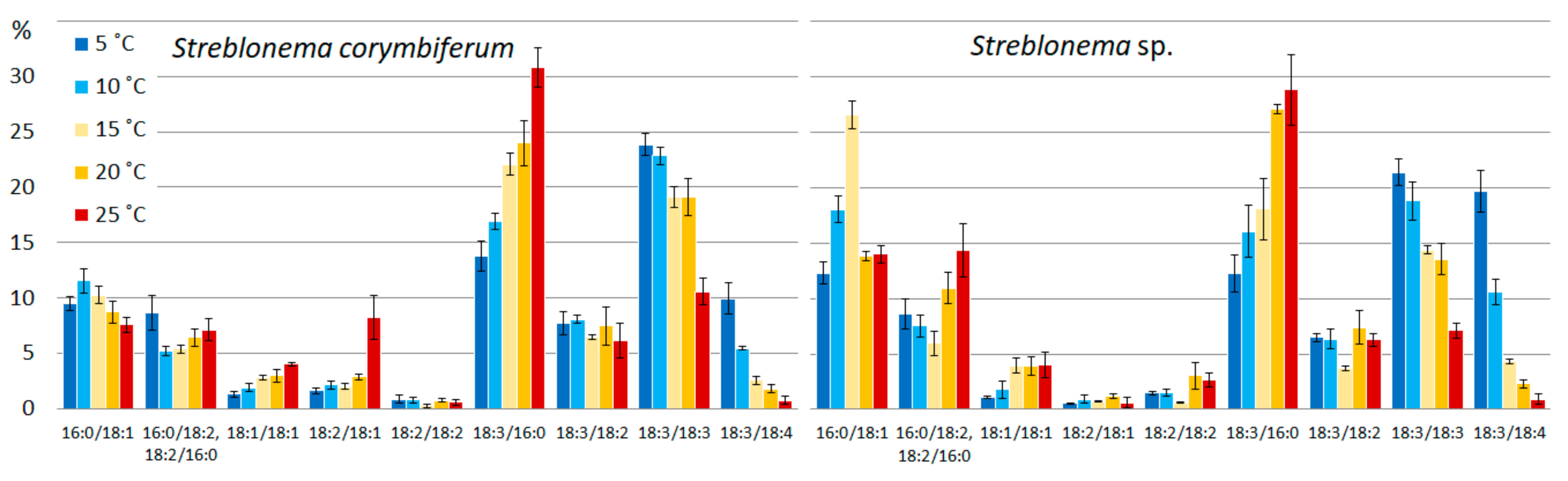
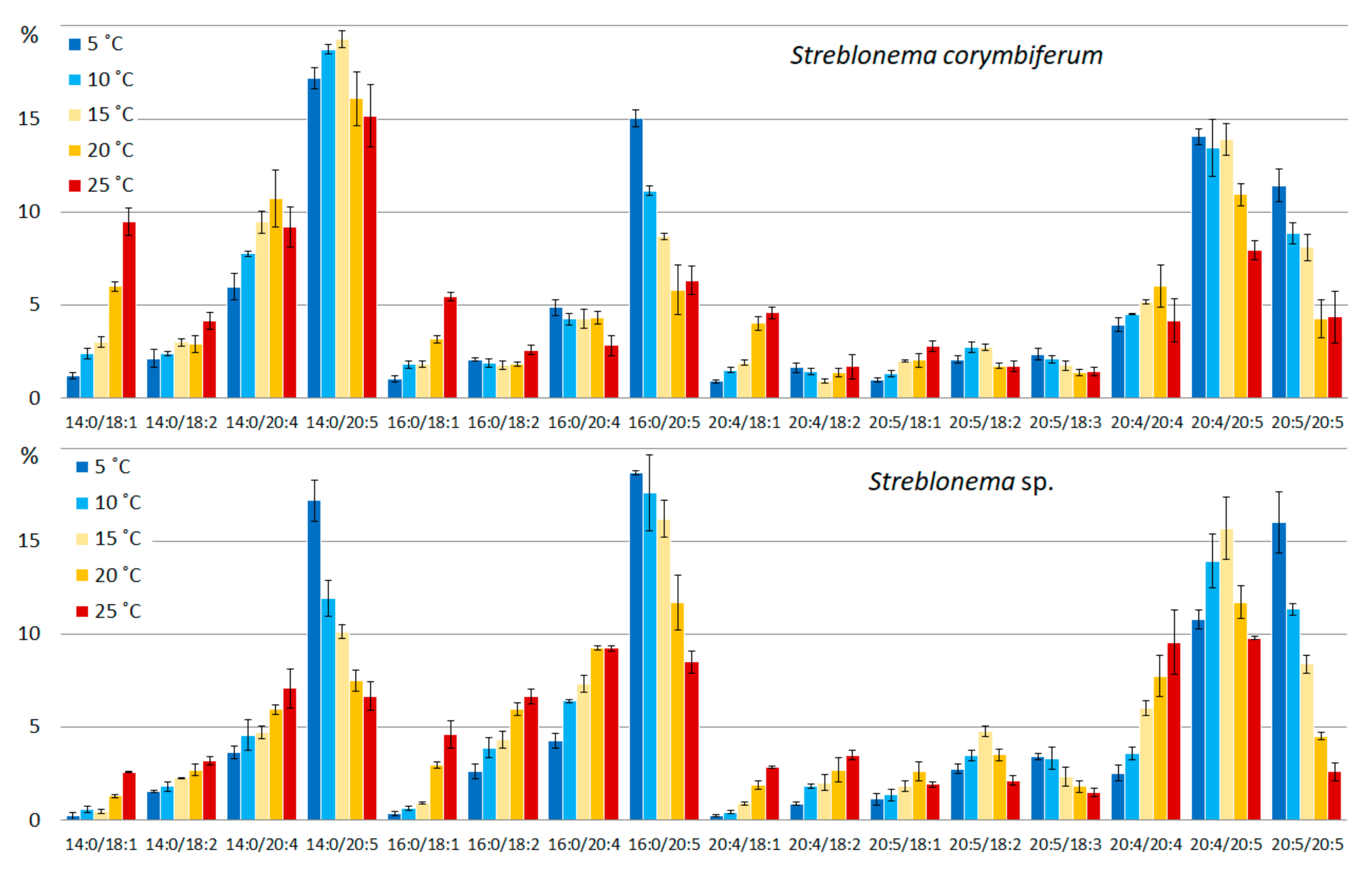
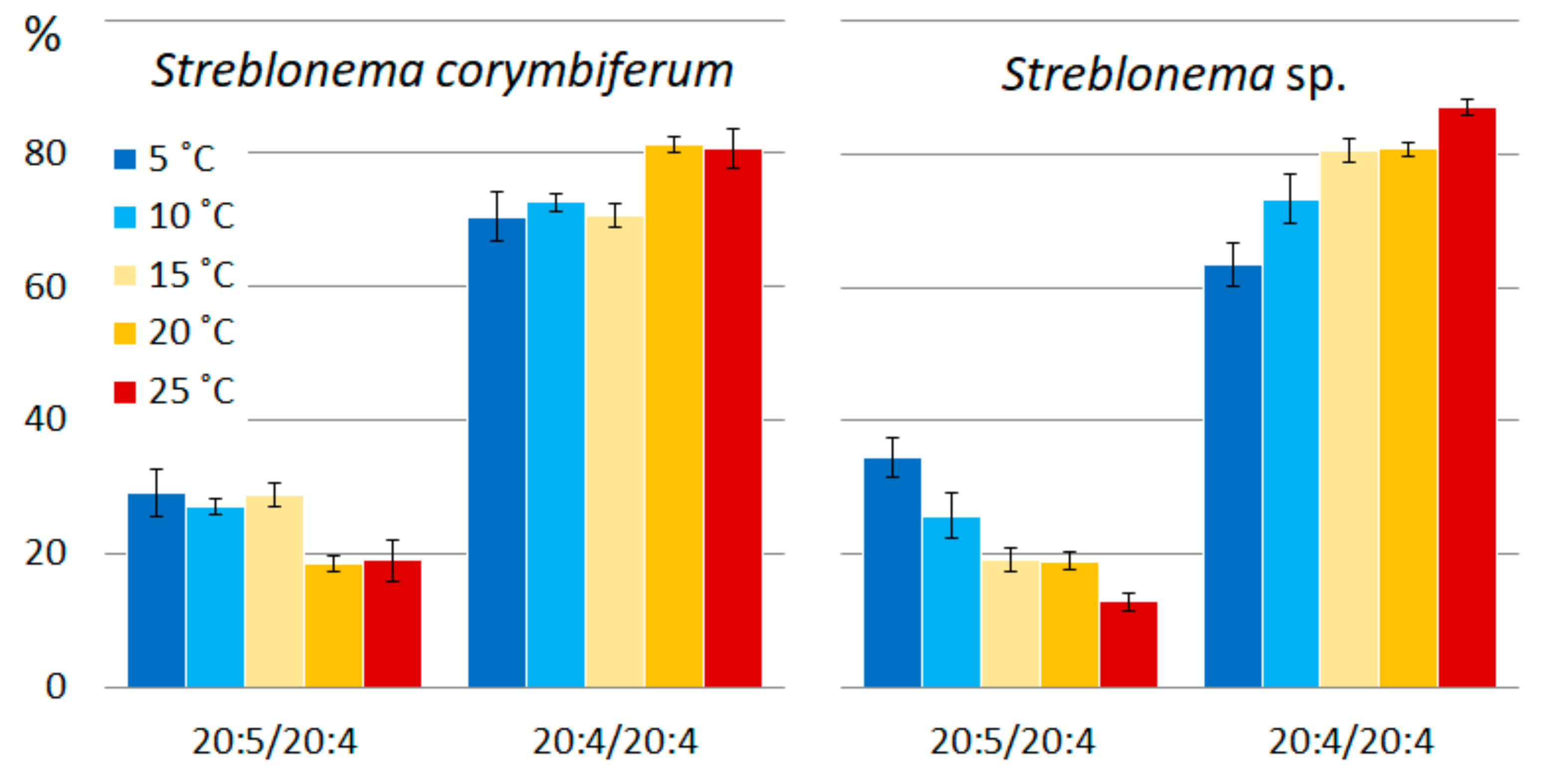
2.2. Effect of Temperature on the Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
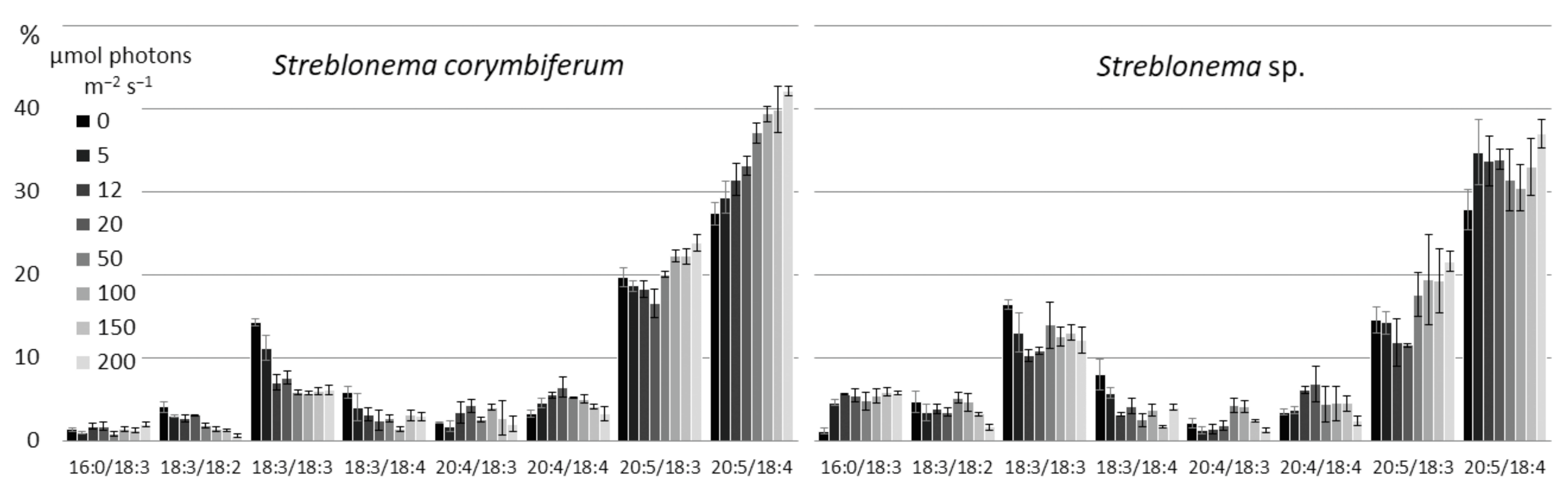
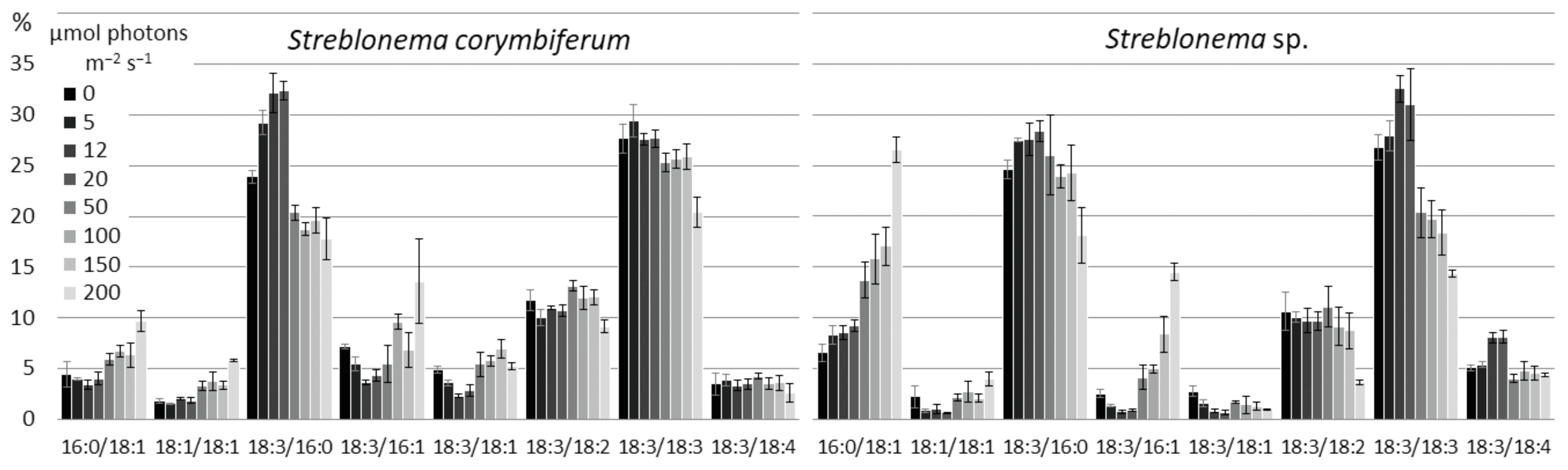
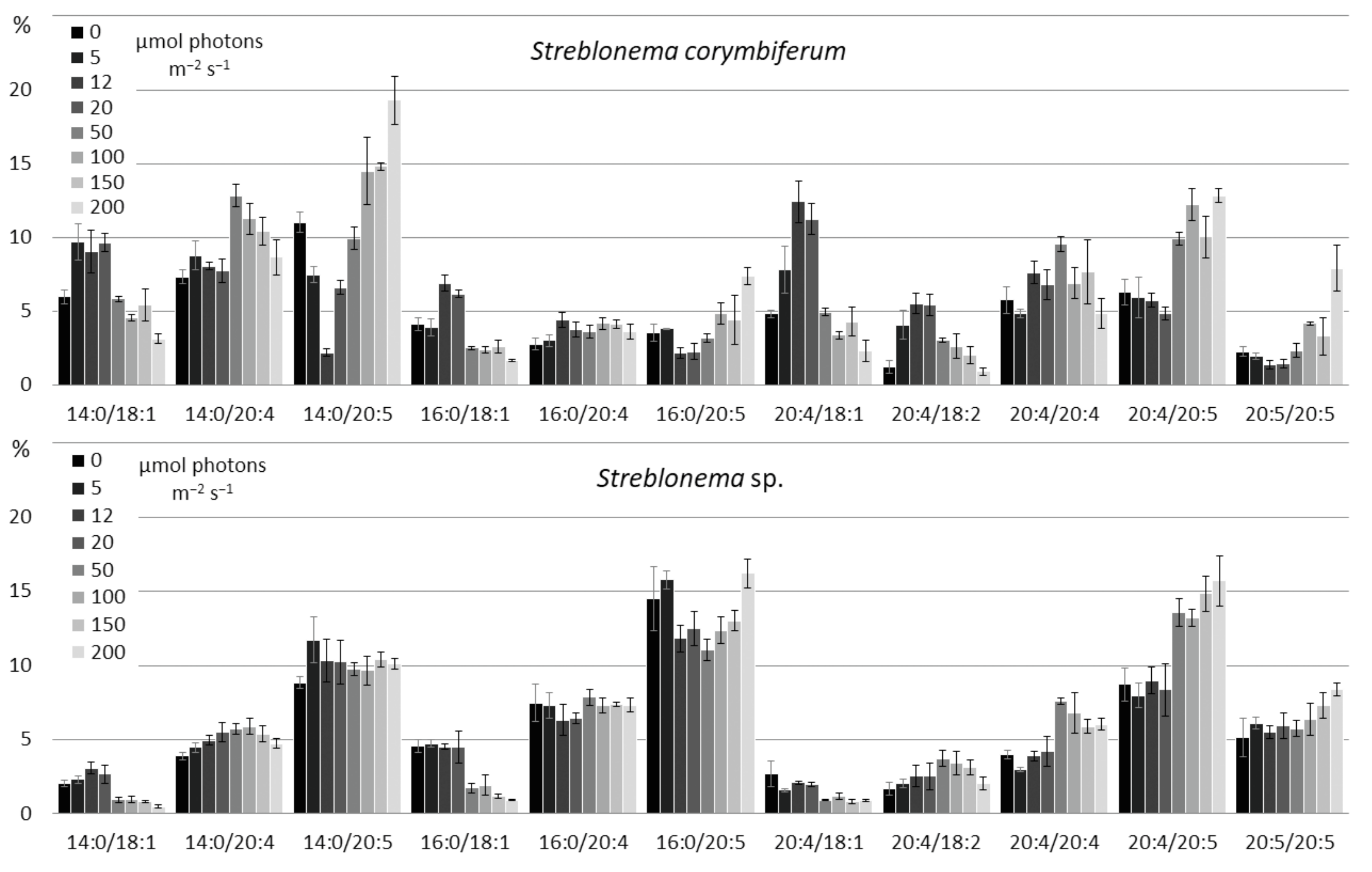
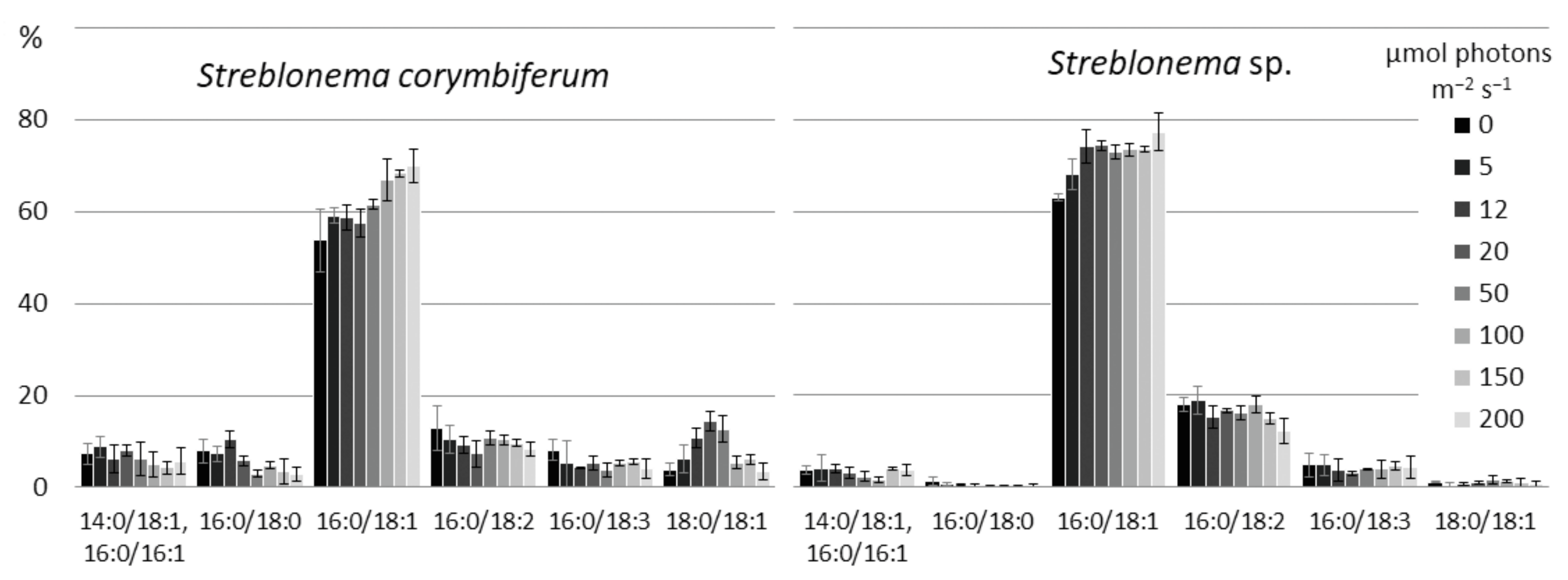
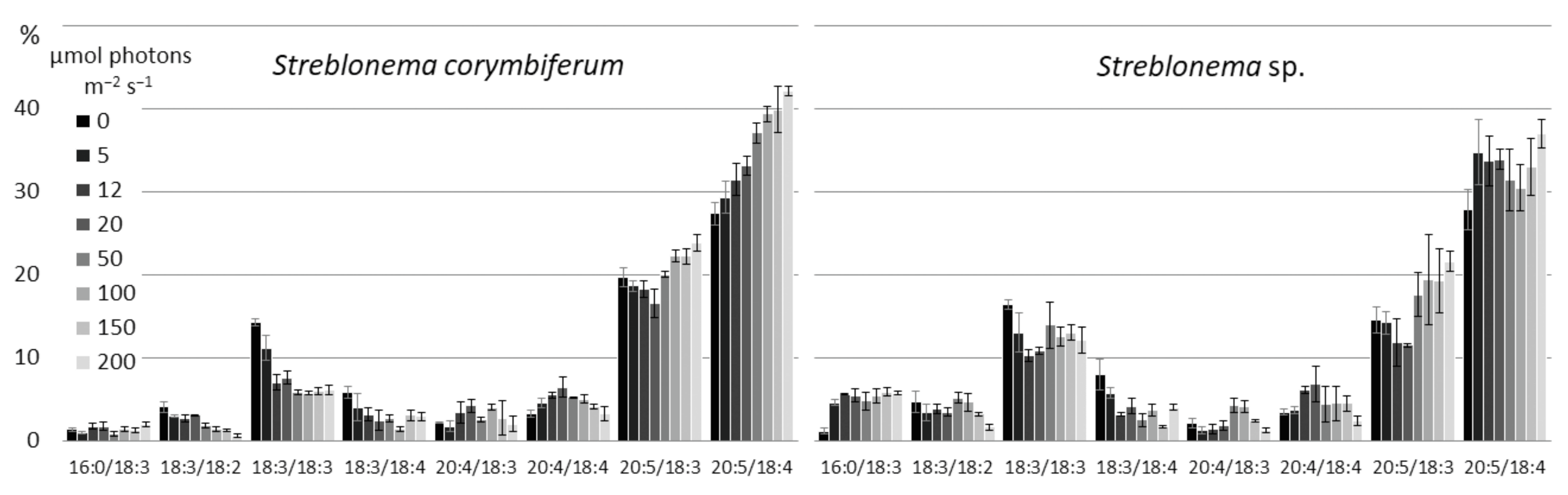
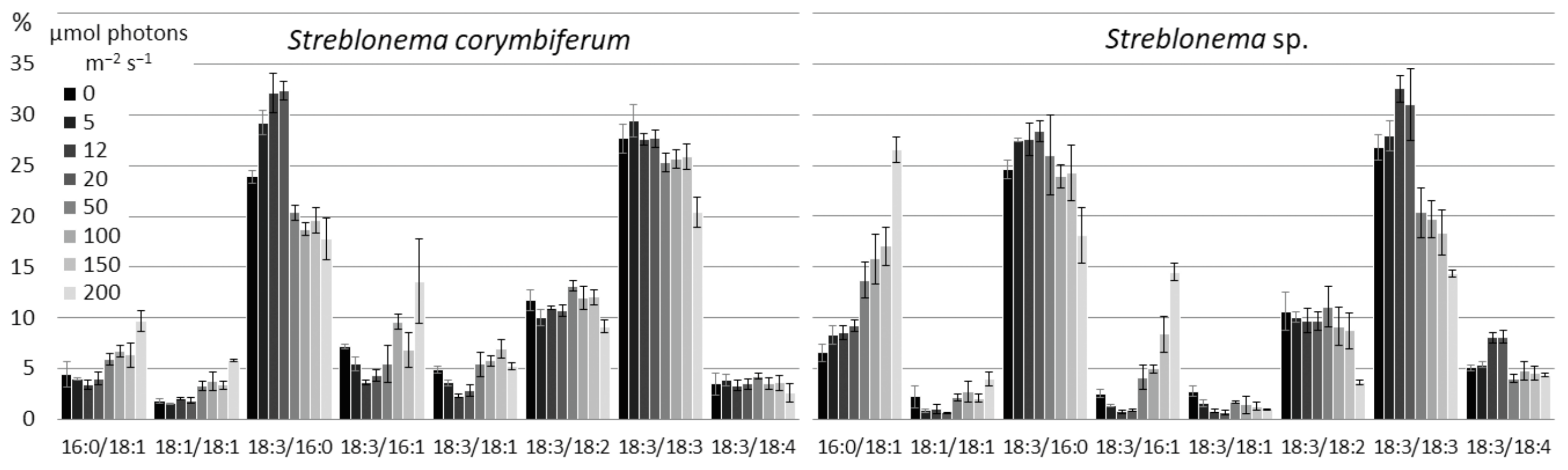
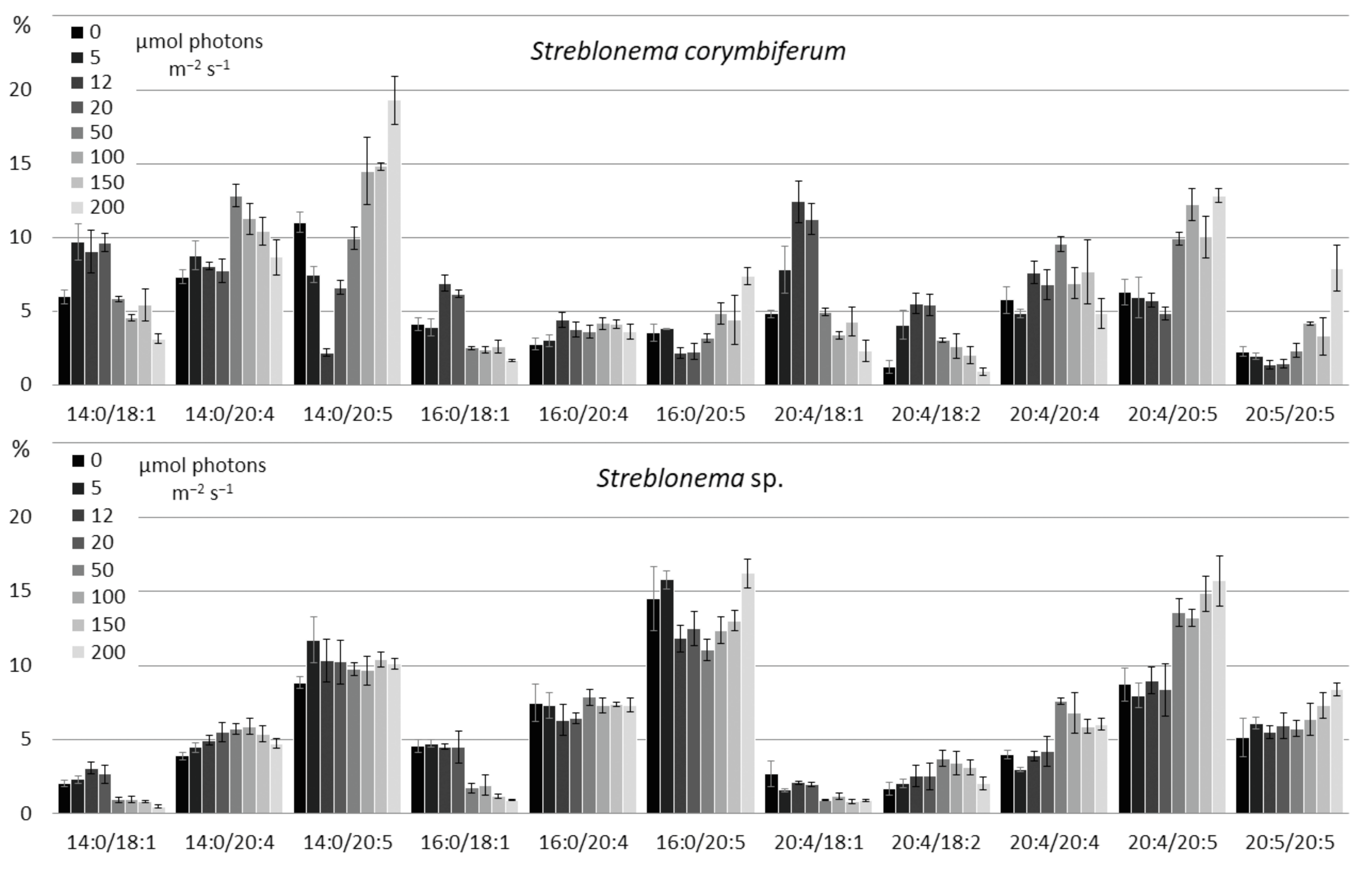
2.3. Effect of Light Intensity on the Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
3. Discussion
3.1. The Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
3.2. Effect of Temperature on the Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
3.3. Effect of Light Intensity on the Polar Lipidome of Streblonema corymbiferum and Streblonema sp.
4. Materials and Methods
4.1. Algae material
4.2. Temperature Treatment
4.3. Light Treatments
4.4. Lipid Extraction
4.5. HPLC-MS/MS Analysis of Molecular Species
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BL | Betaine lipid |
| DAG | Diacylglycerol |
| DGDG | Digalactosyldiacylglycerol |
| DGTS | Diacylglyceryl-N,N,N-trimethylhomoserine |
| ER | Endoplasmic reticulum |
| FA | Fatty acid |
| GL | Glycoglycerolipid |
| GlcADG | Glucuronosyldiacylglycerol |
| LHCII | Light-harvesting complex II |
| MGDG | Monogalactosyldiacylglycerol |
| MUFA | Monounsaturated fatty acid |
| PC | Phosphatidylcholine |
| PE | Phosphatidylethanolamine |
| PG | Phosphatidylglycerol |
| PHEG | Phosphatidylhydroxyethylglycine |
| PI | Phosphatidylinositol |
| PL | Phosphoglycerolipid |
| PSII | Photosystem II |
| PUFA | Polyunsaturated fatty acid |
| SFA | Saturated fatty acid |
| SQD2 | SQDG synthase |
| SQDG | Sulfoquinovosyldiacylglycerol |
| UDP-GlcA | UDP-glucuronic acid |
References
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. Algal Lipids, Fatty Acids and Sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 87–134. ISBN 978–085–709–512–1. [Google Scholar]
- Serviere-Zaragoza, E.; Hurtado, M.A.; Manzano-Sarabia, M.; Mazariegos-Villarreal, A.; Reza, M.; Arjona, O.; Palacios, E. Seasonal and Interannual Variation of Fatty Acids in Macrophytes from the Pacific Coast of Baja California Peninsula (Mexico). J. Appl. Phycol. 2015, 27, 1297–1306. [Google Scholar] [CrossRef]
- Gosch, B.J.; Paul, N.A.; de Nys, R.; Magnusson, M. Seasonal and Within-Plant Variation in Fatty Acid Content and Composition in the Brown Seaweed Spatoglossum macrodontum (Dictyotales, Phaeophyceae). J. Appl. Phycol. 2015, 27, 387–398. [Google Scholar] [CrossRef]
- Kalacheva, G.S.; Zhila, N.O.; Volova, T.G.; Gladyshev, M.I. The Effect of Temperature on the Lipid Composition of the Green Alga Botryococcus. Microbiology 2002, 71, 286–293. [Google Scholar] [CrossRef]
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Seasonal Changes of Fatty Acid Composition and Thermotropic Behavior of Polar Lipids from Marine Macrophytes. Phytochemistry 2008, 69, 1517–1527. [Google Scholar] [CrossRef]
- Tatsuzawa, H.; Takizawa, E. Changes in Lipid and Fatty Acid Composition of Pavlova lutheri. Phytochemistry 1995, 40, 397–400. [Google Scholar] [CrossRef]
- Song, Y.; Zhao, J.; Chen, J.; Luo, Q.; Yang, R.; Xu, J.; Chen, H.; Yan, X. Heat Shock-Induced Metabolic Conversion of Membrane Lipids, Fatty Acids and Volatile Organic Compounds of Pyropia haitanensis under Different Heat Shock Time. Phycol. Res. 2018, 66, 89–99. [Google Scholar] [CrossRef]
- Kostetsky, E.; Chopenko, N.; Barkina, M.; Velansky, P.; Sanina, N. Fatty Acid Composition and Thermotropic Behavior of Glycolipids and Other Membrane Lipids of Ulva lactuca (Chlorophyta) Inhabiting Different Climatic Zones. Mar. Drugs 2018, 16, 494. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Xu, J.; Zong, C.; Zhu, S.; Ye, M.; Zhou, C.; Chen, H.; Yan, X. Effect of High Temperature on the Lipid Composition of Isochrysis galbana Parke in Logarithmic Phase. Aquac. Int. 2017, 25, 327–339. [Google Scholar] [CrossRef]
- Gerasimenko, N.I.; Busarova, N.G.; Logvinov, S.V. Seasonal Changes in the Content of Lipids and Photosynthetic Pigments in a Brown Alga Saccharina cichorioides. Russ. J. Plant Physiol. 2014, 61, 893–898. [Google Scholar] [CrossRef]
- Da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomic Signatures Reveal Seasonal Shifts on the Relative Abundance of High-Valued Lipids from the Brown Algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.S.P.; da Costa, E.; Melo, T.; Sulpice, R.; Cardoso, S.M.; Pitarma, B.; Pereira, R.; Abreu, M.H.; Domingues, P.; Calado, R.; et al. Seasonal Plasticity of the Polar Lipidome of Ulva rigida Cultivated in a Sustainable Integrated Multi-Trophic Aquaculture. Algal Res. 2020, 49, 101958. [Google Scholar] [CrossRef]
- Barkina, M.Y.; Pomazenkova, L.A.; Chopenko, N.S.; Velansky, P.V.; Kostetsky, E.Y.; Sanina, N.M. Effect of Warm Acclimation Rate on Fatty Acid Composition and Phase Transitions of Saccharina japonica (J.E. Areschoug) Glycolipids. Vestn. Tomsk. Gos. Univ. Biol. 2019, 48, 135–157. [Google Scholar] [CrossRef]
- Barkina, M.Y.; Pomazenkova, L.A.; Chopenko, N.S.; Velansky, P.V.; Kostetsky, E.Y.; Sanina, N.M. Influence of Warm-Acclimation Rate on Polar Lipids of Ulva lactuca. Russ. J. Plant Physiol. 2020, 67, 111–121. [Google Scholar] [CrossRef]
- Thompson, P.A.; Harrison, P.J.; Whyte, J.N.C. Influence of Irradiance on the Fatty Acid Composition of Phytoplankton. J. Phycol. 1990, 26, 278–288. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Yakovleva, I.M. Lipid Composition of the Red Alga Tichocarpus crinitus Exposed to Different Levels of Photon Irradiance. Phytochemistry 2005, 66, 73–79. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Yakovleva, I.M. Effect of Solar Irradiance on Lipids of the Green Alga Ulva fenestrata Postels et Ruprecht. Bot. Mar. 2004, 47, 395–401. [Google Scholar] [CrossRef]
- Giossi, C.E.; Cruz, S.; Rey, F.; Marques, R.; Melo, T.; Domingues, M.d.R.; Cartaxana, P. Light Induced Changes in Pigment and Lipid Profiles of Bryopsidales Algae. Front. Mar. Sci. 2021, 8, 745083. [Google Scholar] [CrossRef]
- Wynne, M.J. Marine Algae and Early Explorations in the Upper North Pacific and Bering Sea. Algae 2009, 24, 1–29. [Google Scholar] [CrossRef]
- Skriptsova, A.V. Nitrogen Effect on Water-Soluble Polysaccharide Accumulation in Streblonema sp. (Ectocarpales, Phaeophyceae). Mar. Biotechnol. 2017, 19, 410–419. [Google Scholar] [CrossRef]
- Khotimchenko, S.V. Lipids of Marine Macrophytic Algae and Grasses: Structure, Distribution, Analysis; Svetashev, V.I., Ed.; Dal’nauka: Vladivostok, Russia, 2003; ISBN 5-8044-0347-8. [Google Scholar]
- Okazaki, Y.; Otsuki, H.; Narisawa, T.; Kobayashi, M.; Sawai, S.; Kamide, Y.; Kusano, M.; Aoki, T.; Hirai, M.Y.; Saito, K. A New Class of Plant Lipid Is Essential for Protection against Phosphorus Depletion. Nat. Commun. 2013, 4, 1510. [Google Scholar] [CrossRef] [Green Version]
- Eichenberger, W.; Gribi, C. Diacylglyceryl-α-D-Glucuronide from Ochromonas danica (Chrysophyceae). J. Plant Physiol. 1994, 144, 272–276. [Google Scholar] [CrossRef]
- Koelmel, J.P.; Campbell, J.E.; Guingab-Cagmat, J.; Meke, L.; Garrett, T.J.; Stingl, U. Re-Modeling of Foliar Membrane Lipids in a Seagrass Allows for Growth in Phosphorus-Deplete Conditions. PLoS ONE 2019, 14, e0218690. [Google Scholar] [CrossRef] [Green Version]
- Hölzl, G.; Dörmann, P. Structure and Function of Glycoglycerolipids in Plants and Bacteria. Prog. Lipid Res. 2007, 46, 225–243. [Google Scholar] [CrossRef]
- Chadova, O.A.; Velansky, P.V. Influence of Endophyte Lamimariocolax aecidioides (Rosenvinge) A.F. Peters, 1998 (Phaeophyceae: Ectocarpales) on the Lipid Composition of the Brown Alga Undaria pinnatifida (Harvey) Suringar, 1873 (Phaeophyceae: Laminariales). Russ. J. Mar. Biol. 2022; accepted. [Google Scholar]
- Muller, D.G.; Eichenberger, W. Betaine Lipid Content and Species Delimitation in Ectocarpus, Feldmannia and Hincksia (Ectocarpales, Phaeophyceae). Eur. J. Phycol. 1994, 29, 219–225. [Google Scholar] [CrossRef]
- Vogel, G.; Eichenberger, W. Betaine Lipids in Lower Plants. Biosynthesis of DGTS and DGTA in Ochromonas danica (Chrysophyceae) and the Possible Role of DGTS in Lipid Metabolism. Plant Cell Physiol. 1992, 33, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Los, D.A.; Mironov, K.S.; Allakhverdiev, S.I. Regulatory Role of Membrane Fluidity in Gene Expression and Physiological Functions. Photosynth. Res. 2013, 116, 489–509. [Google Scholar] [CrossRef]
- Dörmann, P.; Hölzl, G. The Role of Glycolipids in Photosynthesis. In Lipids in Photosynthesis: Essential and Regulatory Functions; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; pp. 265–282. ISBN 978–904–812–862–4. [Google Scholar]
- Mizusawa, N.; Wada, H. The Role of Lipids in Photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Gombos, Z.; Wada, H.; Murata, N. The Recovery of Photosynthesis from Low-Temperature Photoinhibition Is Accelerated by the Unsaturation of Membrane Lipids: A Mechanism of Chilling Tolerance. Proc. Natl. Acad. Sci. USA 1994, 91, 8787–8791. [Google Scholar] [CrossRef] [Green Version]
- Moon, B.Y.; Higashi, S.I.; Gombos, Z.; Murata, N. Unsaturation of the Membrane Lipids of Chloroplasts Stabilizes the Photosynthetic Machinery against Low-Temperature Photoinhibition in Transgenic Tobacco Plants. Proc. Natl. Acad. Sci. USA 1995, 92, 6219–6223. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.-Q.; Zhao, H.; Liang, J.-S.; Ji, B.-H.; Jiao, D.-M. Relationships between Phosphatidylglycerol Molecular Species of Thylakoid Membrane Lipids and Sensitivities to Chilling-Induced Photoinhibition in Rice. J. Integr. Plant Biol. 2008, 50, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Botella, C.; Jouhet, J.; Block, M.A. Importance of Phosphatidylcholine on the Chloroplast Surface. Prog. Lipid Res. 2017, 65, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zheng, Q.; Shen, W.; Cram, D.; Brian Fowler, D.; Wei, Y.; Zou, J. Understanding the Biochemical Basis of Temperature-Induced Lipid Pathway Adjustments in Plants. Plant Cell 2015, 27, 86–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Gügel, I.L.; Giavalisco, P.; Zeisler, V.; Schreiber, L.; Soll, J.; Philippar, K. FAX1, a Novel Membrane Protein Mediating Plastid Fatty Acid Export. PLoS Biol. 2015, 13, e1002053. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Benning, C. Chloroplast Lipid Synthesis and Lipid Trafficking through ER-Plastid Membrane Contact Sites. Biochem. Soc. Trans. 2012, 40, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Khozin, I.; Adlerstein, D.; Bigongo, C.; Heimer, Y.M.; Cohen, Z. Elucidation of the Biosynthesis of Eicosapentaenoic Acid in the Microalga Porphyridium cruentum. Plant Physiol. 1997, 114, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.S.; Pinto, R.F.; Sant’Anna, C. Low Light Intensity and Nitrogen Starvation Modulate the Chlorophyll Content of Scenedesmus dimorphus. J. Appl. Microbiol. 2016, 120, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N. ROS-Dependent Signalling Pathways in Plants and Algae Exposed to High Light: Comparisons with Other Eukaryotes. Free Radic. Biol. Med. 2018, 122, 52–64. [Google Scholar] [CrossRef]
- Mock, T.; Kroon, B.M.A. Photosynthetic Energy Conversion under Extreme Conditions—II: The Significance of Lipids under Light Limited Growth in Antarctic Sea Ice Diatoms. Phytochemistry 2002, 61, 53–60. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Yakovleva, I.M. Low Light Acclimation Strategy of the Brown Macroalga Undaria pinnatifida: Significance of Lipid and Fatty Acid Remodeling for Photosynthetic Competence. J. Phycol. 2021, 57, 1792–1804. [Google Scholar] [CrossRef]
- Klyachko-Gurvich, G.L.; Tsoglin, L.N.; Doucha, J.; Kopetskii, J.; Shebalina Ryabykh, I.B.; Semenenko, V.E. Desaturation of Fatty Acids as an Adaptive Response to Shifts in Light Intensity. Physiol. Plant. 1999, 107, 240–249. [Google Scholar] [CrossRef]
- Goss, R.; Lohr, M.; Latowski, D.; Grzyb, J.; Vieler, A.; Wilhelm, C.; Strzalka, K. Role of Hexagonal Structure-Forming Lipids in Diadinoxanthin and Violaxanthin Solubilization and de-Epoxidation. Biochemistry 2005, 44, 4028–4036. [Google Scholar] [CrossRef]
- Mizusawa, N.; Sakurai, I.; Sato, N.; Wada, H. Lack of Digalactosyldiacylglycerol Increases the Sensitivity of Synechocystis sp. PCC 6803 to High Light Stress. FEBS Lett. 2009, 583, 718–722. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Siegert, E.; Stepushenko, O.; Glauser, G.; Farmer, E.E. Membranes as Structural Antioxidants: Recycling of Malondialdehyde to Its Source in Oxidation- Sensitive Chloroplast Fatty Acids. J. Biol. Chem. 2016, 291, 13005–13013. [Google Scholar] [CrossRef] [Green Version]
- Gray, G.R.; Ivanov, A.G.; Król, M.; Williams, J.P.; Kahn, M.U.; Myscich, E.G.; Huner, N.P.A. Temperature and Light Modulate the Trans-Δ3-Hexadecenoic Acid Content of Phosphatidylglycerol: Light-Harvesting Complex II Organization and Non-Photochemical Quenching. Plant Cell Physiol. 2005, 46, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Provasoli, L. Media and Prospects for the Cultivation of Marine Algae. In Cultures and Collections of Agae, Proceedings of the U.S.–Japan Conference, Hakone, Japan, September 1966; Watanabe, A., Hattori, A., Eds.; Japanese Society Plant Physiology: Kyoto, Japan, 1968; pp. 63–75. [Google Scholar]
- Abbott, I.A.; Hollenberg, G.J. Marine Algae of California; Stanford University Press: Stanford, CA, USA, 1976; p. 827. [Google Scholar]
- Luan, R.; Ding, L.; Lu, B.; Tseng, C.K. Flora Algarum Marinarum Sinicarum. Tomus III. Phaeophyta No. I(1) Ectocarpales Ralfsiales Sphaceariales Dictyotales; Science Press: Beijing, China, 2013. (In Chinese) [Google Scholar]
- Norris, J.N. Marine algae of the Northern Gulf of California: Chlorophyta and Phaeophyceae. Smithson. Contrib. Bot. 2010, 94. [Google Scholar] [CrossRef] [Green Version]
- Perestenko, L.P. Vodorosli Zaliva Petra Velikogo [The Seaweeds of Peter the Great Bay]; NAUKA Leningradskoe Otdelenie: Leningrad, Russia, 1980; p. 231. (In Russian) [Google Scholar]
- Tseng, C.K. Seaweeds in Yellow Sea and Bohai Sea of China; Science Press: Beijing, China, 2009; ISBN 9787030249968. (In Chinese) [Google Scholar]
- Womersley, H.B.S. The Marine Benthic Flora of Southern Australia, Part II; Government Printer: Adelaide, South Australia, 1987. [Google Scholar]
- Tatituri, R.V.V.; Brenner, M.B.; Turk, J.; Hsu, F.-F. Structural Elucidation of Diglycosyl Diacylglycerol and Monoglycosyl Diacylglycerol from Streptococcus pneumoniae by Multiple-Stage Linear Ion-Trap Mass Spectrometry with Electrospray Ionization. J. Mass Spectrom. 2012, 47, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Granafei, S.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Unambiguous Regiochemical Assignment of Sulfoquinovosyl Mono- and Diacylglycerols in Parsley and Spinach Leaves by Liquid Chromatography/Electrospray Ionization Sequential Mass Spectrometry Assisted by Regioselective Enzymatic Hydrolysis. Rapid Commun. Mass Spectrom. 2017, 31, 1499–1509. [Google Scholar] [CrossRef]
- Hou, W.; Zhou, H.; Khalil, M.B.; Seebun, D.; Bennett, S.A.L.; Figeys, D. Lyso-Form Fragment Ions Facilitate the Determination of Stereospecificity of Diacyl Glycerophospholipids. Rapid Commun. Mass Spectrom. 2011, 25, 205–217. [Google Scholar] [CrossRef]
- Hsu, F.-F.; Turk, J. Electrospray Ionization with Low-Energy Collisionally Activated Dissociation Tandem Mass Spectrometry of Glycerophospholipids: Mechanisms of Fragmentation and Structural Characterization. J. Chromatogr. B 2009, 877, 2673–2695. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lou, Y.; Mu, T.; Xu, J.; Zhou, C.; Yan, X. Simultaneous Structural Identification of Diacylglyceryl-N-Trimethylhomoserine (DGTS) and Diacylglycerylhydroxymethyl-N,N,N-Trimethyl-β-Alanine (DGTA) in Microalgae Using Dual Li+/H+ Adduct Ion Mode by Ultra-Performance Liquid Chromatography/Quadrupole Ti. Rapid Commun. Mass Spectrom. 2017, 31, 457–468. [Google Scholar] [CrossRef]
- Iba, K. Acclimative Response to Temperature Stress in Higher Plants: Approaches of Gene Engineering for Temperature Tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Total Number of Molecular Species | Major Molecular Species |
|---|---|---|
| Glycoglycerolipids | ||
| MGDG | 113 | 18:2/18:4, 18:3/18:2, 18:3/18:3, 18:3/18:4, 18:4/18:4, 20:5/18:2, 20:5/18:3, 20:5/18:4 |
| DGDG | 63 | 16:0/18:3, 18:3/18:2, 18:3/18:3,18:3/18:4, 20:4/18:3, 20:4/18:4, 20:5/18:1, 20:5/18:2, 20:5/18:3, 20:5/18:4 |
| SQDG | 49 | 14:0/16:0, 14:0/18:1, 14:0/18:2, 14:0/18:3, 16:0/16:0,16:0/16:1, 16:1/16:0, 18:1/16:0, 18:2/16:0, 18:3/16:0 |
| GlcADG | 9 | 16:0/18:1, 16:0/18:2, 16:0/18:3, 18:1/18:1, 18:1/18:2, 18:1/18:3, 20:5/18:2, 20:5/18:3, 20:5/18:4 |
| Phosphoglycerolipids | ||
| PG | 42 | 16:0/18:2, 16:0/18:1, 18:2/16:0, 18:3/16:0, 18:3/16:1, 18:1/18:1, 18:2/18:1, 18:3/18:1, 18:3/18:2, 18:3/18:3, 18:3/18:4 |
| PE | 42 | 14:0/20:4, 16:0/20:5, 16:0/20:4, 18:0/20:4, 20:0/20:4, 18:1/16:1, 18:3/20:5, 20:4/20:4, 20:4/20:5, 20:5/20:5 |
| PC | 85 | 14:0/18:1, 14:0/18:2, 14:0/20:4, 14:0/20:5, 16:0/18:1, 16:0/18:2, 16:0/20:4, 16:0/20:5, 20:4/18:1, 20:4/18:2, 20:4/20:4, 20:4/20:5, 20:5/20:5 |
| PI | 9 | 14:0/18:1, 16:0/16:1, 16:0/18:0, 16:0/18:1, 16:0/18:2, 16:0/18:3, 18:0/18:1 |
| PHEG | 3 | 20:4/20:4, 20:5/20:4 |
| Betaine lipid | ||
| DGTS | 45 | 16:0/18:1, 18:1/18:2, 18:1/18:1, 16:0/20:1, 18:1/18:0, 18:1/19:1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadova, O.; Skriptsova, A.; Velansky, P. Effect of Temperature and Light Intensity on the Polar Lipidome of Endophytic Brown Algae Streblonema corymbiferum and Streblonema sp. In Vitro. Mar. Drugs 2022, 20, 428. https://doi.org/10.3390/md20070428
Chadova O, Skriptsova A, Velansky P. Effect of Temperature and Light Intensity on the Polar Lipidome of Endophytic Brown Algae Streblonema corymbiferum and Streblonema sp. In Vitro. Marine Drugs. 2022; 20(7):428. https://doi.org/10.3390/md20070428
Chicago/Turabian StyleChadova, Oksana, Anna Skriptsova, and Peter Velansky. 2022. "Effect of Temperature and Light Intensity on the Polar Lipidome of Endophytic Brown Algae Streblonema corymbiferum and Streblonema sp. In Vitro" Marine Drugs 20, no. 7: 428. https://doi.org/10.3390/md20070428
APA StyleChadova, O., Skriptsova, A., & Velansky, P. (2022). Effect of Temperature and Light Intensity on the Polar Lipidome of Endophytic Brown Algae Streblonema corymbiferum and Streblonema sp. In Vitro. Marine Drugs, 20(7), 428. https://doi.org/10.3390/md20070428