
Anti-Obesity and Gut Microbiota Regulation Effects of Phospholipids from the Eggs of Crab, Portunus Trituberculatus, in High Fat Diet-Fed Mice

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. The Composition of Phospholipids of Pt-PL and Their Fatty Acids
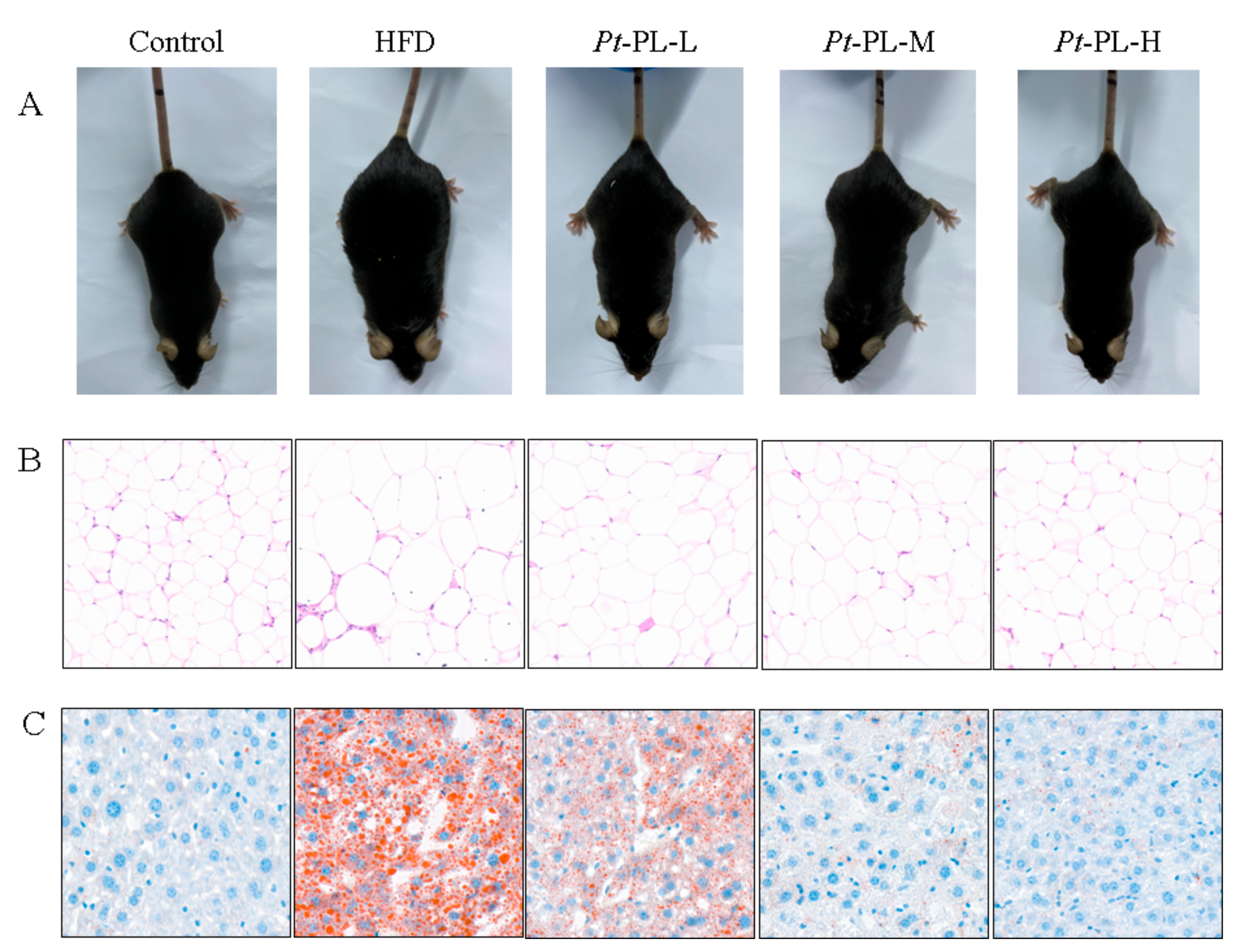
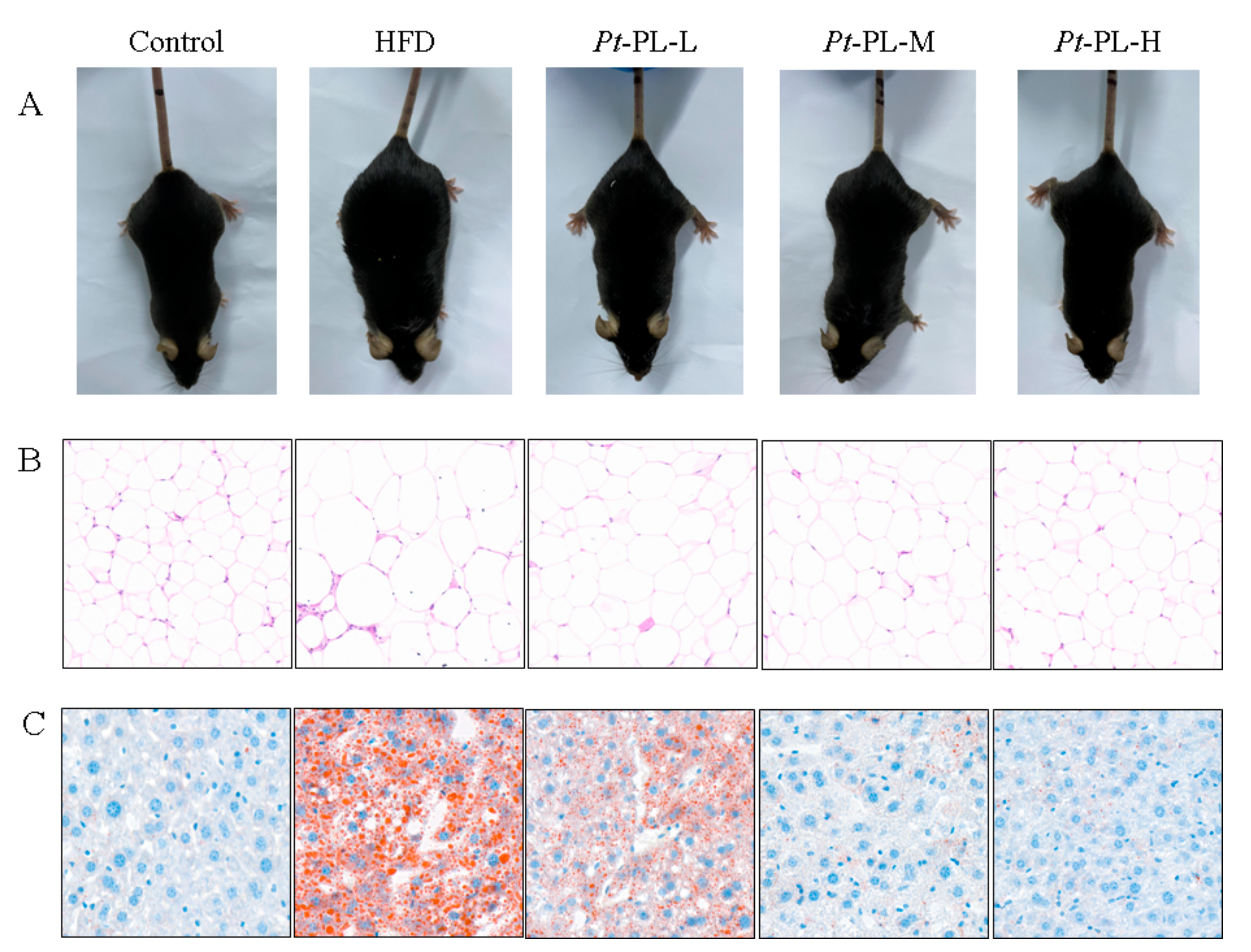
2.2. Effects of Pt-PL on Obesity
2.3. Effects of Pt-PL on Serum Lipids and Hepatic Lipids
2.4. Effects of Pt-PL on Blood Glucose, Insulin, and Inflammatory Cytokines
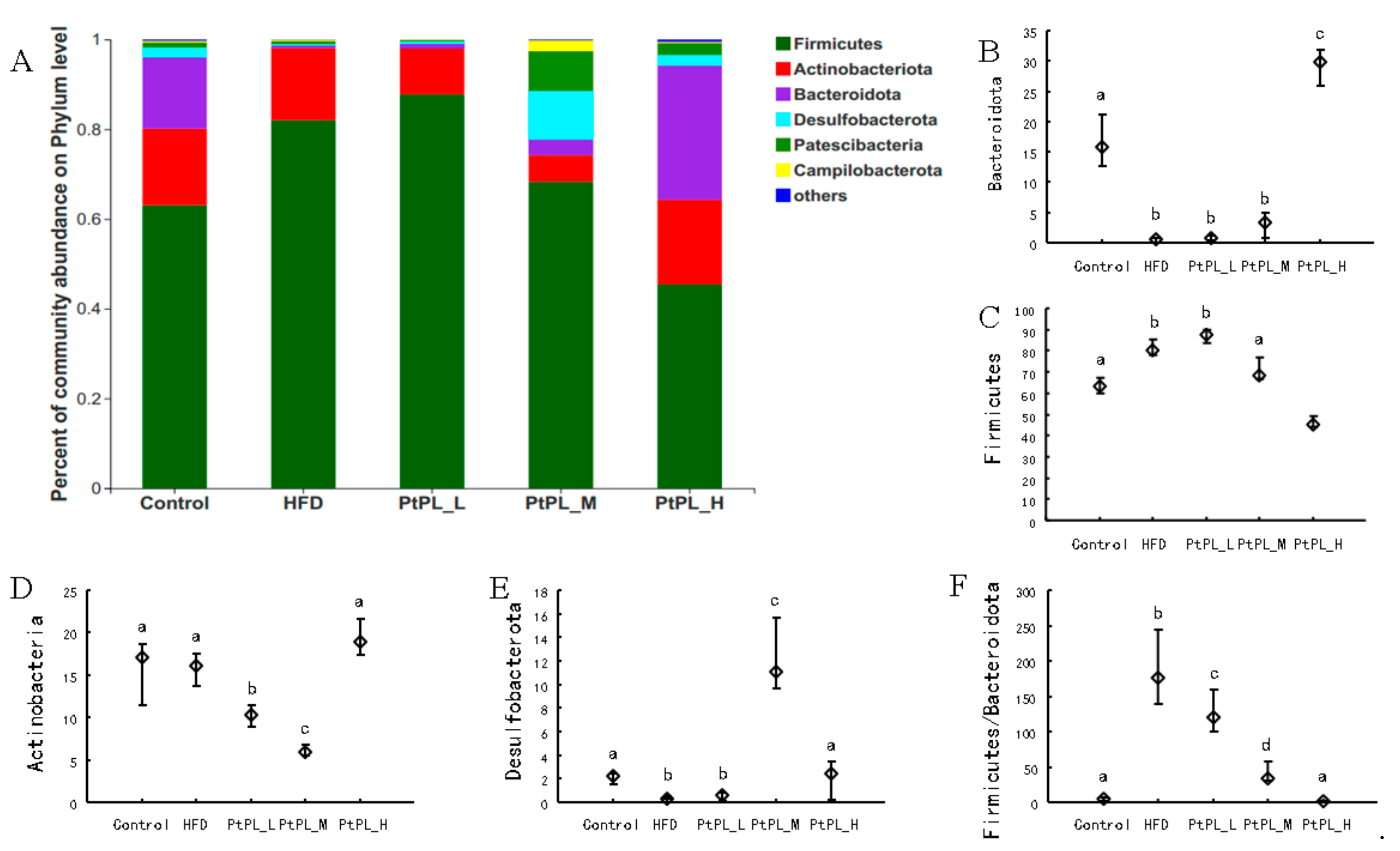
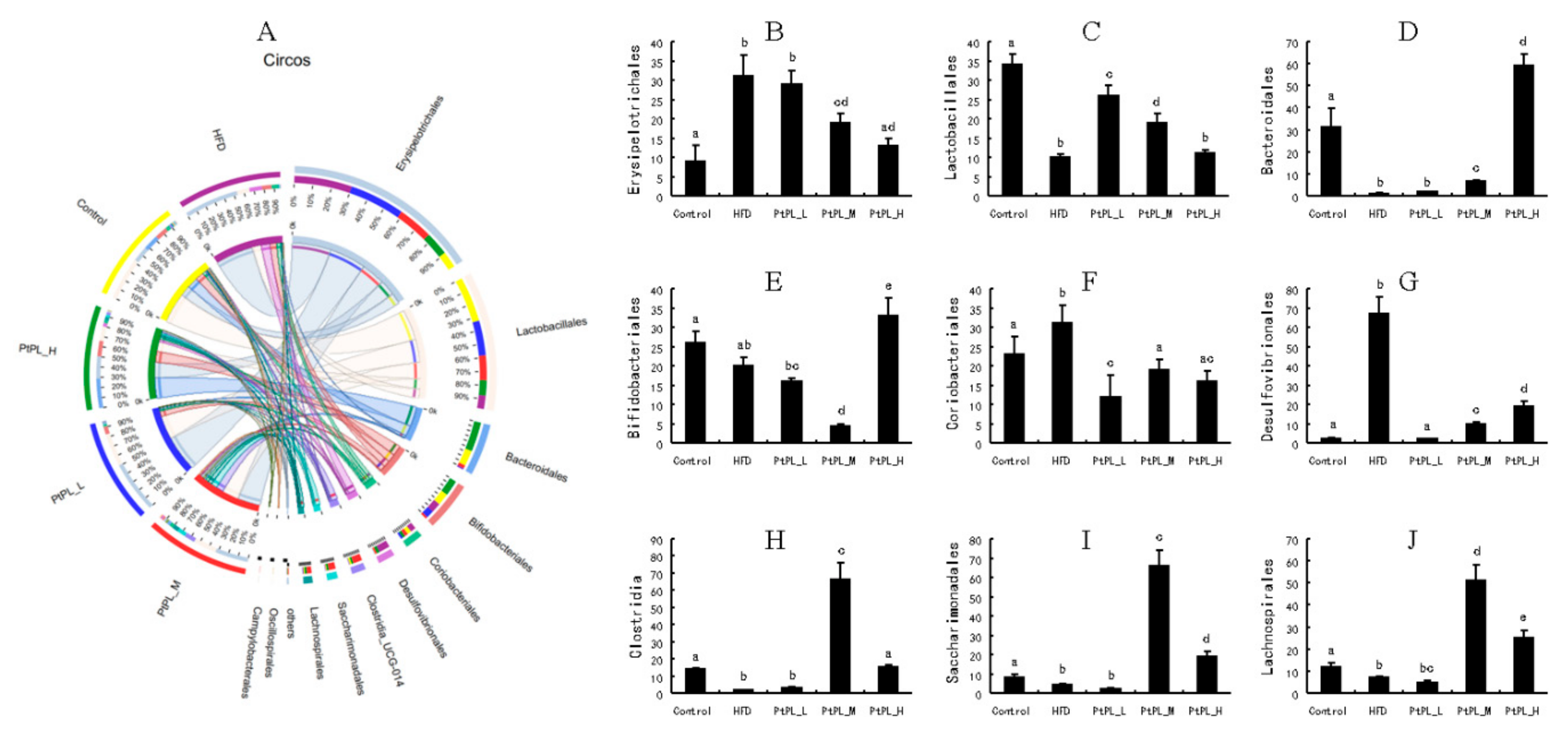
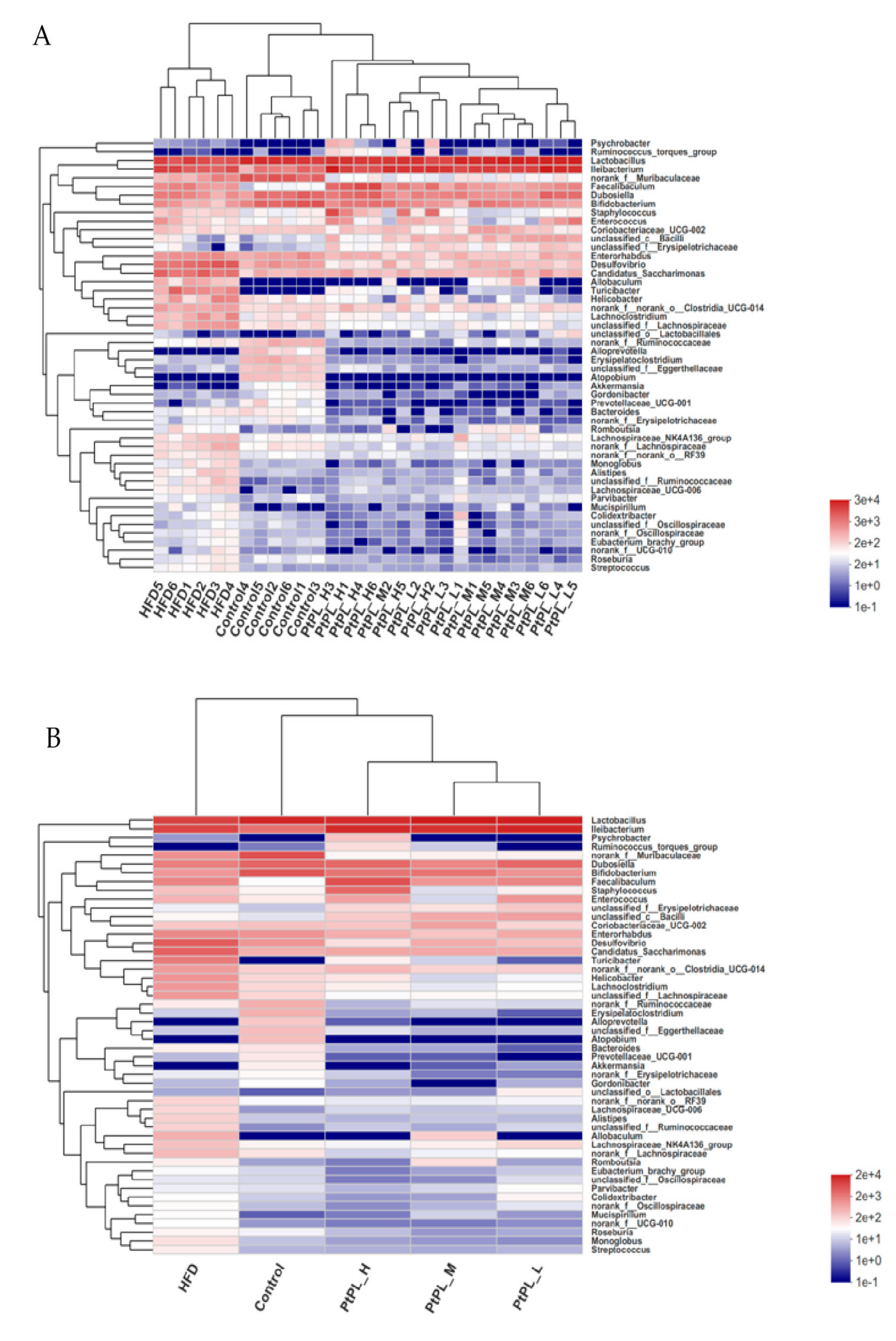
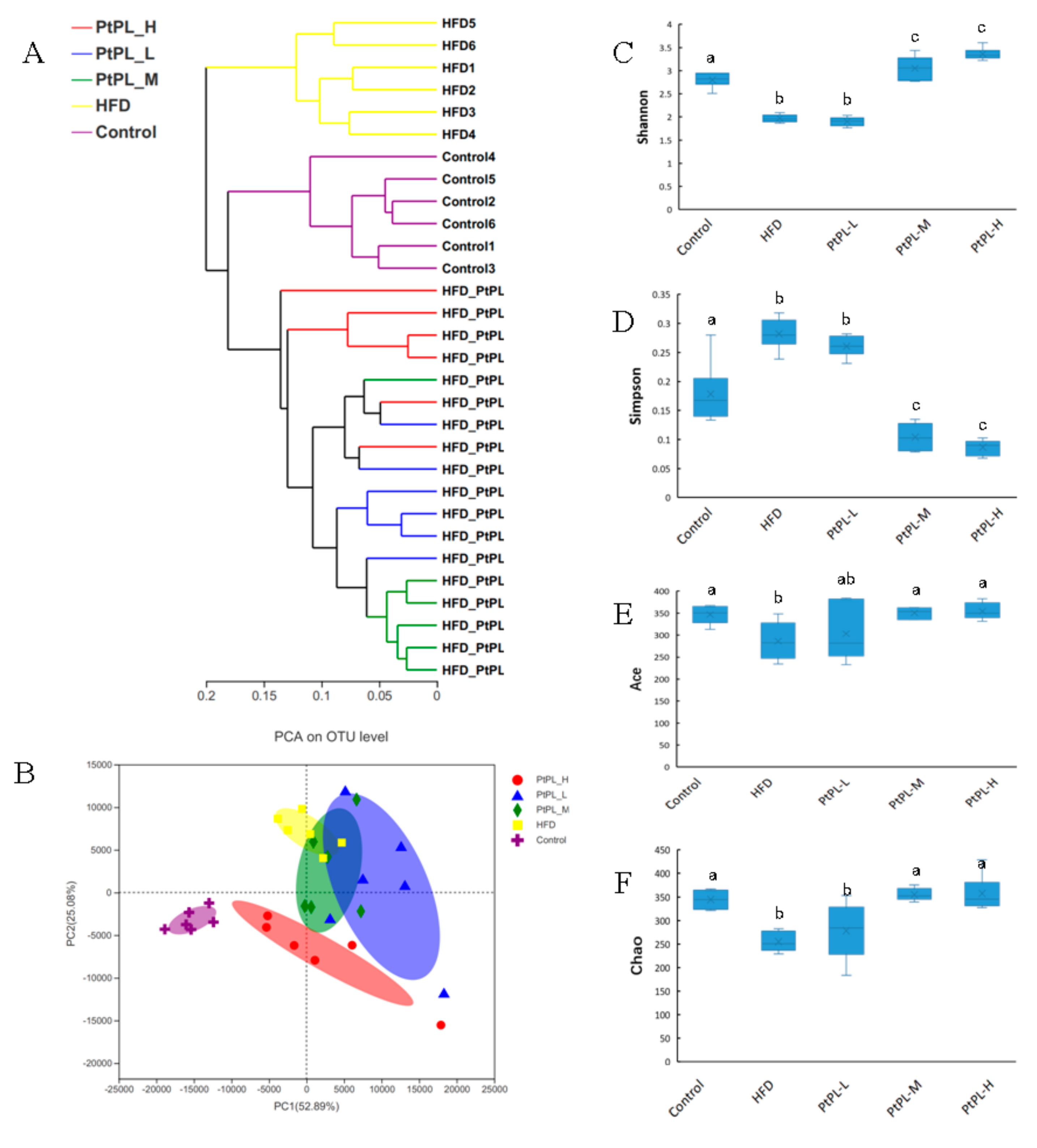
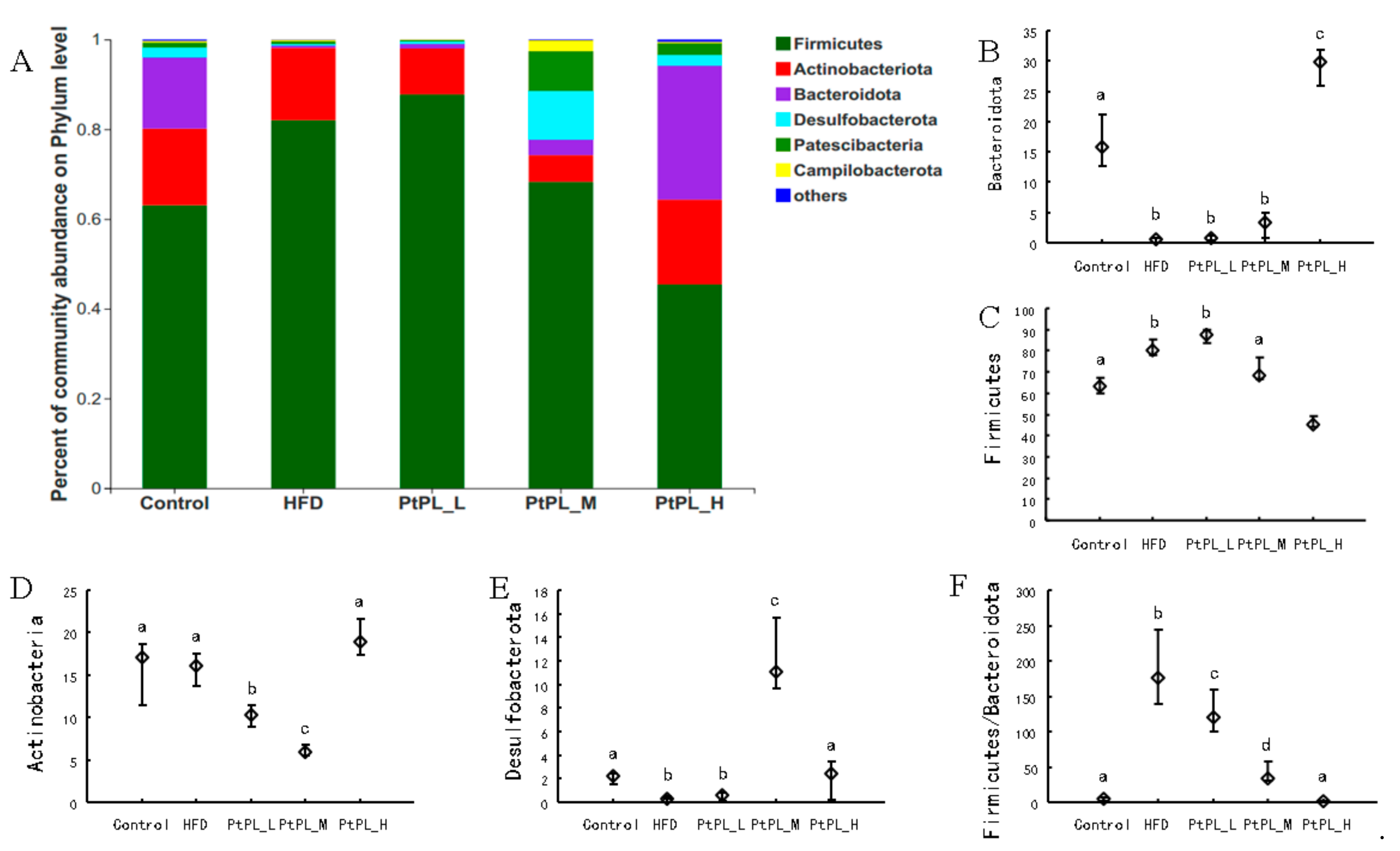
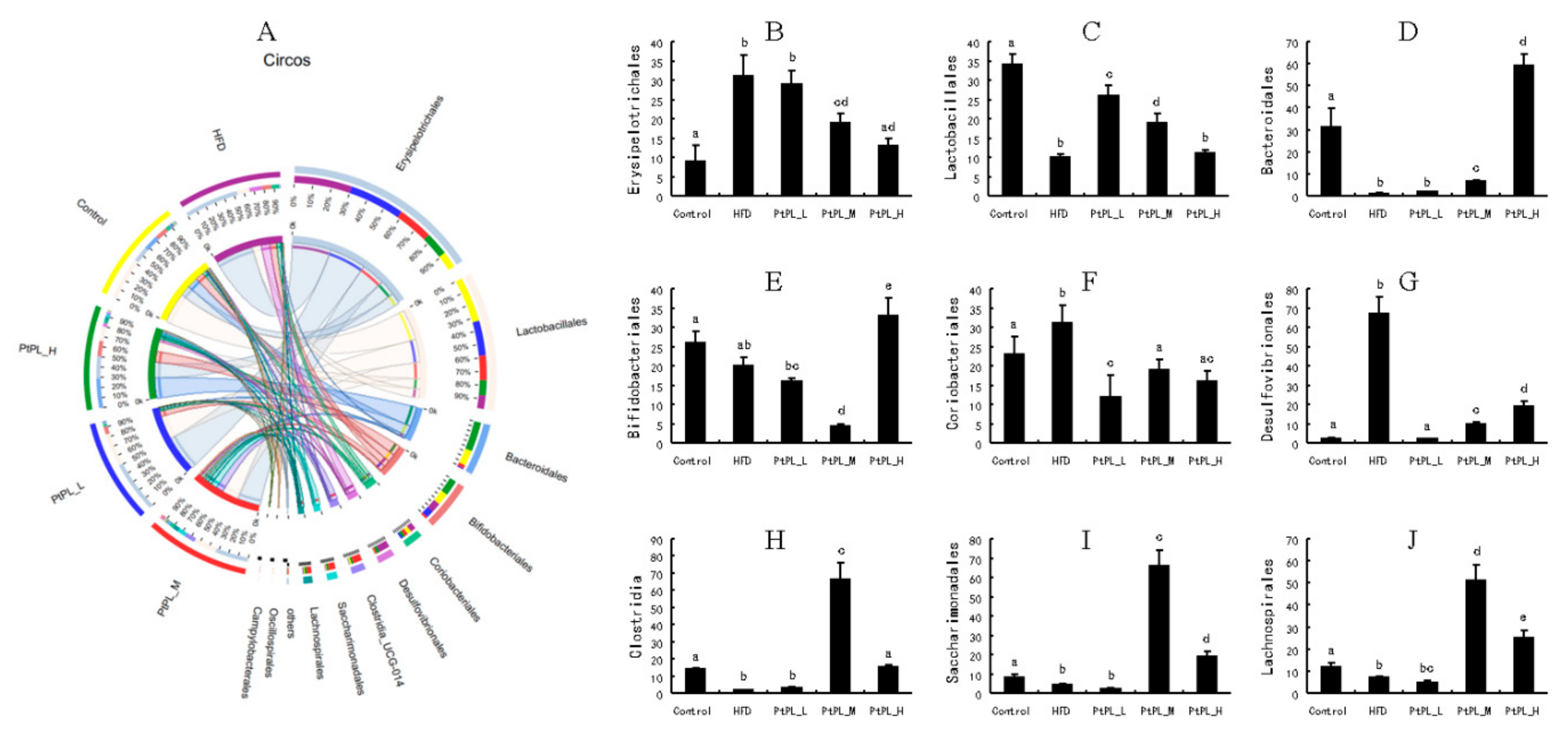
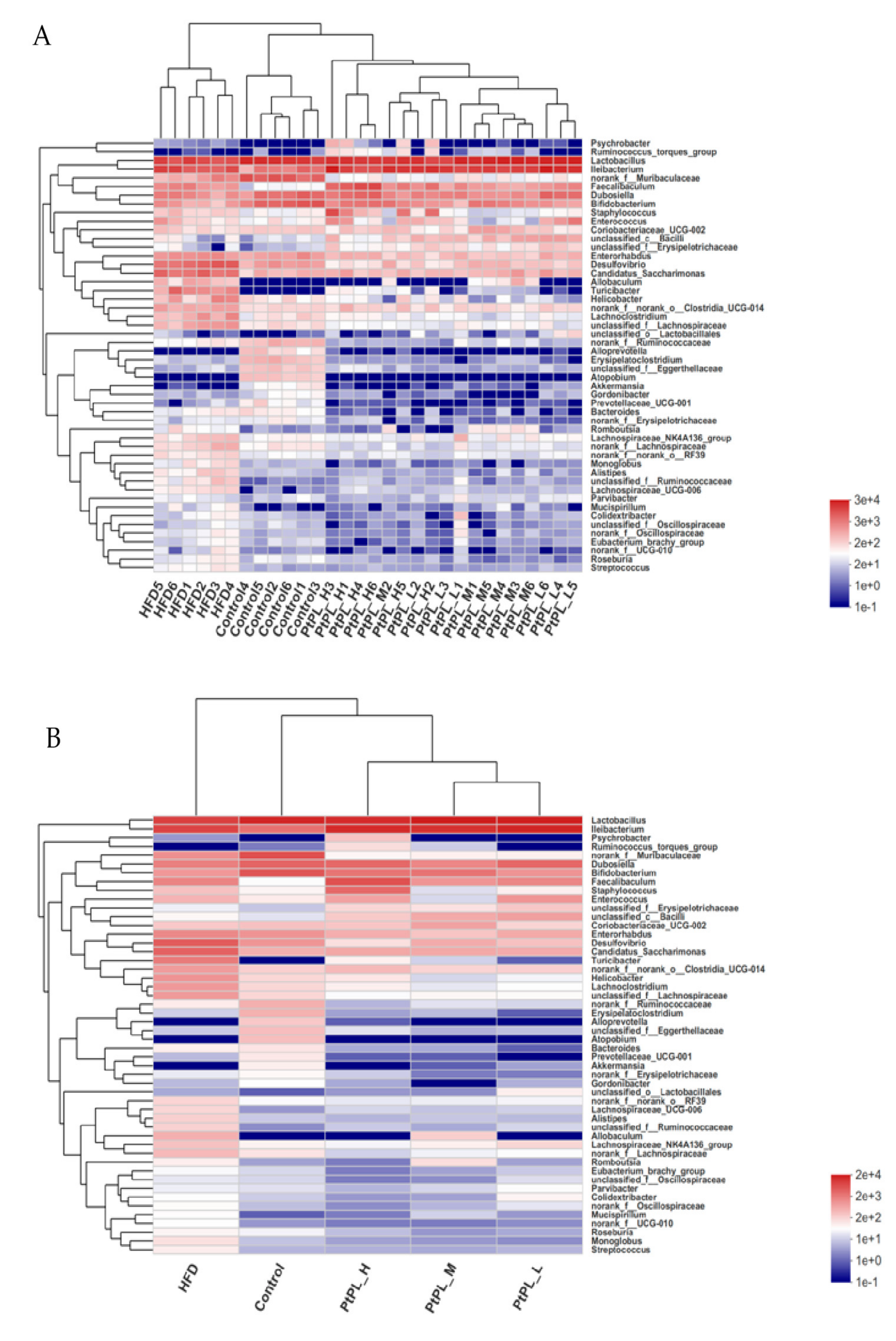
2.5. Effects of Pt-PL on Gut Microbiota
2.6. Effects of Pt-PL on LPS, Total Bile Acids, and SCFAs Concentrations
3. Discussion
4. Materials and Methods
4.1. Preparation of Pt-PL and Their Fatty Acids Composition
4.2. Animal Experiments
4.3. Blood Glucose, Lipids, Insulin, and Inflammatory Cytokines Test
4.4. Hepatic TG and TC Contents Detection
4.5. Hematoxylin and Eosin (H&E) and Oil Red O Staining
4.6. Gut Microbiota Analysis
4.7. Fecal LPS, Total Bile Acids, and SCFAs Determination
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Waise, T.M.Z.; Peppler, W.T.; Lam, T.K.T. The metabolic impact of small intestinal nutrient sensing. Nat. Commun. 2021, 12, 903. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Van Hul, M.; Knauf, C.; Cani, P.D. Gut microbiome, endocrine control of gut barrier function and metabolic diseases. J. Endocrinol. 2021, 248, R67–R82. [Google Scholar] [CrossRef]
- Lu, J.F.; Zhu, M.Q.; Zhang, H.; Liu, H.; Xia, B.; Wang, Y.L.; Shi, X.; Peng, L.; Wu, J.W. Neohesperidin attenuates obesity by altering the composition of the gut microbiota in high-fat diet-fed mice. FASEB J. 2020, 34, 12053–12071. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut microbiota and obesity: A role for probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes. 2018, 9, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking gut microbiota and inflammation to obesity and insulin resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; An, Y.; Tang, H.; Wang, Y. Alterations of bile acids and gut microbiota in obesity induced by high fat diet in rat model. J. Agric. Food Chem. 2019, 67, 3624–3632. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Zhou, X.; Chen, D.; Ouyang, G.; Liu, Y.; Cui, D. Periostin deficiency attenuates lipopolysaccharide- and obesity-induced adipose tissue fibrosis. FEBS Lett. 2021, 595, 2099–2112. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fan, C.; Li, P.; Lu, Y.; Chang, X.; Qi, K. Short chain fatty acids prevent high-fat-diet-induced obesity in mice by regulating G protein-coupled receptors and gut microbiota. Sci. Rep. 2016, 6, 37589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Bureau of Statistics of China. China Fishery Statistics Yearbook; National Bureau of Statistics of China: Beijing, China, 2021; pp. 22–38. (In Chinese) [Google Scholar]
- Ng’ambi, J.W.; Li, R.; Mu, C.; Song, W.; Liu, L.; Wang, C. Dietary administration of saponin stimulates growth of the swimming crab Portunus trituberculatus and enhances its resistance against Vibrio alginolyticus infection. Fish Shellfish Immunol. 2016, 59, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Hu, D.; Liu, M.; Hu, Y.; Liu, S. Molecular cloning and sequence analysis of two carbonic anhydrase in the swimming crab Portunus trituberculatus and its expression in response to salinity and pH stress. Gene 2016, 576, 347–357. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Yan, X.; Li, S.; Jiang, W.; Liu, Y. Egg oil from Portunus trituberculatus improves insulin resistance through activation of insulin signaling in mice. Appl. Physiol. Nutr. Metab. 2019, 44, 1081–1088. [Google Scholar] [CrossRef]
- Hu, S.; Yang, H.; Gao, X.; Li, S.; Jiang, W.; Liu, Y. Egg oil from Portunus trituberculatus alleviated obesity and regulated gut microbiota in mice. Sci. Rep. 2020, 10, 8454. [Google Scholar] [CrossRef]
- Hu, S.; Du, M.; Su, L.; Yang, H. Phosphatidylserine from Portunus trituberculatus eggs alleviates insulin resistance and alters the gut microbiota in high-fat-diet-fed mice. Mar. Drug. 2020, 18, 483. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of animal and marine origin: Structure, function, and anti-Inflammatory properties. Molecules 2017, 22, 1964. [Google Scholar] [CrossRef] [Green Version]
- Han, X.Q.; Zhang, L.Y.; Ding, L.; Shi, H.H.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Synergistic effect of sea cucumber saponins and EPA-enriched phospholipids on insulin resistance in high-fat diet-induced obese mice. Food Funct. 2019, 10, 3955–3964. [Google Scholar] [CrossRef]
- Xiang, X.; Zhou, X.; Wang, W.; Zhou, Y.; Zhou, X.; Deng, S.; Zheng, B.; Wen, Z. Effect of Antarctic krill phospholipid (KOPL) on high fat diet-induced obesity in mice. Food Res. Int. 2021, 148, 110456. [Google Scholar] [CrossRef]
- Li, X.; He, Q.; Hou, H.; Zhang, S.; Zhang, X.; Zhang, Y.; Wang, X.; Han, L.; Liu, K. Targeted lipidomics profiling of marine phospholipids from different resources by UPLC-Q-Exactive Orbitrap/MS approach. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1096, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, P.; Wang, F.; Zhang, S.; Li, H.; Zhang, Y.; Wang, X.; Liu, K.; Li, X. Separation, identification and cardiovascular activities of phospholipid classes from the head of Penaeus vannamei by lipidomics and zebrafish models. Food Funct. 2021, 12, 2282–2291. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Li, R.; Sun, H.; Zhang, M.; Cheng, W.; Chen, L.; Cheng, X.; Akoh, C.C. Phospholipids composition and molecular species of large yellow croaker (Pseudosciaena crocea) roe. Food Chem. 2018, 245, 806–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliu, F.; Leoni, B.; Pergola, R.D. Lipid classes and fatty acids composition of the roe of wild Silurus glanis from subalpine freshwater. Food Chem. 2017, 232, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, M.K.; Ahmmed, F.; Stewart, I.; Carne, A.; Tian, H.; Bekhit, A.E.A. Omega-3 phospholipids in Pacific blue mackerel (Scomber australasicus) processing by-products. Food Chem. 2021, 353, 129451. [Google Scholar] [CrossRef]
- Heymsfield, S.B.; Wadden, T.A. Mechanisms, pathophysiology, and management of obesity. N. Engl. J. Med. 2017, 376, 254. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wang, L.; Wen, C.; Yang, J.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber and its depolymerized derivative prevent obesity in association with modification of gut microbiota in high-fat diet-fed mice. Mol. Nutr. Food Res. 2018, 62, e1800446. [Google Scholar] [CrossRef]
- Lu, X.; Zhong, R.; Hu, L.; Huang, L.; Chen, L.; Cheng, W.; Zheng, B.; Liang, P. DHA-enriched phospholipids from large yellow croaker roe regulate lipid metabolic disorders and gut microbiota imbalance in SD rats with a high-fat diet. Food Funct. 2021, 12, 1825–4841. [Google Scholar] [CrossRef]
- Kroupova, P.; van Schothorst, E.M.; Keijer, J.; Bunschoten, A.; Vodicka, M.; Irodenko, I.; Oseeva, M.; Zacek, P.; Kopecky, J.; Rossmeisl, M.; et al. Omega-3 phospholipids from krill oil enhance intestinal fatty acid oxidation more effectively than omega-3 triacylglycerols in high-fat diet-fed obese mice. Nutritions 2020, 12, 2037. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The links between gut microbiota and obesity and obesity related diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, A.; Mujawdiya, P.K.; Pivina, L.; Doşa, A.; Semenova, Y.; Benahmed, A.G.; Bjørklund, G. Relationship between gut microbiota, gut hyperpermeability and obesity. Curr. Med. Chem. 2021, 28, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutritions 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Liu, J.; Yue, S.; Yang, Z.; Feng, W.; Meng, X.; Wang, A.; Peng, C.; Wang, C.; Yan, D. Oral hydroxysafflor yellow A reduces obesity in mice by modulating the gut microbiota and serum metabolism. Pharmacol. Res. 2018, 134, 40–50. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, N. Effects of metformin on the gut microbiota in obesity and type 2 diabetes mellitus. Diabetes Metab. Syndr. Obes. 2020, 13, 5003–5014. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, W.; Li, B.; Qian, S.; Wei, B.; Gong, S.; Wang, J.; Liu, M.; Wei, M. Nuciferine modulates the gut microbiota and prevents obesity in high-fat diet-fed rats. Exp. Mol. Med. 2020, 52, 1959–1975. [Google Scholar] [CrossRef]
- Guo, J.; Han, X.; Zhan, J.; You, Y.; Huang, W. Vanillin alleviates high fat diet-induced obesity and improves the gut microbiota composition. Front. Microbiol. 2018, 9, 2733. [Google Scholar] [CrossRef] [Green Version]
- Karrar, E.; Ahmed, I.A.M.; Huppertz, T.; Wei, W.; Jin, J.; Wang, X. Fatty acid composition and stereospecificity and sterol composition of milk fat from different species. Int. Dairy J. 2022, 128, 105313. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phospholipids | Proportion (%) | Phospholipids | Proportion (%) |
|---|---|---|---|
| LPC | 2.87 ± 0.15 | LPE | 0.27 ± 0.03 |
| LPA | 0.19 ± 0.03 | LPS | 0.07 ± 0.01 |
| PA | 19.61 ± 2.34 | PC | 32.28 ± 5.87 |
| PE | 8.81 ± 1.04 | PG | 0.22 ± 0.04 |
| PI | 7.96 ± 0.83 | PS | 26.51 ± 4.59 |
| SM | 0.87 ± 0.06 | Others | 0.49 ± 0.07 |
| CL | 0.07 ± 0.02 |
| Fatty acids | Proportion (%) | ||||
|---|---|---|---|---|---|
| PA | PC | PE | PI | PS [18] | |
| C15:0 | --- | --- | --- | --- | 0.59 ± 0.08 |
| C16:0 | --- | 0.62 ± 0.26 | --- | --- | 10.95 ± 1.57 |
| C16:1 n-9 | --- | 0.91 ± 0.68 | --- | --- | |
| C16:1 n-7 | --- | --- | 0.10 ± 0.06 | 0.83 ± 0.01 | |
| C17:0 | --- | --- | 0.03 ± 0.01 | --- | 5.18 ± 0.84 |
| C17:1 n-7 | --- | --- | --- | --- | 7.32 ± 0.86 |
| C17:3 n-9 | --- | --- | --- | --- | 0.12 ± 0.01 |
| C18:0 | 4.30 ± 1.03 | --- | --- | --- | 3.45 ± 0.61 |
| C18:1 n-9 | --- | 0.22 ± 0.03 | 1.77 ± 0.65 | --- | 1.16 ± 0.22 |
| C18:1 n-7 | --- | 0.80 ± 0.16 | 1.05 ± 0.34 | --- | 0.71 ± 0.14 |
| C18:2 n-6 | --- | 0.50 ± 0.01 | 0.11 ± 0.05 | 0.20 ± 0.07 | 0.59 ± 0.01 |
| C18:3 n-3 | --- | --- | --- | --- | 0.50 ± 0.02 |
| C20:0 | --- | 0.70 ± 0.13 | 0.12 ± 0.03 | --- | 2.01 ± 0.54 |
| C20:1 n-9 | --- | 1.43 ± 0.39 | 1.07 ± 0.36 | --- | 1.56 ± 0.39 |
| C20:2 n-6 | --- | 2.26 ± 0.19 | 0.34 ± 0.11 | --- | --- |
| C20:3 n-6 | 3.38 ± 1.01 | 1.01 ± 0.23 | 1.54 ± 0.99 | --- | 2.49 ± 0.27 |
| C20:4 n-6 | 20.11 ± 6.07 | 13.14 ± 2.80 | 8.27 ± 1.37 | 16.82 ± 3.36 | 0.44 ± 0.09 |
| C20:4 n-3 | 3.52 ± 0.56 | --- | --- | 20.83 ± 4.40 | 1.16 ± 0.25 |
| C20:5 n-3 | 51.84 ± 6.04 | 28.86 ± 6.01 | 53.76 ± 10.83 | 24.46 ± 5.31 | 18.70 ± 3.32 |
| C22:0 | --- | 0.58 ± 0.13 | --- | --- | --- |
| C22:1 n-9 | --- | 15.53 ± 3.67 | --- | --- | --- |
| C22:2 n-6 | 7.90 ± 4.71 | 2.41 ± 0.46 | 2.70 ± 0.56 | ||
| C22:3 n-6 | --- | 0.60 ± 0.16 | --- | --- | --- |
| C22:4 n-6 | --- | 2.14 ± 0.57 | 3.96 ± 1.13 | --- | --- |
| C22:5 n-6 | --- | 0.82 ± 0.19 | 6.54 ± 0.95 | --- | --- |
| C22:5 n-3 | --- | --- | 1.66 ± 0.33 | --- | --- |
| C22:6 n-3 | 8.95 ± 5.33 | 26.92 ± 5.60 | 19.48 ± 3.25 | 37.69 ± 4.79 | 30.43 ± 2.08 |
| C23:1 n-6 | --- | --- | --- | --- | 0.60 ± 0.11 |
| C23:3 n-6 | --- | 0.02 ± 0.00 | --- | --- | --- |
| Others | 0.00 ± 0.00 | 0.55 ± 0.00 | 0.21 ± 0.06 | 0.00 ± 0.00 | 8.52 ± 1.40 |
| ∑SFA | 4.30 ± 1.03 | 1.90 ± 0.17 | 0.15 ± 0.02 | 0.00 ± 0.00 | 22.19 ± 3.15 |
| ∑UFA | 95.70 ± 34.76 | 97.55 ± 0.84 | 99.64 ± 1.59 | 100 ± 8.96 | 69.29 ± 5.20 |
| ∑MUFA | 0.00 ± 0.00 | 18.89 ± 1.23 | 3.99 ± 0.44 | 0.00 ± 0.00 | 12.16 ± 2.24 |
| ∑PUFA | 95.70 ± 34.76 | 78.66 ± 1.07 | 95.65 ± 1.94 | 100 ± 8.96 | 57.13 ± 5.63 |
| ∑(EPA+DHA) | 60.79 ± 18.89 | 55.78 ± 5.81 | 73.24 ± 7.45 | 62.16 ± 5.35 | 50.13 ± 2.81 |
| Control | HFD | Pt-PL-L | Pt-PL-M | Pt-PL-H | |
|---|---|---|---|---|---|
| Food intake (g/w) | 29.74 ± 2.37 a | 25.43 ± 2.19 b | 25.62 ± 2.30 b | 25.18 ± 3.21 b | 25.35 ± 2.82 b |
| Calorie intake (kcal/w) | 121.79 ± 10.08 a | 166.82 ± 11.55 b | 168.07 ± 12.43 b | 165.18 ± 15.26 b | 166.29 ± 14.04 b |
| Body weight gain (g) | 11.44 ± 0.48 a | 21.55 ± 1.18 b | 18.97 ± 0.64 c | 14.96 ± 1.09 d | 14.55 ± 0.18 d |
| Perirenal fat weight (g) | 0.17 ± 0.03 a | 0.95 ± 0.08 b | 0.49 ± 0.07 c | 0.40 ± 0.08 cd | 0.25 ± 0.04 ad |
| Epididymal fat weight (g) | 0.49 ± 0.10 a | 4.09 ± 0.54 b | 1.84 ± 0.50 c | 1.17 ± 0.62 cd | 0.99 ± 0.11 d |
| Abdominal subcutaneous fat weight (g) | 0.23 ± 0.05 a | 1.17 ± 0.38 b | 0.73 ± 0.17 c | 0.56 ± 0.11 cd | 0.44 ± 0.10 d |
| Hepatic weight (g) | 1.01 ± 0.07 a | 2.11 ± 0.29 b | 1.63 ± 0.15 c | 1.21 ± 0.10 cd | 1.13 ± 0.12 ad |
| Fasting blood glucose (mmol/L) | 7.84 ± 0.38 a | 15.19 ± 0.67 b | 11.08 ± 0.50 c | 8.56 ± 0.46 d | 7.74 ± 0.37 d |
| Serum insulin (mU/L) | 26.00 ± 0.99 a | 32.64 ± 1.09 b | 28.79 ± 0.30 c | 26.60 ± 1.38 ad | 25.95 ± 2.05 ad |
| Serum TG (mmol/L) | 1.38 ± 0.05 a | 2.66 ± 0.12 b | 2.30 ± 0.13 b | 2.09 ± 0.06 c | 1.46 ± 0.09 a |
| Serum TC (mmol/L) | 3.79 ± 0.07 a | 7.25 ± 0.31 b | 5.66 ± 0.47 b | 4.54 ± 0.28 c | 4.06 ± 0.24 ac |
| Serum HDL-C (mmol/L) | 1.19 ± 0.20 a | 0.32 ± 0.04 b | 0.52 ± 0.05 c | 0.71 ± 0.11 d | 1.15 ± 0.06 a |
| Serum LDL-C (mmol/L) | 2.50 ± 0.20 a | 5.26 ± 0.37 b | 3.56 ± 0.30 c | 3.28 ± 0.29 c | 3.01 ± 0.14 ac |
| Hepatic TG (mg/g) | 20.56 ± 2.14 a | 36.28 ± 2.87 b | 30.01 ± 2.35 c | 28.28 ± 1.98 cd | 25.37 ± 2.17 d |
| Hepatic TC (mg/g) | 3.55 ± 0.28 a | 5.62 ± 0.41 b | 5.05 ± 0.39 bc | 4.46 ± 0.40 c | 4.38 ± 0.36 c |
| Serum TNF-α (pg/mL) | 56.08 ± 2.87 a | 106.01 ± 13.82 b | 94.73 ± 6.03 b | 76.99 ± 6.43 c | 56.94 ± 2.46 d |
| Serum IL-6 (pg/mL) | 57.60 ± 5.45 a | 83.46 ± 6.32 b | 66.77 ± 4.17 c | 60.04 ± 5.86 ac | 56.49 ± 4.31 a |
| Serum IL-1β (pg/mL) | 49.28 ± 3.00 a | 81.20 ± 4.27 b | 80.78 ± 4.16 b | 75.31 ± 5.36 b | 65.52 ± 2.79 c |
| Serum IL-10 (pg/mL) | 271.86 ± 8.90 a | 240.43 ± 7.74b | 248.74 ± 7.28 b | 254.15 ± 10.12 b | 272.16 ± 8.51 a |
| Parameters | Control | HFD | Pt-PL-L | Pt-PL-M | Pt-PL-H | |
|---|---|---|---|---|---|---|
| LPS (EU/g faeces) | 6.40 ± 0.67 a | 13.24 ± 0.98 b | 11.58 ± 1.04 c | 11.03 ± 0.75 c | 6.72 ± 0.54 a | |
| Total bile acids (µmol/g faeces) | 6.57 ± 0.62 a | 13.34 ± 0.85 b | 9.14 ± 0.76 c | 8.24 ± 0.59 c | 6.89 ± 0.71 a | |
| SCFAs (mg/g faeces) | Acetate | 17.63 ± 1.44 a | 6.09 ± 0.89 b | 6.08 ± 0.77 b | 14.51 ± 1.14 c | 14.93 ± 1.34 c |
| Propionate | 5.03 ± 0.34 a | 0.62 ± 0.05 b | 0.67 ± 0.05 b | 1.40 ± 0.09 | 2.35 ± 0.21 d | |
| Butyrate | 3.69 ± 0.31 a | 0.21 ± 0.01 b | 0.42 ± 0.02 c | 1.80 ± 0.12 c | 8.30 ± 0.87 d | |
| Isobutyrate | 0.33 ± 0.02 a | 0.02 ± 0.00 b | 0.08 ± 0.01 c | 0.24 ± 0.02 c | 0.35 ± 0.03 a | |
| Valerate | 0.61 ± 0.04 a | 0.04 ± 0.01 b | 0.04 ± 0.00 b | 0.08 ± 0.00 c | 0.25 ± 0.01 d | |
| Isovalerate | 0.42 ± 0.03 a | 0.12 ± 0.00 b | 0.11 ± 0.01 b | 0.29 ± 0.02 c | 0.28 ± 0.01 c | |
| Hexanoate | 0.10 ± 0.00 a | ND | ND | 0.01 ± 0.00 b | 0.02 ± 0.00 c | |
| Total SCFAs | 27.76 ± 2.04 a | 7.09 ± 0.63 b | 7.41 ± 0.65 b | 18.33 ± 1.70 c | 26.49 ± 2.18 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, L.; Zhu, H.; Chen, S.; Du, M.; Wan, X.; Liu, Y.; Hu, S.; Xu, Y. Anti-Obesity and Gut Microbiota Regulation Effects of Phospholipids from the Eggs of Crab, Portunus Trituberculatus, in High Fat Diet-Fed Mice. Mar. Drugs 2022, 20, 411. https://doi.org/10.3390/md20070411
Su L, Zhu H, Chen S, Du M, Wan X, Liu Y, Hu S, Xu Y. Anti-Obesity and Gut Microbiota Regulation Effects of Phospholipids from the Eggs of Crab, Portunus Trituberculatus, in High Fat Diet-Fed Mice. Marine Drugs. 2022; 20(7):411. https://doi.org/10.3390/md20070411
Chicago/Turabian StyleSu, Laijin, Hongli Zhu, Sichun Chen, Mengyu Du, Xiaofeng Wan, Yishu Liu, Shiwei Hu, and Yangli Xu. 2022. "Anti-Obesity and Gut Microbiota Regulation Effects of Phospholipids from the Eggs of Crab, Portunus Trituberculatus, in High Fat Diet-Fed Mice" Marine Drugs 20, no. 7: 411. https://doi.org/10.3390/md20070411
APA StyleSu, L., Zhu, H., Chen, S., Du, M., Wan, X., Liu, Y., Hu, S., & Xu, Y. (2022). Anti-Obesity and Gut Microbiota Regulation Effects of Phospholipids from the Eggs of Crab, Portunus Trituberculatus, in High Fat Diet-Fed Mice. Marine Drugs, 20(7), 411. https://doi.org/10.3390/md20070411