
Extracellular Polymeric Substances Produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and Assessment of Their Antioxidant and Metal-Chelating Activities




Abstract
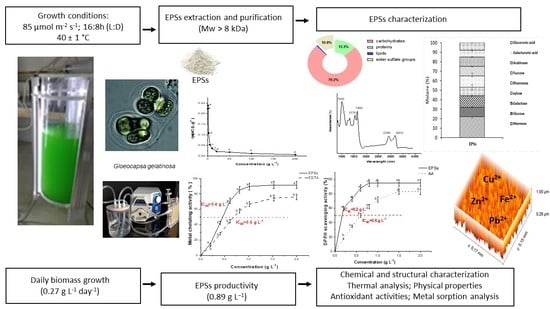
:
1. Introduction
2. Results and discussion
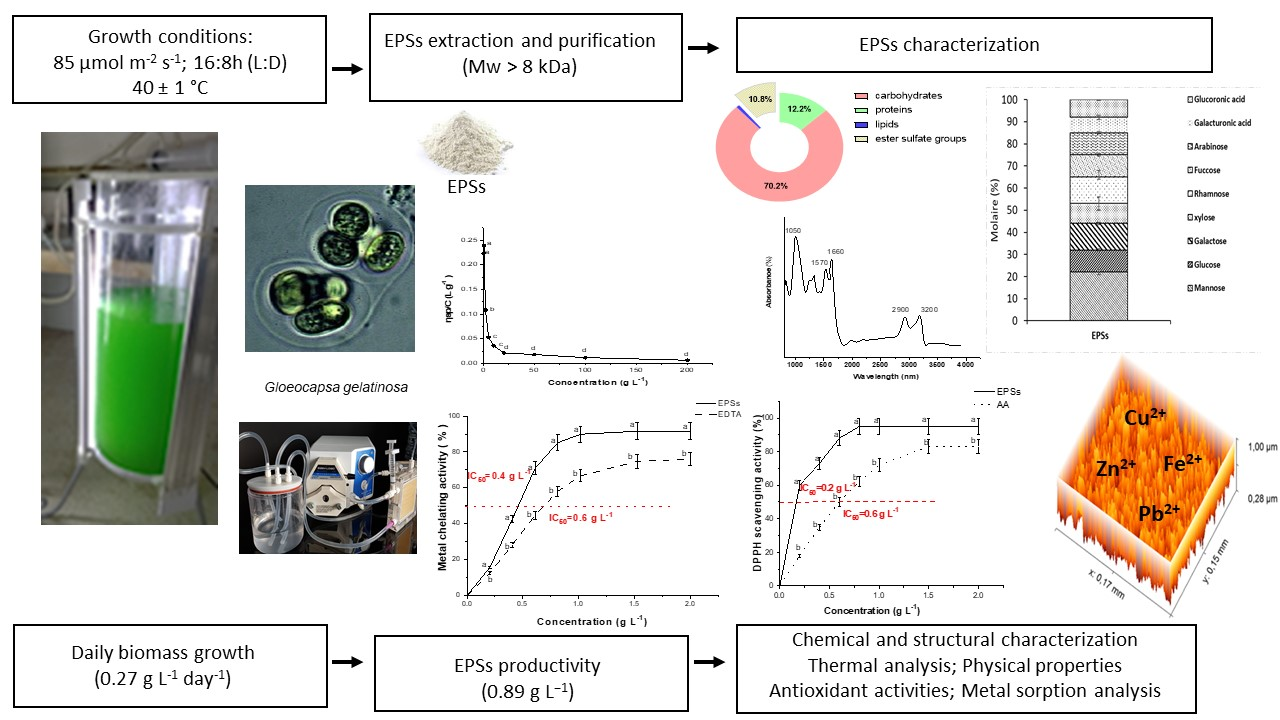
2.1. G. gelatinosa Biomass and EPSs Production
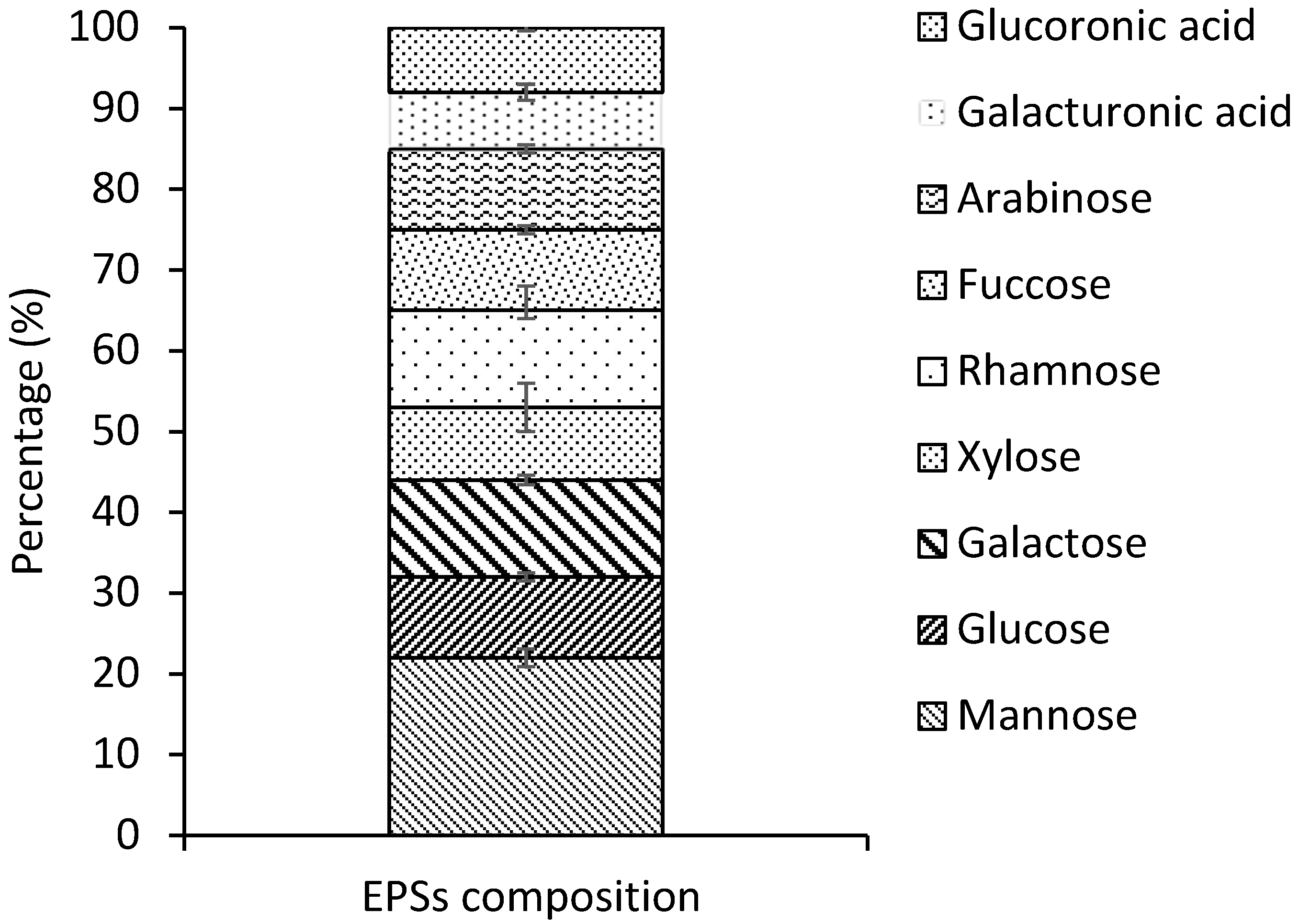
2.2. Chemical Composition Analysis and Monosaccharide’s Composition of G. gelatinosa EPSs
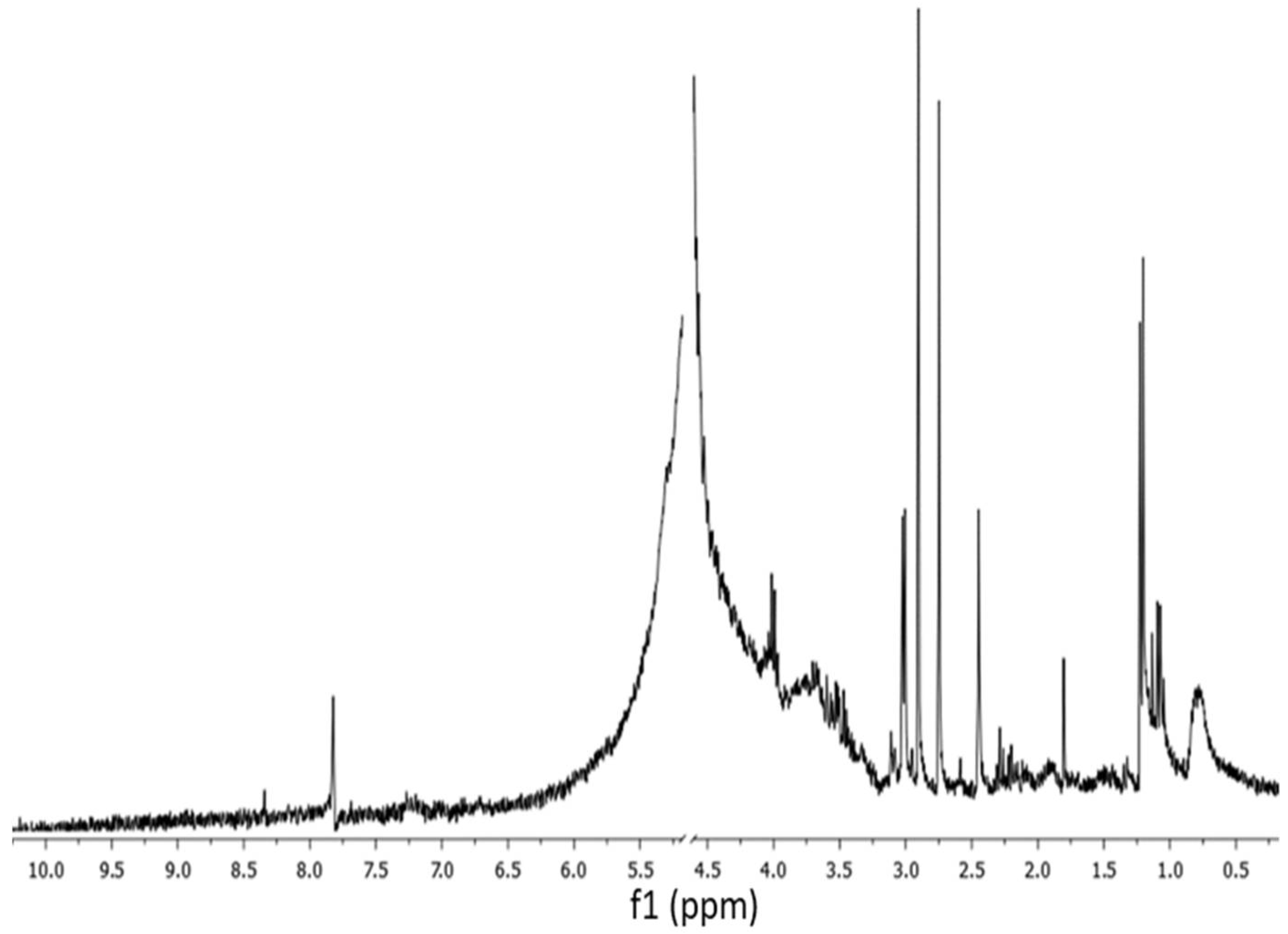
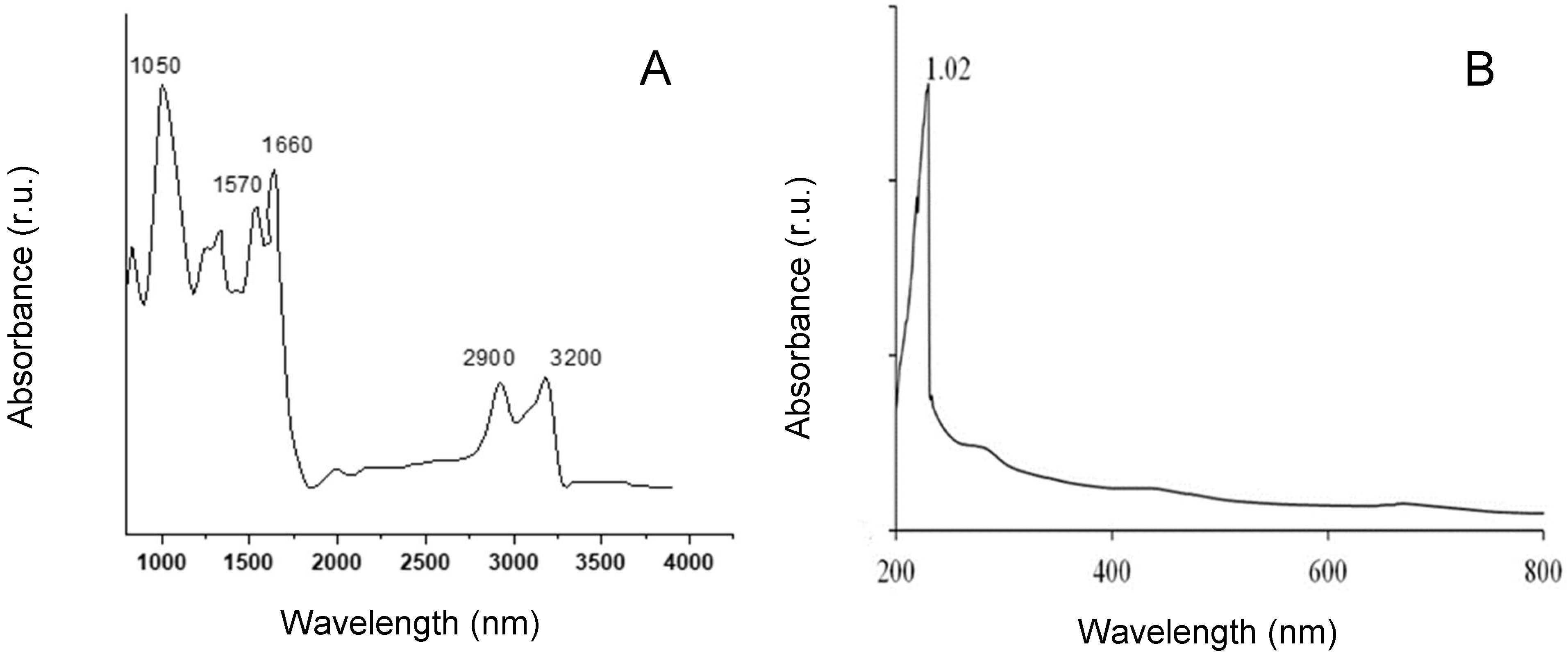
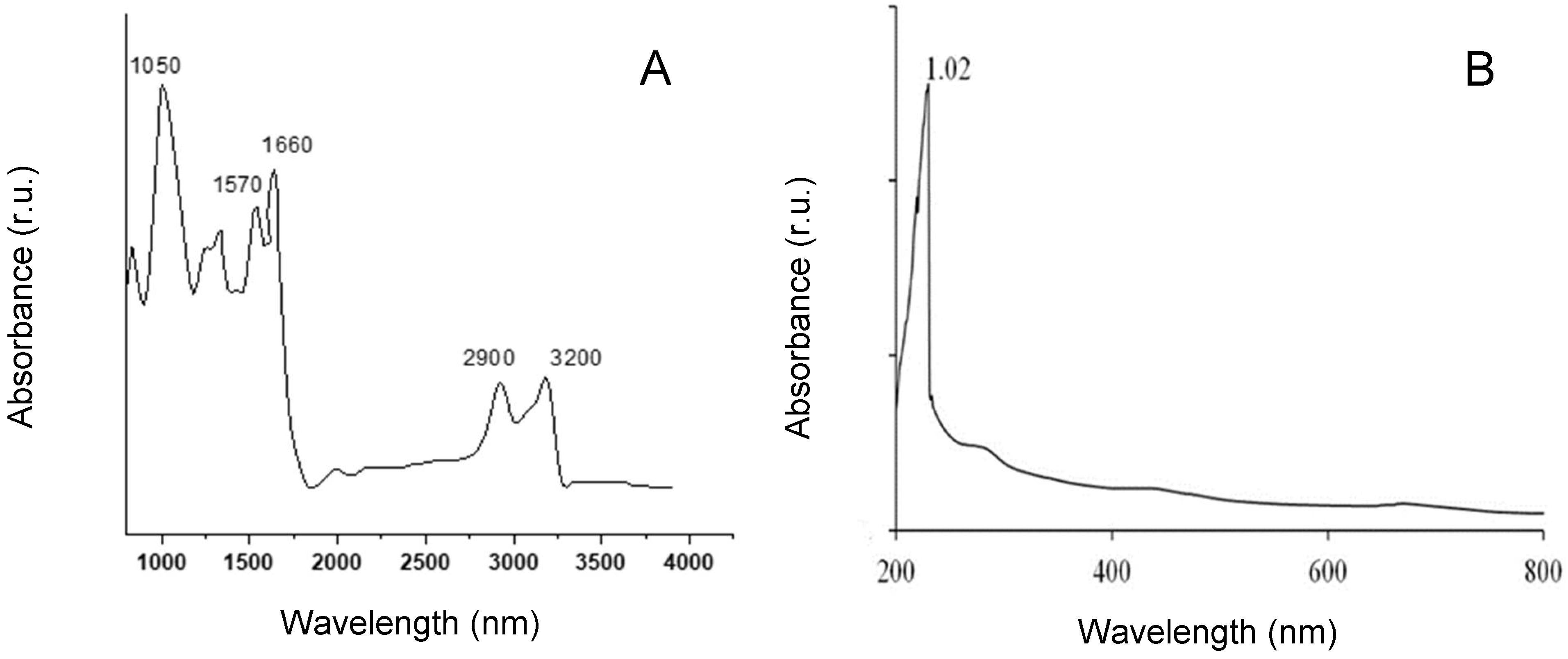
2.3. 1H NMR and FTIR Spectra of G. gelatinosa EPSs
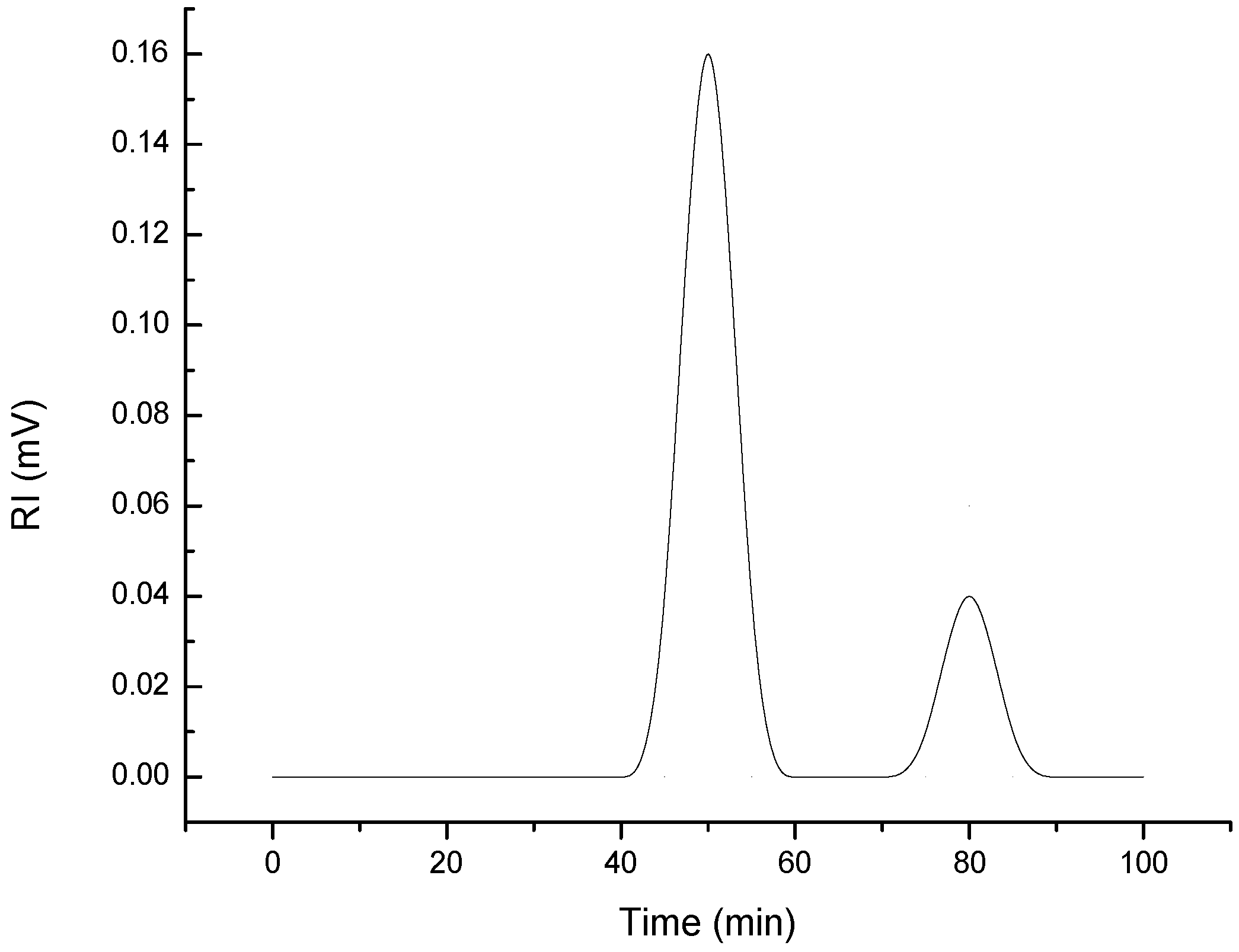
2.4. Molecular Weight and Zeta-Potential of G. gelatinosa EPSs
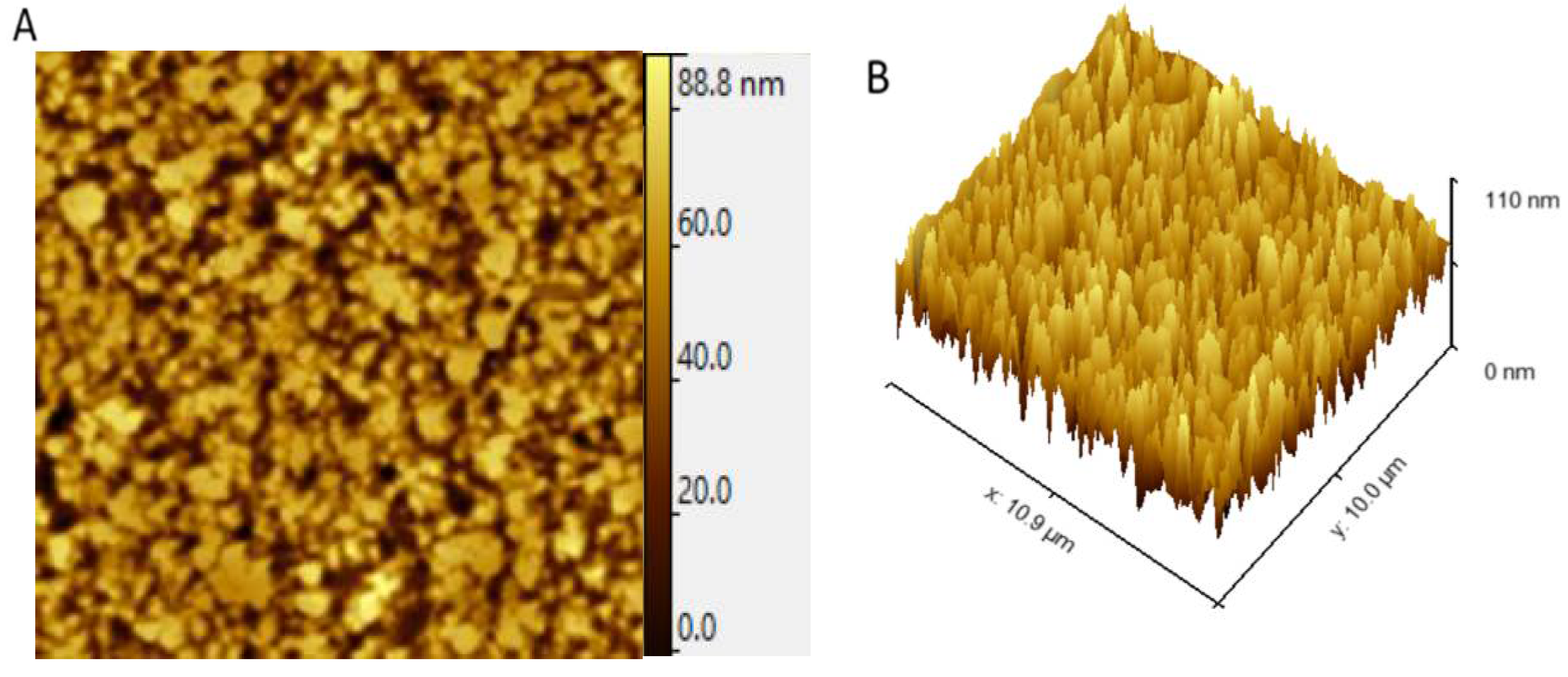
2.5. Atomic Force Microscopy Analysis of G. gelatinosa EPSs
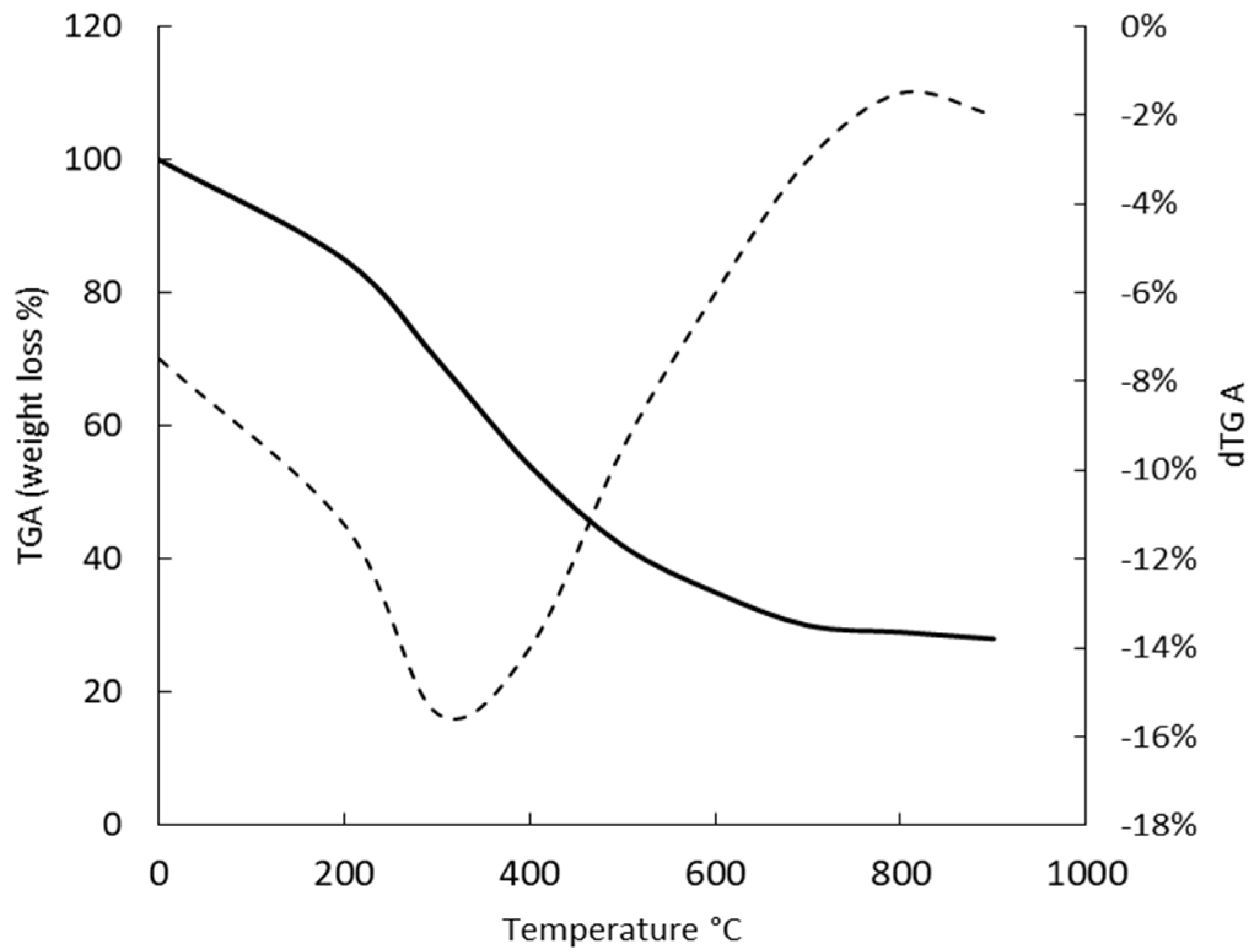
2.6. Thermal Properties of G. gelatinosa EPSs
2.7. WHC and WSI Analysis
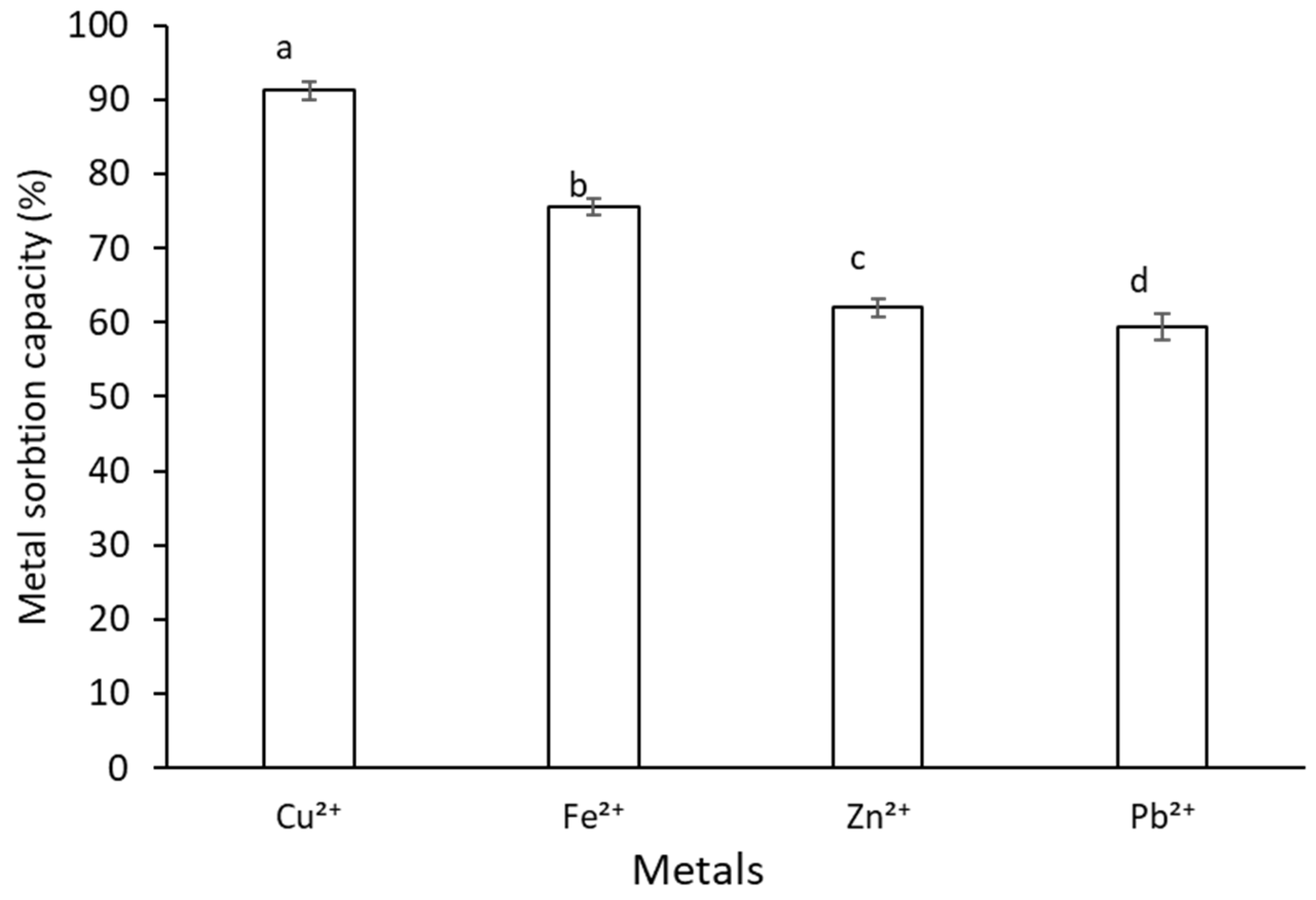
2.8. Metal Sorption Activity of G. gelatinosa EPSs
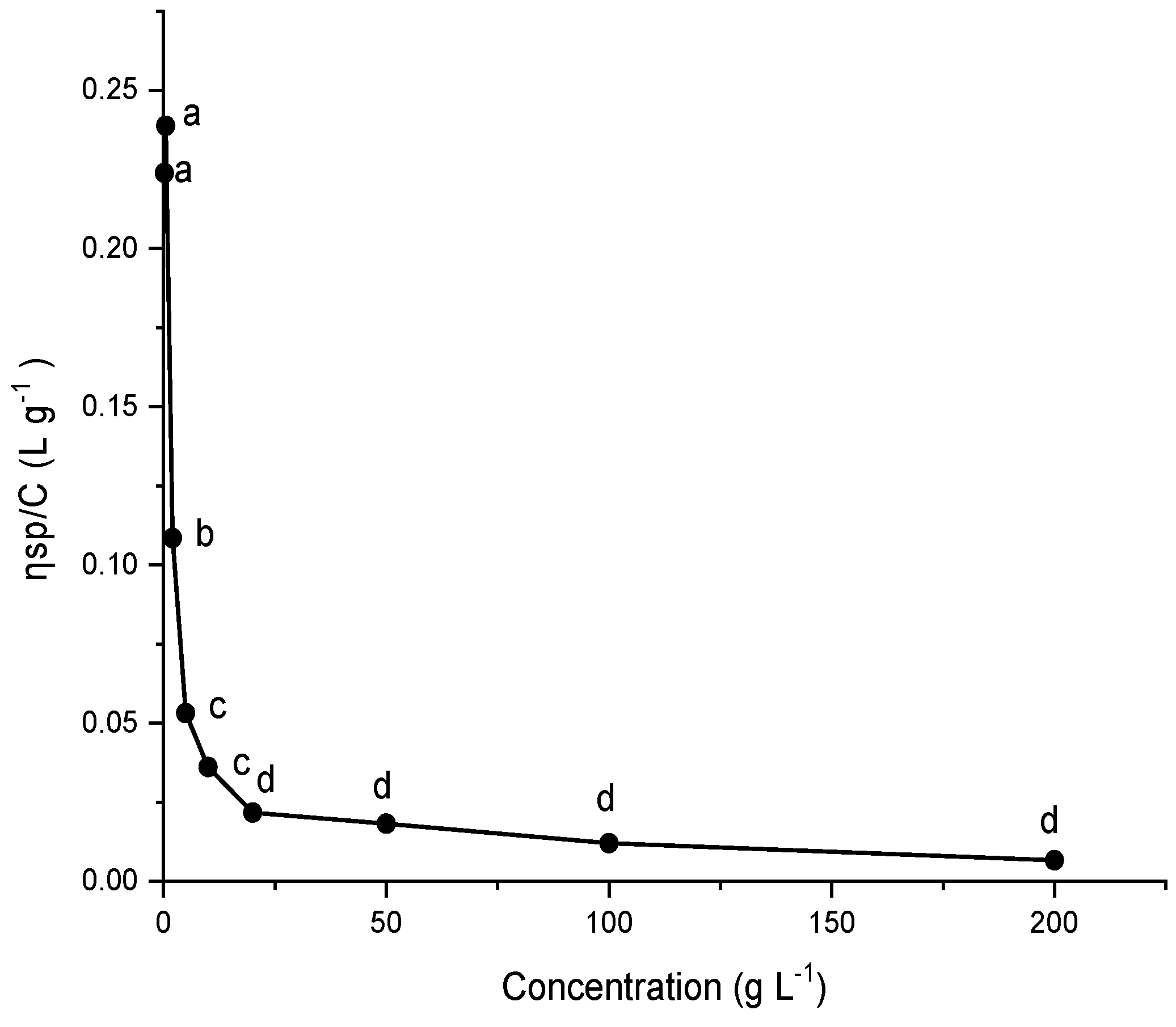
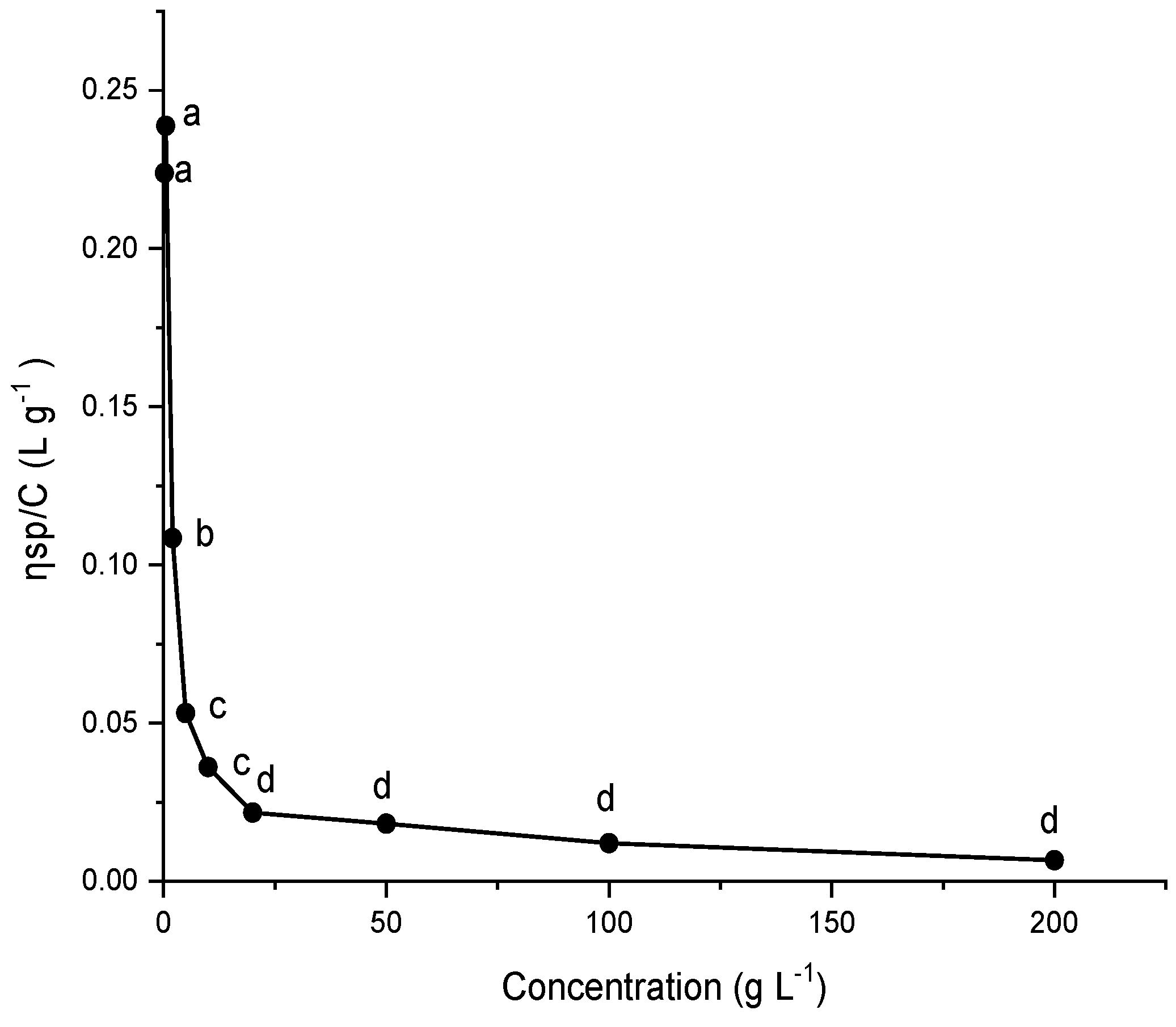
2.9. Viscosity of G. gelatinosa EPSs
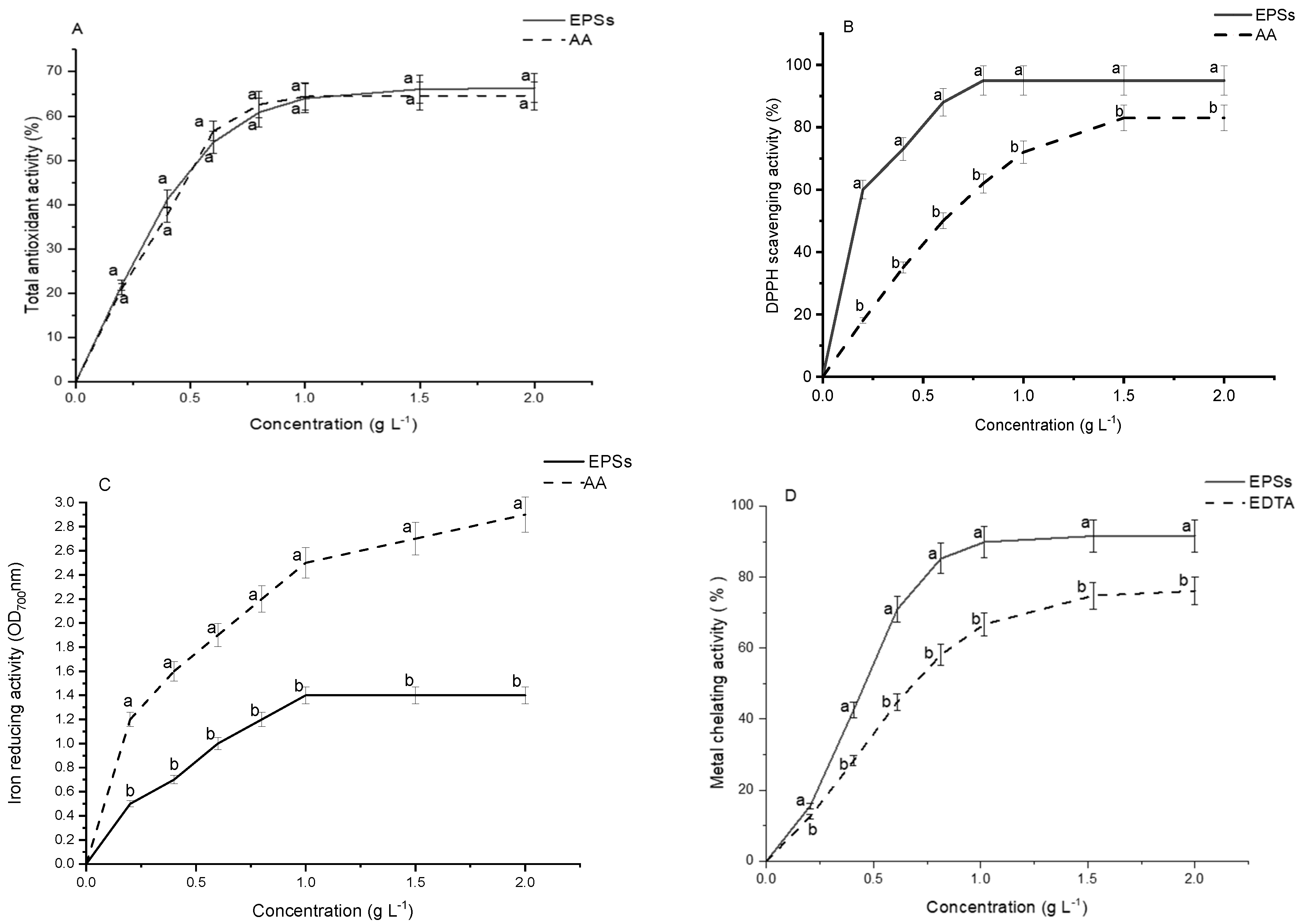
2.10. Antioxidant Capacity of G. gelatinosa EPSs
3. Materials and Methods
3.1. Organism and Culture Conditions
3.2. Extraction and Purification of Extracellular Polymeric Substances
3.3. Biochemical Composition and Monosaccharide Profile
3.4. Proton Nuclear Magnetic Resonance Spectroscopy Analysis
3.5. Fourier Transform Infrared Spectroscopy and UV-Visible Spectroscopy Analyse
3.6. Determination of Molecular Weight
3.7. Zeta Potential
3.8. Atomic Force Microscopy Analysis
3.9. Thermal Gravimetric Analysis
3.10. Water-Holding Capacity and Water Solubility Indexes
3.11. Metal Sorption Activity
3.12. Viscosity Measurements
3.13. Screening of the Antioxidant Capacity
3.13.1. Total Antioxidant Activity
3.13.2. α,α-diphenyl-β-picrylhydrazyl Radical Scavenging Activity
3.13.3. Iron-Reducing Activity
3.13.4. Metal-Chelating Activity
3.14. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Decho, A.W. Biochemical Characterization of Cyanobacterial Extracellular Polymers (EPS) from Modern Marine Stromatolites (Bahamas). Prep. Biochem. Biotechnol. 2000, 30, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Gongi, W.; Cordeiro, N.; Pinchetti, J.L.G.; Sadok, S.; Ben Ouada, H. Extracellular polymeric substances with high radical scavenging ability produced in outdoor cultivation of the thermotolerant chlorophyte Graesiella sp. J. Appl. Phycol. 2021, 33, 357–369. [Google Scholar] [CrossRef]
- Bhatnagar, M.; Parwani, L.; Sharma, V.; Ganguly, J.; Bhatnagar, A. Exopolymers from Tolypothrix tenuis and three Anabaena sp. (Cyanobacteriaceae) as novel blood clotting agents for wound management. Carbohydr. Polym. 2014, 99, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.; Fidelis, G.P.; Cordeiro, S.; Oliveira, R.; Sabry, D.; Câmara, R.; Nobre, L.; Costa, M.; Almeida-Lima, J.; Farias, E.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Parwani, L.; Bhatnagar, M.; Bhatnagar, A.; Sharma, V. Antioxidant and iron-chelating activities of cyanobacterial exopolymers with potential for wound healing. J. Appl. Phycol. 2014, 26, 1473–1482. [Google Scholar] [CrossRef]
- Abed, R.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef]
- Pereira, S.B.; Sousa, A.; Santos, M.; Araújo, M.; Serôdio, F.; Granja, P.; Tamagnini, P. Strategies to Obtain Designer Polymers Based on Cyanobacterial Extracellular Polymeric Substances (EPS). Int. J. Mol. Sci. 2019, 20, 5693. [Google Scholar] [CrossRef] [Green Version]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Reisser, W. Polyextremophilic Photoautotrophic Eukaryotic Algae. In Polyextremophiles. Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Oren, A., Stan-Lotter, H., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 27, pp. 567–580. [Google Scholar] [CrossRef]
- Ramachandra, T.V.; Mahapatra, D.M.; Karthick, B.; Gordon, R. Milking Diatoms for Sustainable Energy: Biochemical Engineering versus Gasoline-Secreting Diatom Solar Panels. Ind. Eng. Chem. Res. 2009, 48, 8769–8788. [Google Scholar] [CrossRef]
- Mezhoud, N.; Zili, F.; Bouzidi, N.; Helaoui, F.; Ammar, J.; Ben Ouada, H. The effects of temperature and light intensity on growth, reproduction and EPS synthesis of a thermophilic strain related to the genus Graesiella. Bioprocess Biosyst. Eng. 2014, 37, 2271–2280. [Google Scholar] [CrossRef] [PubMed]
- Ghozzi, K.; Zemzem, M.; Ben Dhiab, R.; Challouf, R.; Yahia, A.; Omrane, H.; Ben Ouada, H. Screening of thermophilic microalgae and cyanobacteria from Tunisian geothermal sources. J. Arid Environ. 2013, 97, 14–17. [Google Scholar] [CrossRef]
- Bleeke, F.; Rwehumbiza, V.M.; Winckelmann, D.; Klöck, G. Isolation and Characterization of New Temperature Tolerant Microalgal Strains for Biomass Production. Energies 2014, 7, 7847–7856. [Google Scholar] [CrossRef] [Green Version]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Baker, K.K.; Baldwin, C.L.; Bolis, C.M.; Caplan, F.R.; Larsen, L.K.; Levine, I.A.; Moore, R.E.; Nelson, C.S.; Tschappat, K.D.; et al. Antiviral activity of cultured blue-green algae. Cyanophyta 1993, 130, 125–130. [Google Scholar]
- Gacheva, G.; Gigova, L.; Ivanova, N.; Iliev, I.; Toshkova, R.; Gardeva, E.; Kussovski, V.; Najdenski, H. Suboptimal growth temperatures enhance the biological activity of cultured cyanobacterium Gloeocapsa sp. J. Appl. Phycol. 2012, 25, 183–194. [Google Scholar] [CrossRef]
- Chentir, I.; Hamdi, M.; Doumandji, A.; HadjSadok, A.; Ouada, H.B.; Nasri, M.; Jridi, M. Enhancement of extracellular polymeric substances (EPS) production in Spirulina (Arthrospira sp.) by two-step cultivation process and partial characterization of their polysaccharidic moiety. Int. J. Biol. Macromol. 2017, 105, 1412–1420. [Google Scholar] [CrossRef]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Paulsen, B.S.; Klaveness, D. Studies on Polysaccharides from Three Edible Species of Nostoc (Cyanobacteria) with Different Colony Morphologies: Comparison of Monosaccharide Compositions and Viscosities of Poly-Saccharides from Field Colonies and Suspension Cultures. J. Phycol. 1998, 34, 962–968. [Google Scholar] [CrossRef]
- Pereira, S.; Zille, A.; Micheletti, E.; Moradas-Ferreira, P.; De Philippis, R.; Tamagnini, P. Complexity of cyanobacterial exopolysaccharides: Composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 2009, 33, 917–941. [Google Scholar] [CrossRef]
- Micheletti, E.; Pereira, S.; Mannelli, F.; Moradas-Ferreira, P.; Tamagnini, P.; De Philippis, R. Sheathless Mutant of Cyanobacterium Gloeothece sp. Strain PCC 6909 with Increased Capacity To Remove Copper Ions from Aqueous Solutions. Appl. Environ. Microbiol. 2008, 74, 2797–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Kavita, K.; Jha, B. Characterization of extracellular polymeric substances produced by micro-algae Dunaliella salina. Carbohydr. Polym. 2011, 83, 852–857. [Google Scholar] [CrossRef]
- Santhiya, D.; Subramanian, S.; Natarajan, K. Surface Chemical Studies on Sphalerite and Galena Using Extracellular Polysaccharides Isolated from Bacillus polymyxa. J. Colloid Interface Sci. 2002, 256, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, M.; Nucara, A. Secondary structure of food proteins by Fourier transform spectroscopy in the mid-infrared region. Amino Acids 2010, 38, 679–690. [Google Scholar] [CrossRef]
- Spirli, H.; Sagdic, O.; Yılmaz, M.T.; Dertli, E. Physicochemical characterization of an α-glucan from Lactobacillus reuteri E81 as a potential exopolysaccharide suitable for food applications. Process. Biochem. 2019, 79, 91–96. [Google Scholar] [CrossRef]
- Alasonati, E.; Slaveykova, V.I. And Composition and molar mass characterisation of bacterial extracellular polymeric substances by using chemical, spectroscopic and fractionation techniques. Environ. Chem. 2011, 8, 155. [Google Scholar] [CrossRef]
- Chi, Y.; Ye, H.; Li, H.; Li, Y.; Guan, H.; Mou, H.; Wang, P. Structure and molecular morphology of a novel moisturizing exopolysaccharide produced by Phyllobacterium sp. 921F. Int. J. Biol. Macromol. 2019, 135, 998–1005. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Jiang, Y.-Y.; Zhao, X.; Hao, X.N.; Li, L.; Yang, Z.-N. Characterization and Antioxidant Activity of the Exopolysaccharide Produced by Bacillus amyloliquefaciens GSBa-J. Microbiol. Biotechnol. 2018, 28, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Angel, S.J.; Vidyadharani, G.; Santhosh, S.; Dhandapani, R. Optimization and characterisation of thermo stable exopolysac-charide produced from Bacillus licheniformis WSF-1 strain. J. Polym. Environ. 2018, 26, 3824–3833. [Google Scholar] [CrossRef]
- Zohuriaan-Mehr, M.J.; Shokrolahi, F. Thermal studies on natural and modified gums. Polym. Test. 2004, 23, 575–579. [Google Scholar] [CrossRef]
- Chowdhury, S.; Manna, S.; Saha, P.; Basak, R.; Sen, R.; Roy, D.; Adhikari, B. Composition analysis and material characterization of an emulsifying extracellular polysaccharide (EPS) produced by Bacillus megaterium RB-05: A hydrodynamic sediment-attached isolate of freshwater origin. J. Appl. Microbiol. 2011, 111, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Vinothini, G.; Latha, S.; Arulmozhi, M.; Dhanasekaran, D. Statistical optimization, physio-chemical and bio-functional attributes of a novel exopolysaccharide from probiotic Streptomyces griseorubens GD5. Int. J. Biol. Macromol. 2019, 134, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Zhu, L.; Chen, X.; Wang, P.G.; Zhang, Y. Structural characterization and ecological roles of a novel exopolysaccharide from the deep-sea psychrotolerant bacterium Pseudoalteromonas sp. SM9913. Microbiology 2007, 153, 1566–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gongi, W.; Cordeiro, N.; Pinchetti, J.L.G.; Ben Ouada, H. Functional, rheological, and antioxidant properties of extracellular polymeric substances produced by a thermophilic cyanobacterium Leptolyngbya sp. J. Appl. Phycol. 2022, 1007, 1573–5176. [Google Scholar] [CrossRef]
- Yang, H.; Zhu, P.; Peng, C.; Ma, S.; Zhu, Q.; Fan, C. Viscometric study of polyvinyl alcohol in NaCl/water solutions ranged from dilute to extremely dilute concentration. Eur. Polym. J. 2001, 37, 1939–1942. [Google Scholar] [CrossRef]
- Lan, M. Antioxidant and anti-tumor activities of purified polysaccharides with low molecular weights from Magnolia officinalis. J. Med. Plants Res. 2012, 6, 1025–1034. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2020, 487, 107881. [Google Scholar] [CrossRef]
- Zili, F.; Mezhoud, N.; Trabelsi, L.; Chreif, I.; Ben Ouada, H. Fatty acid composition of the thermophilic Gloeocapsa gelatinosa under different combinations of temperature, light intensity, and NaNO3 concentration. J. Appl. Phycol. 2015, 27, 97–107. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Craigie, J.S.; Wen, Z.C.; Van der Meer, J.P. Interspecific and nutritionally-determination variations in the composition of agars from Gracilaria sp. Bot. Mar. 1984, 27, 55–61. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Åman, P.; McNeil, M.; Franzén, L.-E.; Darvill, A.G.; Albersheim, P. Structural elucidation, using h.p.l.c.-m.s. and g.l.c.-m.s., of the acidic polysaccharide secreted by rhizobium meliloti strain 1021. Carbohydr. Res. 1981, 95, 263–282. [Google Scholar] [CrossRef]
- Streeter, J.G.; Strimbu, C. Simultaneous Extraction and Derivatization of Carbohydrates from Green Plant Tissues for Analysis by Gas–Liquid Chromatography. Anal. Biochem. 1998, 259, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; You, S. Molecular Characteristics of Water-Soluble Extracts from Hypsizigus marmoreusand Their in Vitro Growth Inhibition of Various Cancer Cell Lines and Immunomodulatory Function in Raw 264.7 Cells. Biosci. Biotechnol. Biochem. 2011, 75, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Q.; Li, M.-M.; Chen, T.-H.; Zhou, Y.-F.; Yue, Z.-B. Competitive adsorption of heavy metal by extracellular polymeric substances (EPS) extracted from sulfate-reducing bacteria. Bioresour. Technol. 2014, 163, 374–376. [Google Scholar] [CrossRef]
- Dapkevicius, A.; Venskutonis, R.; Van Beek, T.A.; Linssen, J.P.H. Antioxidant activity of extracts obtained by different isolation procedures from some aromatic herbs grown in Lithuania. J. Sci. Food Agric. 1998, 77, 140–146. [Google Scholar] [CrossRef]
- Bersuder, P.; Hole, M.; Smith, G. Antioxidants from a heated histidine-glucose model system. I: Investigation of the antioxidant role of histidine and isolation of antioxidants by high-performance liquid chromatography. J. Am. Oil Chem. Soc. 1998, 75, 181–187. [Google Scholar] [CrossRef]
- Adjimani, J.P.; Asare, P. Antioxidant and free radical scavenging activity of iron chelators. Toxicol. Rep. 2015, 2, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Wettasinghe, M.; Shahidi, F. Iron (II) chelation activity of extracts of borage and evening primrose meals. Food Res. Int. 2002, 35, 65–71. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipids | Carbohydrates | Proteins | Ester Sulfate | C→N | H→S→P |
|---|---|---|---|---|---|
| 0.8 ± 0.1 | 70.2 ± 0.1 | 12.2 ± 0.02 | 10.8 ± 0.06 | 30.5→16.5 | 21.1→0.5→0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gongi, W.; Gomez Pinchetti, J.L.; Cordeiro, N.; Ouada, H.B. Extracellular Polymeric Substances Produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and Assessment of Their Antioxidant and Metal-Chelating Activities. Mar. Drugs 2022, 20, 227. https://doi.org/10.3390/md20040227
Gongi W, Gomez Pinchetti JL, Cordeiro N, Ouada HB. Extracellular Polymeric Substances Produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and Assessment of Their Antioxidant and Metal-Chelating Activities. Marine Drugs. 2022; 20(4):227. https://doi.org/10.3390/md20040227
Chicago/Turabian StyleGongi, Wejdene, Juan Luis Gomez Pinchetti, Nereida Cordeiro, and Hatem Ben Ouada. 2022. "Extracellular Polymeric Substances Produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and Assessment of Their Antioxidant and Metal-Chelating Activities" Marine Drugs 20, no. 4: 227. https://doi.org/10.3390/md20040227
APA StyleGongi, W., Gomez Pinchetti, J. L., Cordeiro, N., & Ouada, H. B. (2022). Extracellular Polymeric Substances Produced by the Thermophilic Cyanobacterium Gloeocapsa gelatinosa: Characterization and Assessment of Their Antioxidant and Metal-Chelating Activities. Marine Drugs, 20(4), 227. https://doi.org/10.3390/md20040227