
In Vitro and In Vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Ethics
2.3. DC Treatment
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Mixed Lymphatic Reaction (MLR)
2.6. Animal Model
2.7. Western Blot
2.8. Flow Cytometry
2.9. Statistical Analysis
3. Results and Discussions
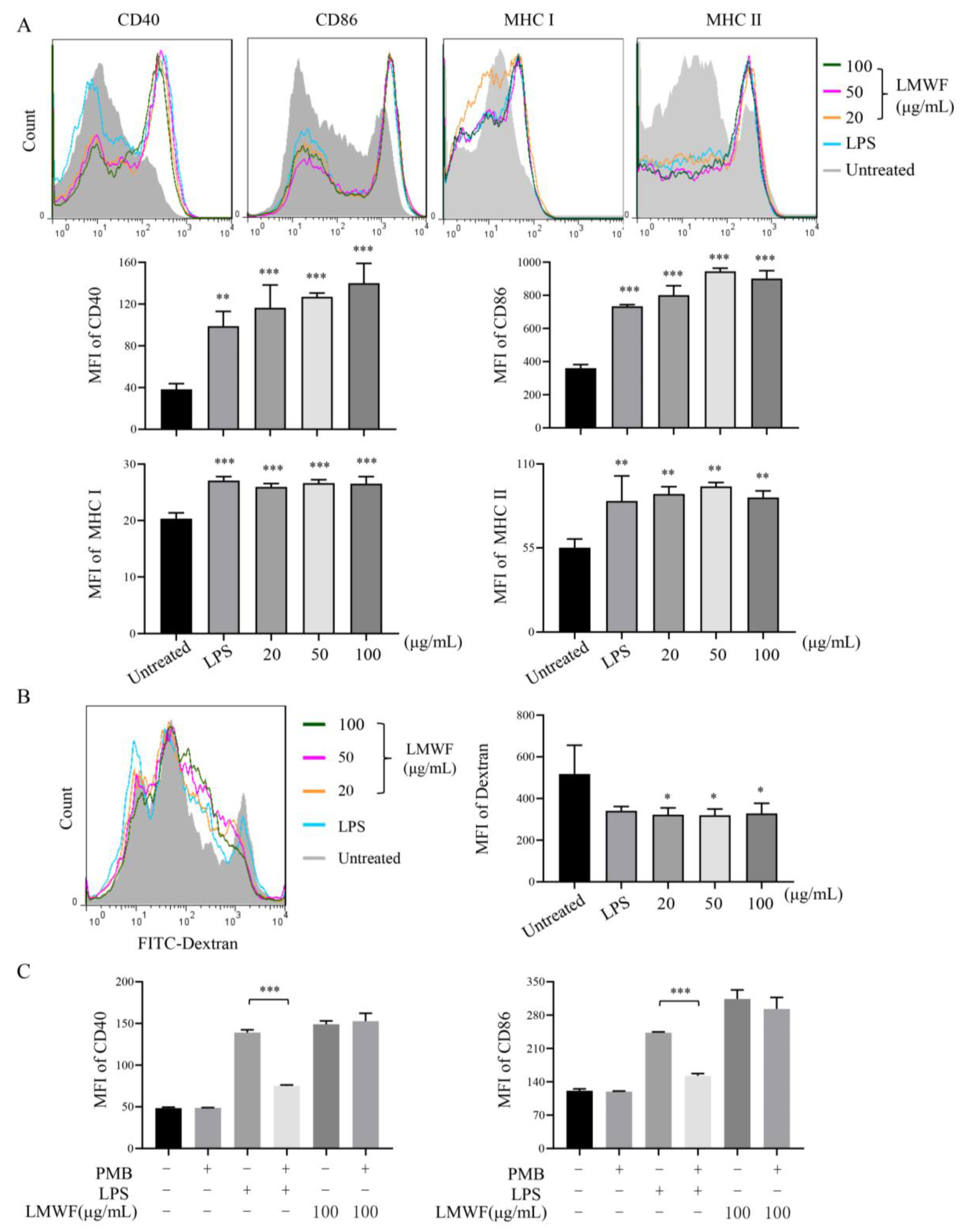
3.1. LMWF Can Stimulate the Maturation of DCs In Vitro and In Vivo
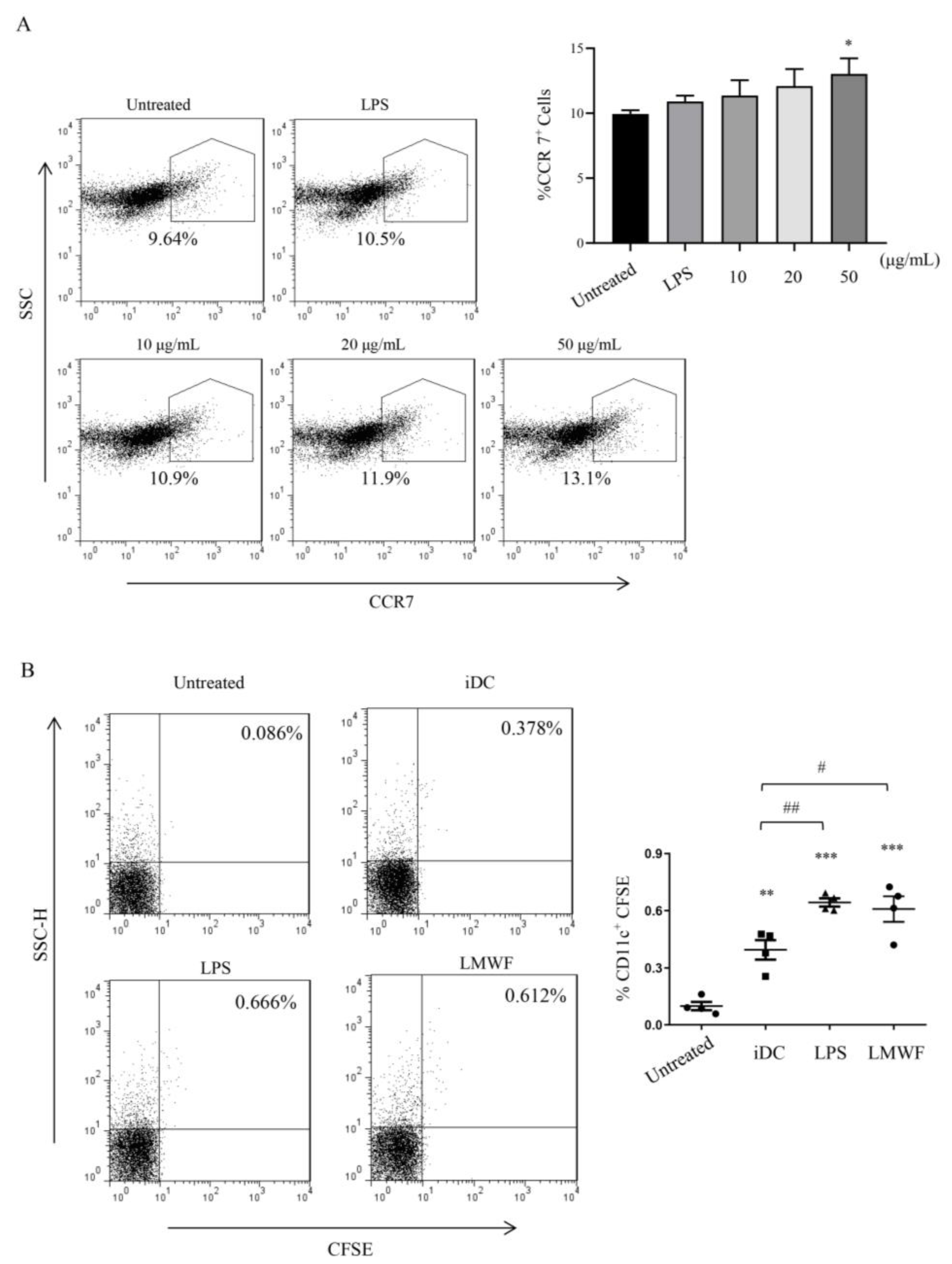
3.2. LMWF Enhances the Migration of DCs In Vitro and In Vivo
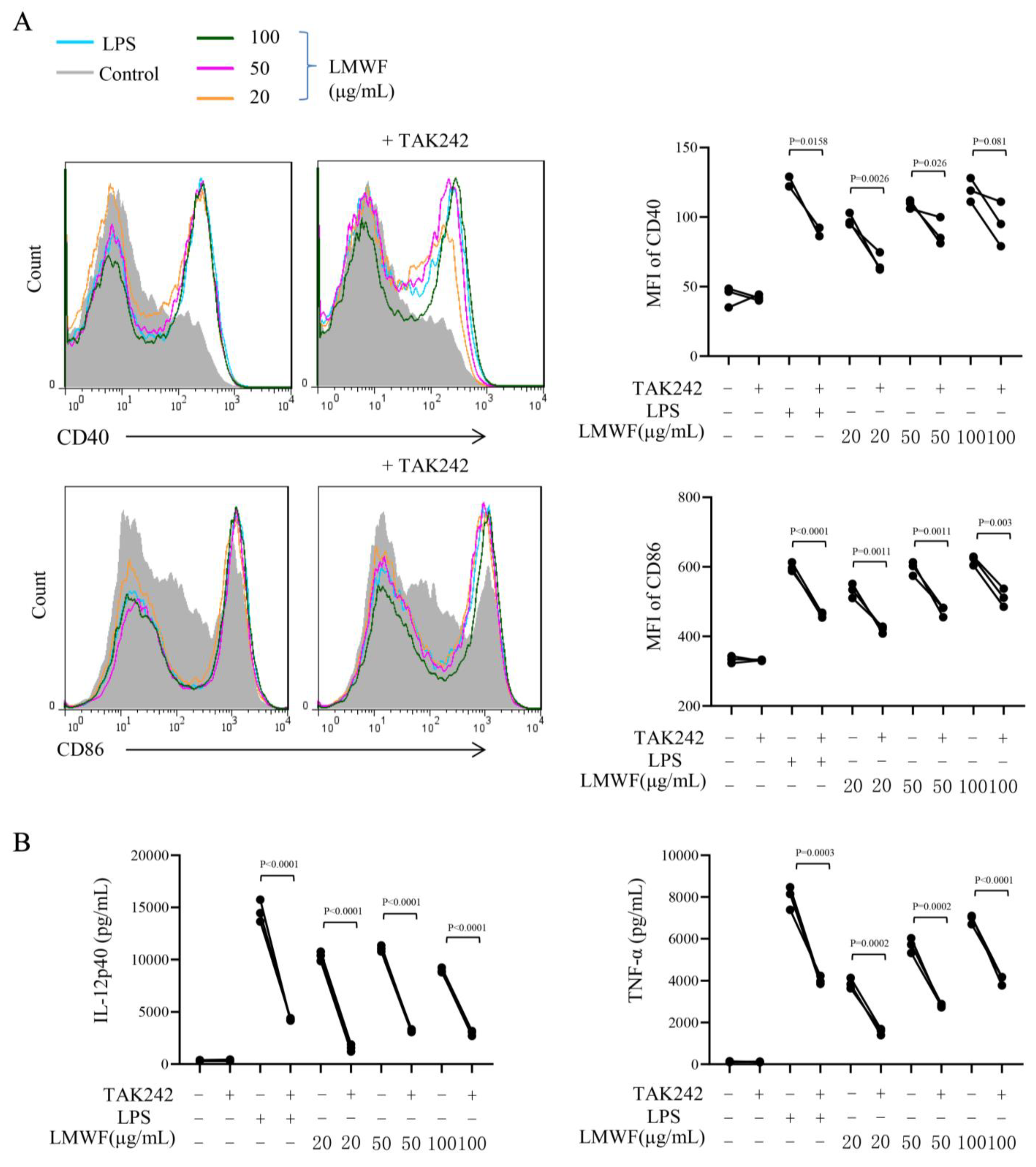
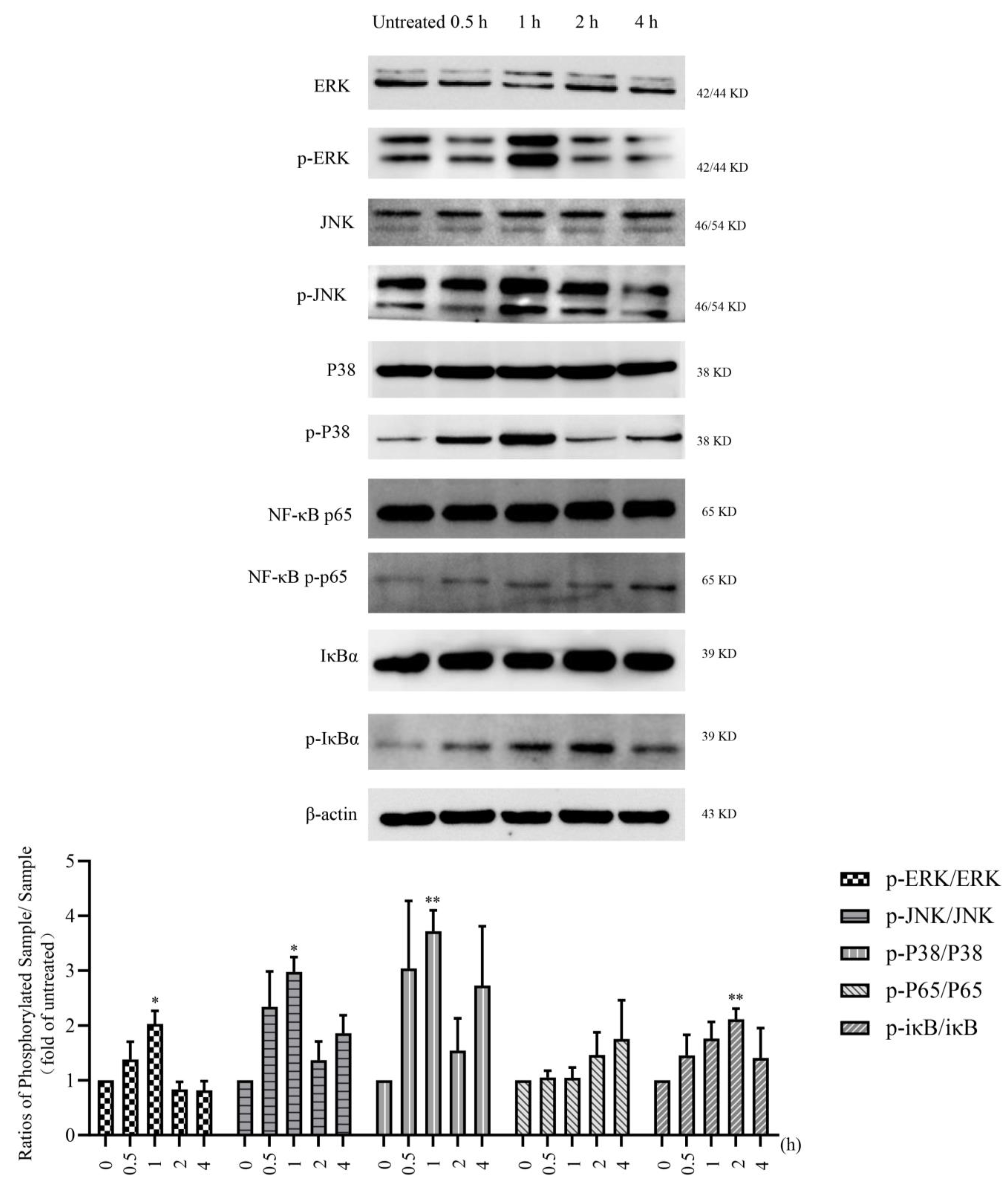
3.3. LMWF Promotes DC Maturation via TLR4 Signaling Pathway
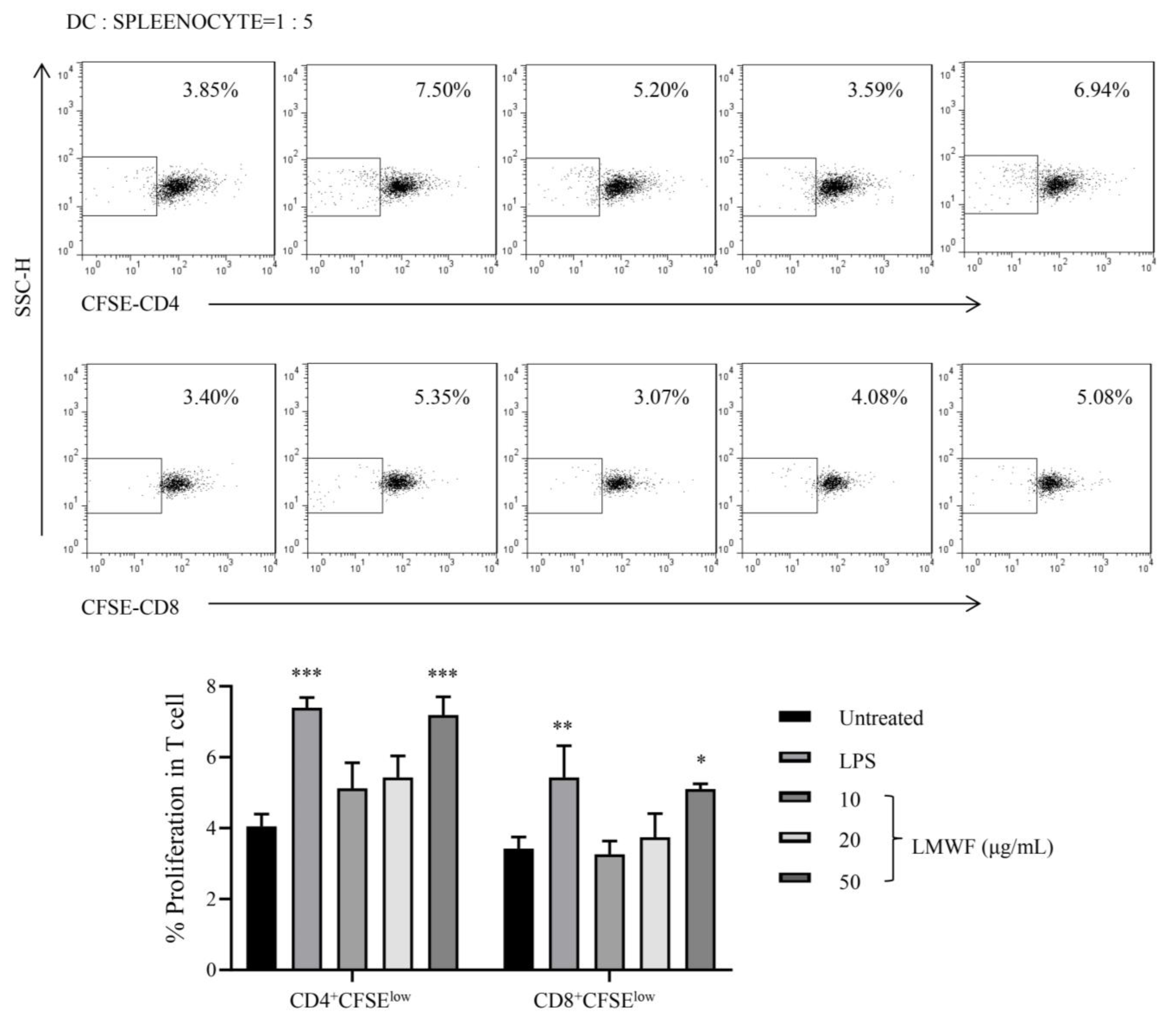
3.4. LMWF Shows Capability to Induce the Proliferation of Allogeneic T Cells
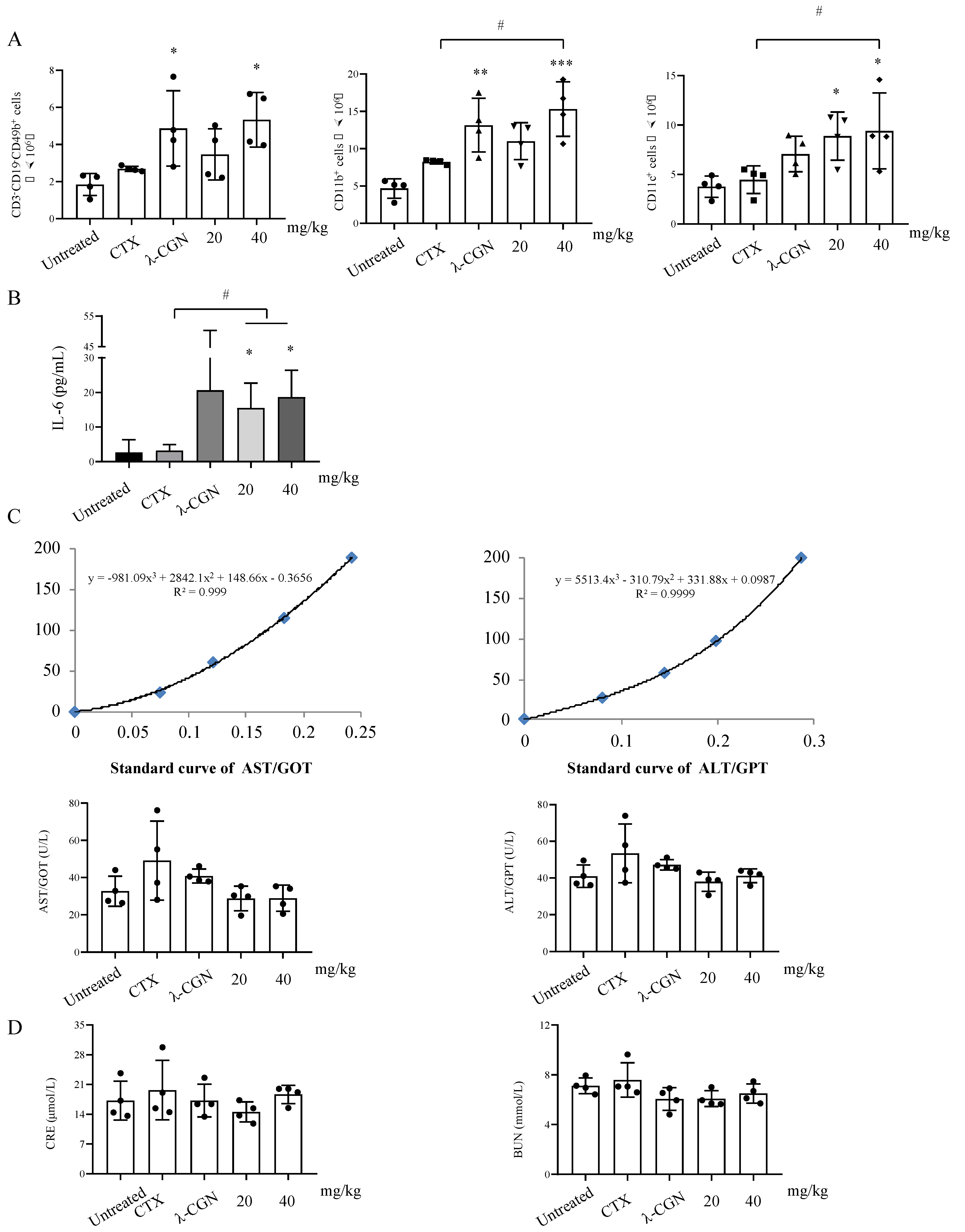
3.5. Immune Enhancement Effect of LMWF on Murine Immunity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vaamonde-García, C.; Flórez-Fernández, N.; Torres, M.D.; Lamas-Vázquez, M.J.; Blanco, F.J.; Domínguez, H.; Meijide-Faílde, R. Study of fucoidans as natural biomolecules for therapeutical applications in osteoarthritis. Carbohydr. Polym. 2021, 258, 117692. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhang, X.; Miao, Y.; Zhou, Y.; Shi, J.; Yan, M.; Chen, A. Studies on Antiviral and Immuno-Regulation Activity of Low Molecular Weight Fucoidan from Laminaria japonica. J. Ocean Univ. China 2018, 17, 705–711. [Google Scholar] [CrossRef]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structure characterization and antioxidant activity of fucoidan isolated from Undaria pinnatifida grown in New Zealand. Carbohydr. Polym. 2019, 212, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Li, H.; Liang, J.; Xu, Y.; Zhu, L.; Ye, X.; Wu, J.; Li, W.; Xiong, Q.; Li, S. Sulfated polysaccharide from Undaria pinnatifida stabilizes the atherosclerotic plaque via enhancing the dominance of the stabilizing components. Int. J. Biol. Macromol. 2019, 140, 621–630. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, T.; Li, J.; Xin, M.; Zhao, X. Structural characterization and effect on leukopenia of fucoidan from Durvillaea antarctica. Carbohydr. Polym. 2021, 256, 117529. [Google Scholar] [CrossRef]
- Mak, W.; Wang, S.K.; Liu, T.; Hamid, N.; Li, Y.; Lu, J.; White, W.L. Anti-Proliferation Potential and Content of Fucoidan Extracted from Sporophyll of New Zealand Undaria pinnatifida. Front. Nutr. 2014, 1, 9. [Google Scholar] [CrossRef]
- Oliveira, C.; Neves, N.M.; Reis, R.L.; Martins, A.; Silva, T.H. A review on fucoidan antitumor strategies: From a biological active agent to a structural component of fucoidan-based systems. Carbohydr. Polym. 2020, 239, 116131. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan Extracted from Undaria pinnatifida: Source for Nutraceuticals/Functional Foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef]
- Fletcher, H.; Biller, P.; Ross, A.; Adams, J. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef]
- Chen, Q.; Kou, L.; Wang, F.; Wang, Y. Size-dependent whitening activity of enzyme-degraded fucoidan from Laminaria japonica. Carbohydr. Polym. 2019, 225, 115211. [Google Scholar] [CrossRef]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from Macrocystis pyrifera Has Powerful Immune-Modulatory Effects Compared to Three Other Fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ma, C.; Sun, J.; Shao, Q.; Gao, W.; Zhang, Y.; Li, Z.; Xie, Q.; Dong, Z.; Qu, X. Fucoidan stimulation induces a functional maturation of human monocyte-derived dendritic cells. Int. Immunopharmacol. 2008, 8, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.-Y.; Jiang, Z.; Okimura, T.; Oda, T.; Yu, Q.; Jin, J.-O. Ascophyllan Purified from Ascophyllum nodosum Induces Th1 and Tc1 Immune Responses by Promoting Dendritic Cell Maturation. Mar. Drugs 2014, 12, 4148–4164. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, P.; Wang, X.; Aipire, A.; Li, M.; Yang, J.; Tao, H.; Ying, T.; Fu, C.; Wei, X.; et al. Purification, characterization and bioactivities of polysaccharides from Pleurotus ferulae. Food Funct. 2017, 8, 1905–1914. [Google Scholar] [CrossRef]
- Zhang, S.-N.; Choi, I.-K.; Huang, J.-H.; Yoo, J.-Y.; Choi, K.-J.; Yun, C.-O. Optimizing DC Vaccination by Combination with Oncolytic Adenovirus Coexpressing IL-12 and GM-CSF. Mol. Ther. 2011, 19, 1558–1568. [Google Scholar] [CrossRef]
- Yuan, P.; Aipire, A.; Yang, Y.; Wei, X.; Fu, C.; Zhou, F.; Mahabati, M.; Li, J.; Li, J. Comparison of the structural characteristics and immunostimulatory activities of polysaccharides from wild and cultivated Pleurotus feruleus. J. Funct. Foods 2020, 72, 104050. [Google Scholar] [CrossRef]
- Bi, D.; Yu, B.; Han, Q.; Lu, J.; White, W.L.; Lai, Q.; Cai, N.; Luo, W.; Gu, L.; Li, S.; et al. Immune Activation of RAW264.7 Macrophages by Low Molecular Weight Fucoidan Extracted from New Zealand Undaria pinnatifida. J. Agric. Food Chem. 2018, 66, 10721–10728. [Google Scholar] [CrossRef]
- Li, J.; Valentin, A.; Beach, R.K.; Alicea, C.; Felber, B.K.; Pavlakis, G.N. DNA is an efficient booster of dendritic cell-based vaccine. Hum. Vaccines Immunother. 2015, 11, 1927–1935. [Google Scholar] [CrossRef][Green Version]
- Schromm, A.B.; Paulowski, L.; Kaconis, Y.; Kopp, F.; Gutsmann, T. Cathelicidin and PMB neutralize endotoxins by multifactorial mechanisms including LPS interaction and targeting of host cell membranes. Proc. Natl. Acad. Sci. USA 2021, 118, 2101721118. [Google Scholar] [CrossRef]
- Aipire, A.; Mahabati, M.; Cai, S.; Wei, X.; Yuan, P.; Aimaier, A.; Wang, X.; Li, J. The immunostimulatory activity of polysaccharides from Glycyrrhiza uralensis. PeerJ 2020, 8, e8294. [Google Scholar] [CrossRef]
- Gumusay, O.; Renslo, J.R.; Wabl, C.A.; Chien, A.J.; Melisko, M.E.; Majure, M.C.; Rahmaputri, S.D.; Moasser, M.; Park, J.; Rugo, H.S. A phase Ib/II study of eribulin with cyclophosphamide (EC) in patients (pts) with advanced breast cancer (ABC). J. Clin. Oncol. 2020, 38, e13079. [Google Scholar] [CrossRef]
- McCurdy, S.R.; Luznik, L. How we perform haploidentical stem cell transplantation with posttransplant cyclophosphamide. Blood 2019, 134, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Yi, R.; Li, G.; Sun, P.; Qian, Y.; Zhao, X. Immunomodulatory Effect of Tremella Polysaccharides against Cyclophosphamide-Induced Immunosuppression in Mice. Molecules 2018, 23, 239. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Aipire, A.; Luo, J.; Yuan, P.; Zhang, F. The combination of Pleurotus ferulae water extract and CpG-ODN enhances the immune responses and antitumor efficacy of HPV peptides pulsed dendritic cell-based vaccine. Vaccine 2016, 34, 3568–3575. [Google Scholar] [CrossRef] [PubMed]
- Worbs, T.; Hammerschmidt, S.I.; Förster, R. Dendritic cell migration in health and disease. Nat. Rev. Immunol. 2016, 17, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Chun, K.R.; Kim, J.K.; Suk, K.; Jung, Y.M.; Lee, W.H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef]
- Xie, S.-Z.; Hao, R.; Zha, X.-Q.; Pan, L.-H.; Liu, J.; Luo, J.-P. Polysaccharide of Dendrobium huoshanense activates macrophages via toll-like receptor 4-mediated signaling pathways. Carbohydr. Polym. 2016, 146, 292–300. [Google Scholar] [CrossRef]
- Park, M.J.; Ryu, H.S.; Kim, J.S.; Lee, H.K.; Kang, J.S.; Yun, J.; Kim, S.Y.; Lee, M.K.; Hong, J.T.; Kim, Y.; et al. Platycodon grandiflorum polysaccharide induces dendritic cell maturation via TLR4 signaling. Food Chem. Toxicol. 2014, 72, 212–220. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Wang, W.; Luo, J.; Aipire, A.; Li, J.; Zhang, F. Pleurotus ferulae water extract enhances the maturation and function of murine bone marrow-derived dendritic cells through TLR4 signaling pathway. Vaccine 2015, 33, 1923–1933. [Google Scholar] [CrossRef]
- Makarenkova, I.; Logunov, D.Y.; Tukhvatulin, A.; Semenova, I.; Besednova, N.; Zvyagintseva, T. Interactions between sulfated polysaccharides from sea brown algae and Toll-like receptors on HEK293 eukaryotic cells in vitro. Bull. Exp. Biol. Med. 2021, 154, 241–244. [Google Scholar] [CrossRef]
- Jin, J.-O.; Zhang, W.; Du, J.-Y.; Wong, K.-W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.-C.; Aboelella, N.S.; Bryan, L.; Shi, H.; Zhou, G. The Monocytes That Repopulate in Mice After Cyclophosphamide Treatment Acquire a Neutrophil Precursor Gene Signature and Immunosuppressive Activity. Front. Immunol. 2021, 11, 3576. [Google Scholar] [CrossRef] [PubMed]
- Ahlmann, M.; Hempel, G. The effect of cyclophosphamide on the immune system: Implications for clinical cancer therapy. Cancer Chemother. Pharmacol. 2016, 78, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.-Y.; Moon, S.-Y.; Joo, H.-G. Differential effects of fucoidans with low and high molecular weight on the viability and function of spleen cells. Food Chem. Toxicol. 2014, 68, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Techel, I.; Lahrsen, E.; Alban, S. Degraded fucoidan fractions and beta-1,3-glucan sulfates inhibit CXCL12-induced Erk1/2 activation and chemotaxis in Burkitt lymphoma cells. Int. J. Biol. Macromol. 2019, 143, 968–976. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
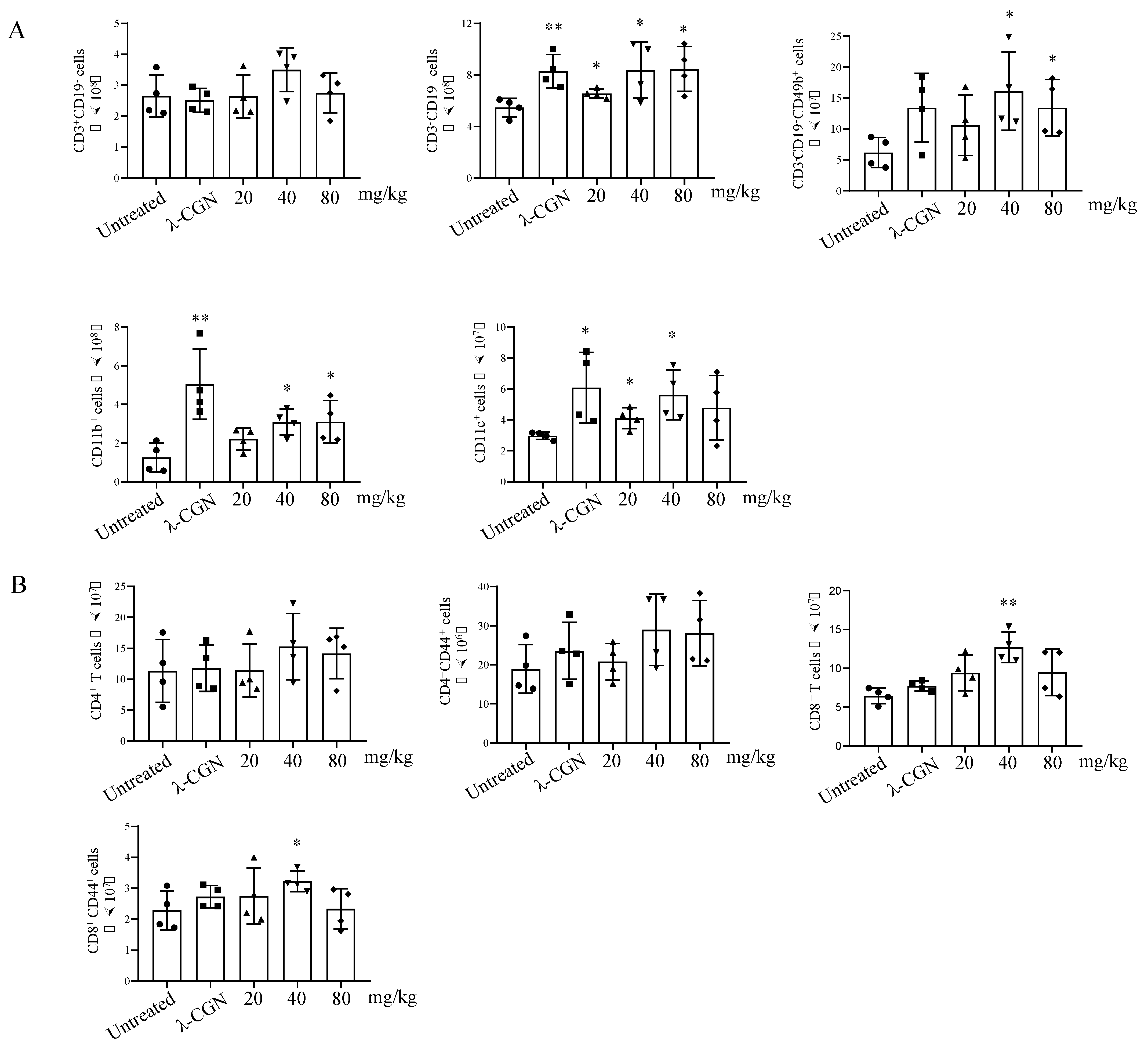{kind=link}
{kind=link}
| Untreated | λ-CGN | 20 mg/kg | 40 mg/kg | 80 mg/kg | |
|---|---|---|---|---|---|
| Heart | 5.03 ± 0.44 | 5.09 ± 0.16 | 5.08 ± 0.79 | 5.94 ± 0.37 | 5.64 ± 0.66 |
| Liver | 63.80 ± 3.64 | 72.23 ± 4.32 * | 65.99 ± 5.01 | 71.19 ± 1.52 | 70.11 ± 7.24 |
| Spleen | 3.82 ± 0.84 | 8.76 ± 1.29 *** | 6.32 ± 1.37 * | 6.29 ± 0.63 * | 6.81 ± 0.67 ** |
| Lung | 8.42 ± 0.67 | 7.00 ± 0.23 | 8.42 ± 1.17 | 8.17 ± 0.40 | 8.07 ± 0.90 |
| Kidney | 13.31 ± 0.64 | 13.28 ± 1.02 | 13.50 ± 0.86 | 14.50 ± 0.84 | 14.24 ± 0.85 |
| Thymus | 3.10 ± 0.53 | 3.29 ± 0.76 | 3.36 ± 0.69 | 5.02 ± 0.73 * | 4.28 ± 1.29 |
| Body Weight (g) | Spleen Index (mg/g) | Thymus Index (mg/g) | |
|---|---|---|---|
| Control | 25.73 ± 1.24 | 4.36 ± 0.32 | 4.49 ± 0.87 |
| CTX | 18.57 ± 4.99 * | 11.04 ± 0.46 ** | 3.88 ± 1.52 |
| λ-CGN | 20.83 ± 2.77 | 13.13 ± 2.60 *** | 4.44 ± 1.28 |
| 20 mg/kg | 22.87 ± 0.88 | 16.85 ± 2.59 ***/# | 3.62 ± 0.29 |
| 40 mg/kg | 20.66 ± 2.13 | 19.65 ± 2.34 ***/### | 2.95 ± 1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Yang, X.; Yuan, P.; Cai, S.; Bao, J.; Zhao, Y.; Aimaier, A.; Aipire, A.; Lu, J.; Li, J. In Vitro and In Vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida. Mar. Drugs 2022, 20, 197. https://doi.org/10.3390/md20030197
Liu L, Yang X, Yuan P, Cai S, Bao J, Zhao Y, Aimaier A, Aipire A, Lu J, Li J. In Vitro and In Vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida. Marine Drugs. 2022; 20(3):197. https://doi.org/10.3390/md20030197
Chicago/Turabian StyleLiu, Litong, Xu Yang, Pengfei Yuan, Shanshan Cai, Jing Bao, Yanan Zhao, Alimu Aimaier, Adila Aipire, Jun Lu, and Jinyao Li. 2022. "In Vitro and In Vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida" Marine Drugs 20, no. 3: 197. https://doi.org/10.3390/md20030197
APA StyleLiu, L., Yang, X., Yuan, P., Cai, S., Bao, J., Zhao, Y., Aimaier, A., Aipire, A., Lu, J., & Li, J. (2022). In Vitro and In Vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida. Marine Drugs, 20(3), 197. https://doi.org/10.3390/md20030197