
Effect of Different Initial Fermentation pH on Exopolysaccharides Produced by Pseudoalteromonas agarivorans Hao 2018 and Identification of Key Genes Involved in Exopolysaccharide Synthesis via Transcriptome Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
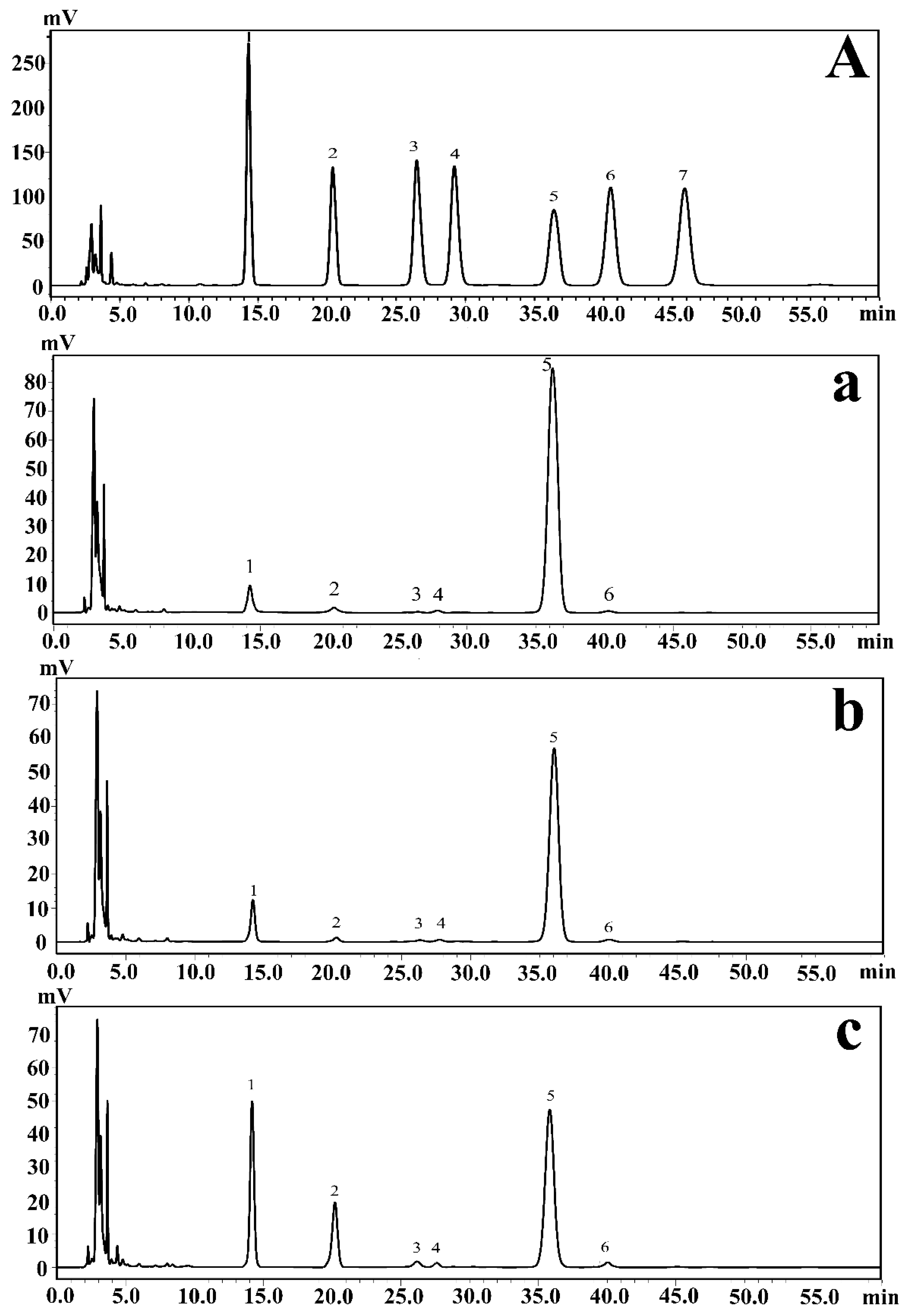
2.1. Analysis of the Monosaccharide Composition of EPSs via HPLC
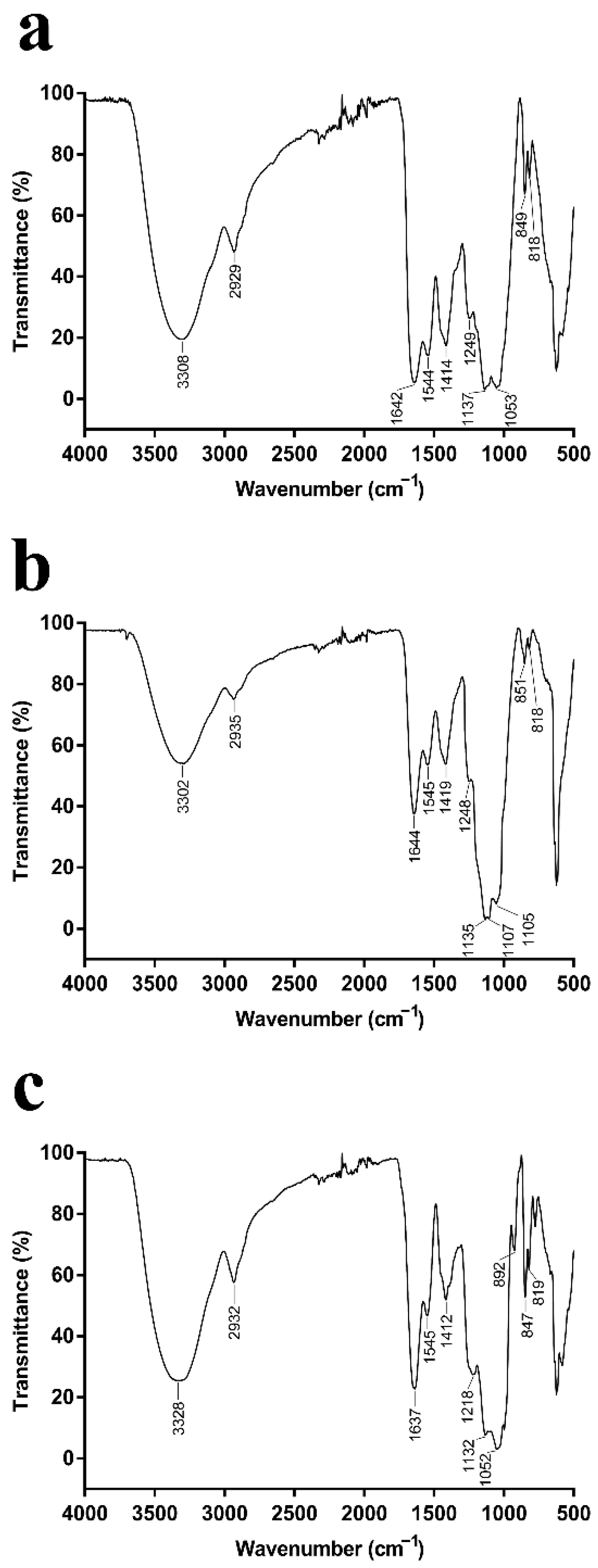
2.2. Analysis of the EPSs via FT-IR
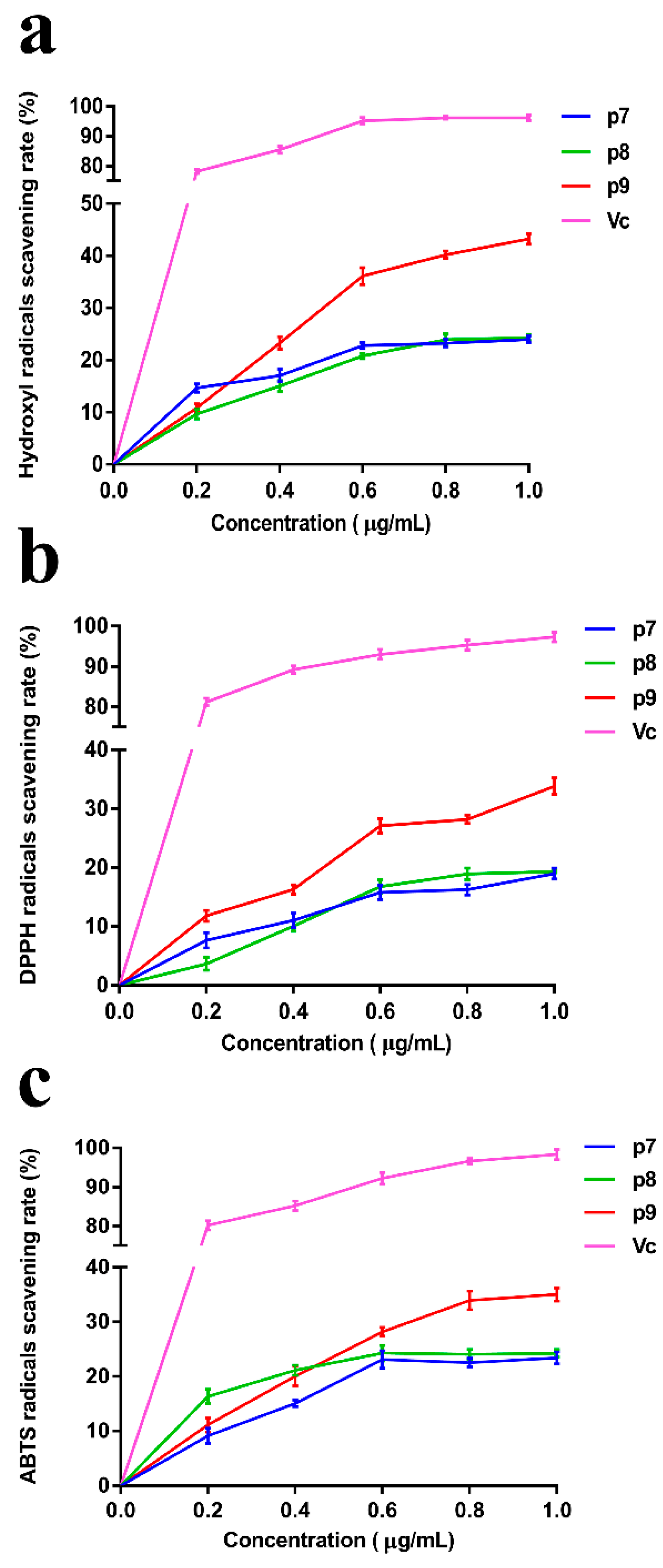
2.3. Analysis of the Antioxidant Activity of EPSs
2.4. Sequencing of the Pseudoalteromonas agarivorans Hao 2018 Transcriptome
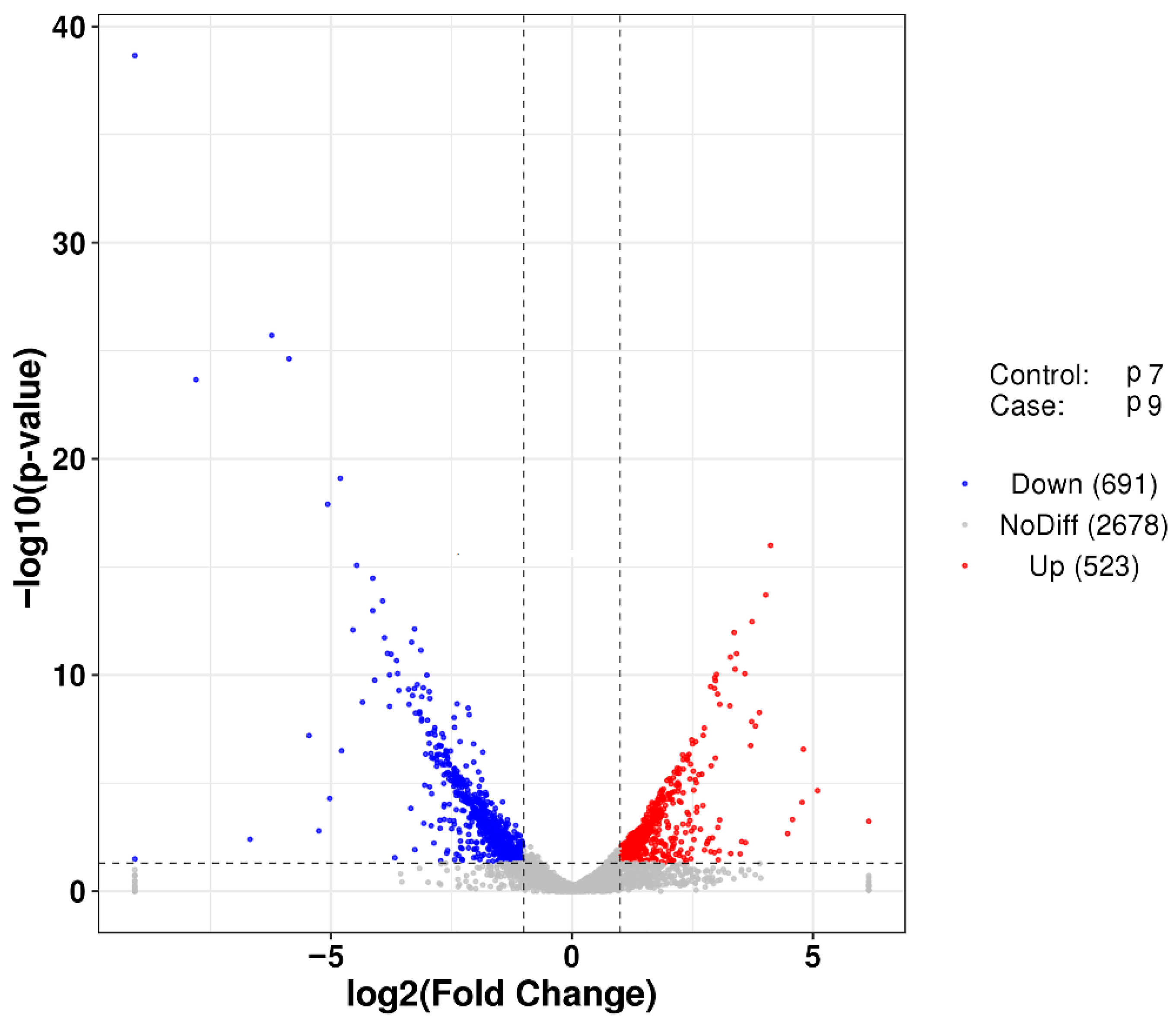
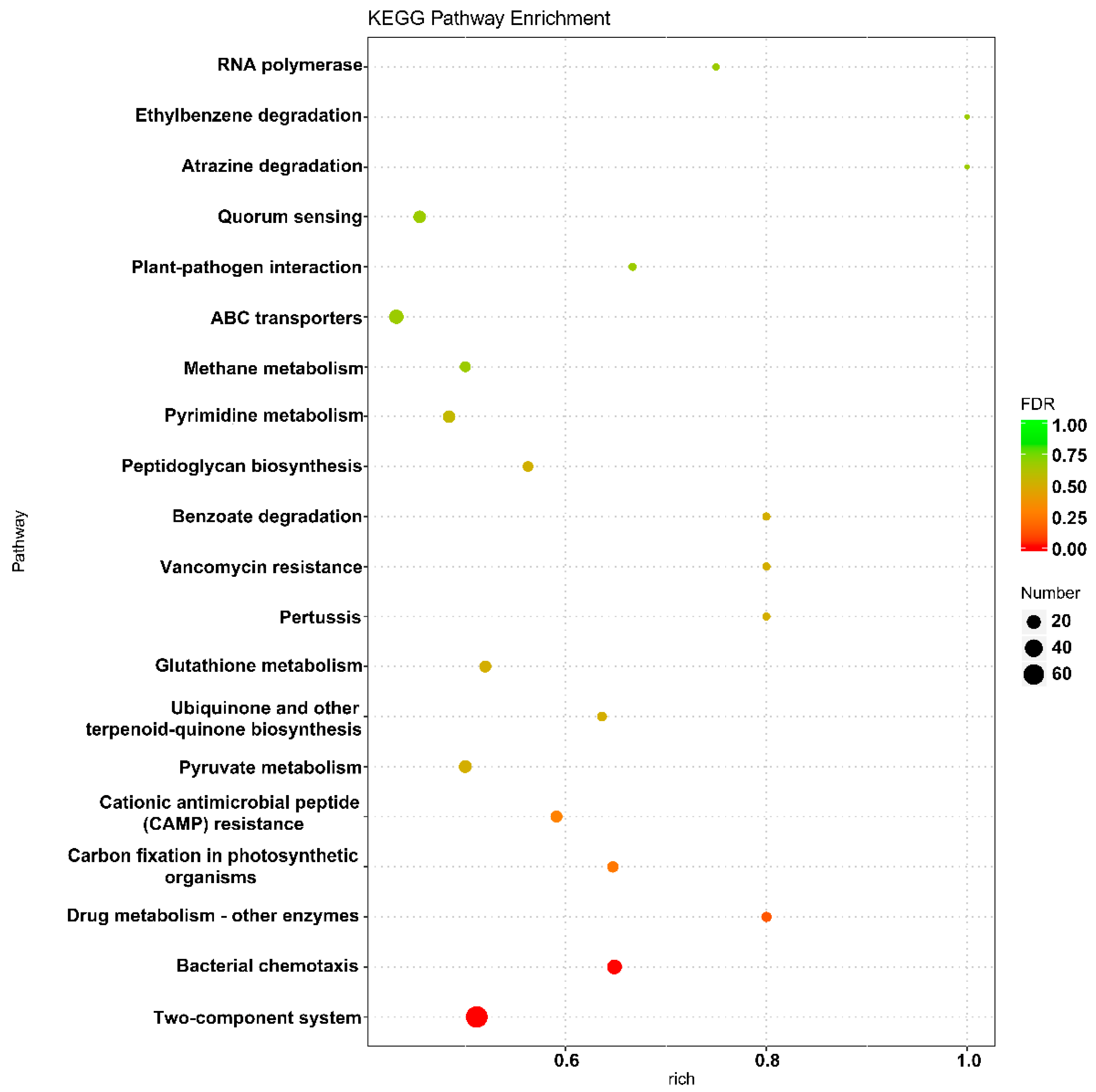
2.5. GO and KEGG Analysis to Identify the Metabolic Processes Involving Genes with Significant Differences
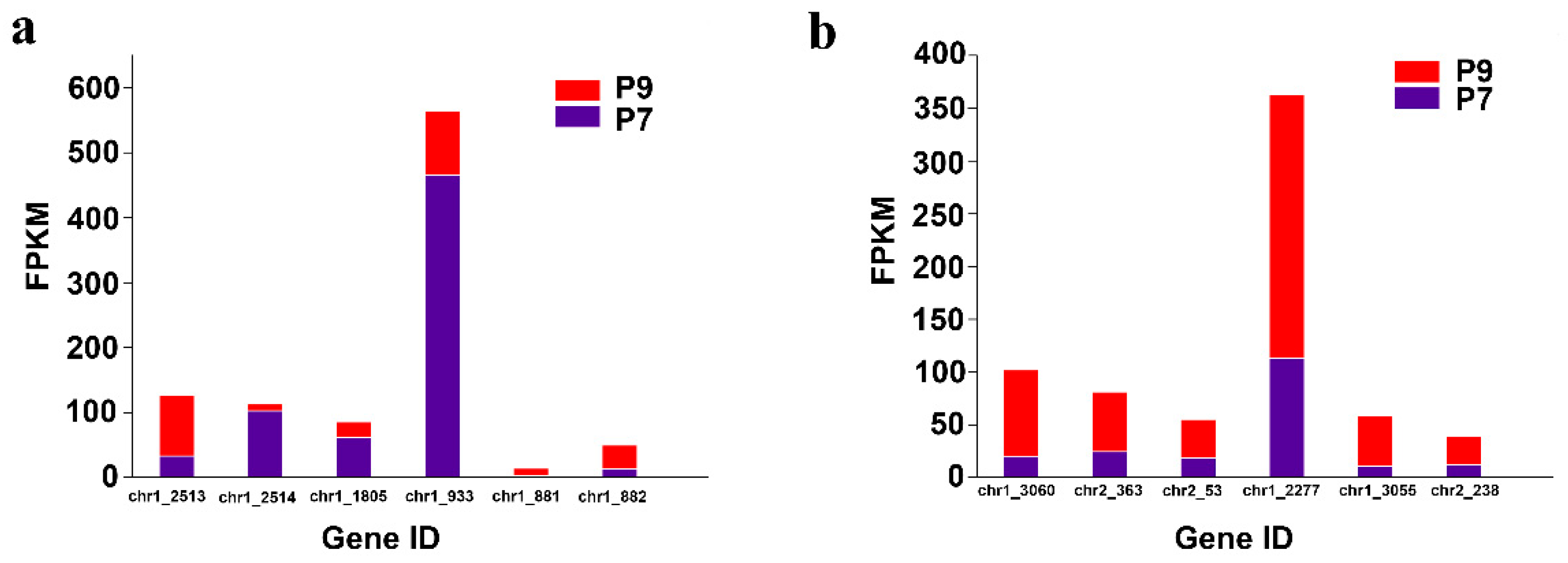
2.6. Analysis of the Expression of Genes Related to the Two-Component Regulatory System
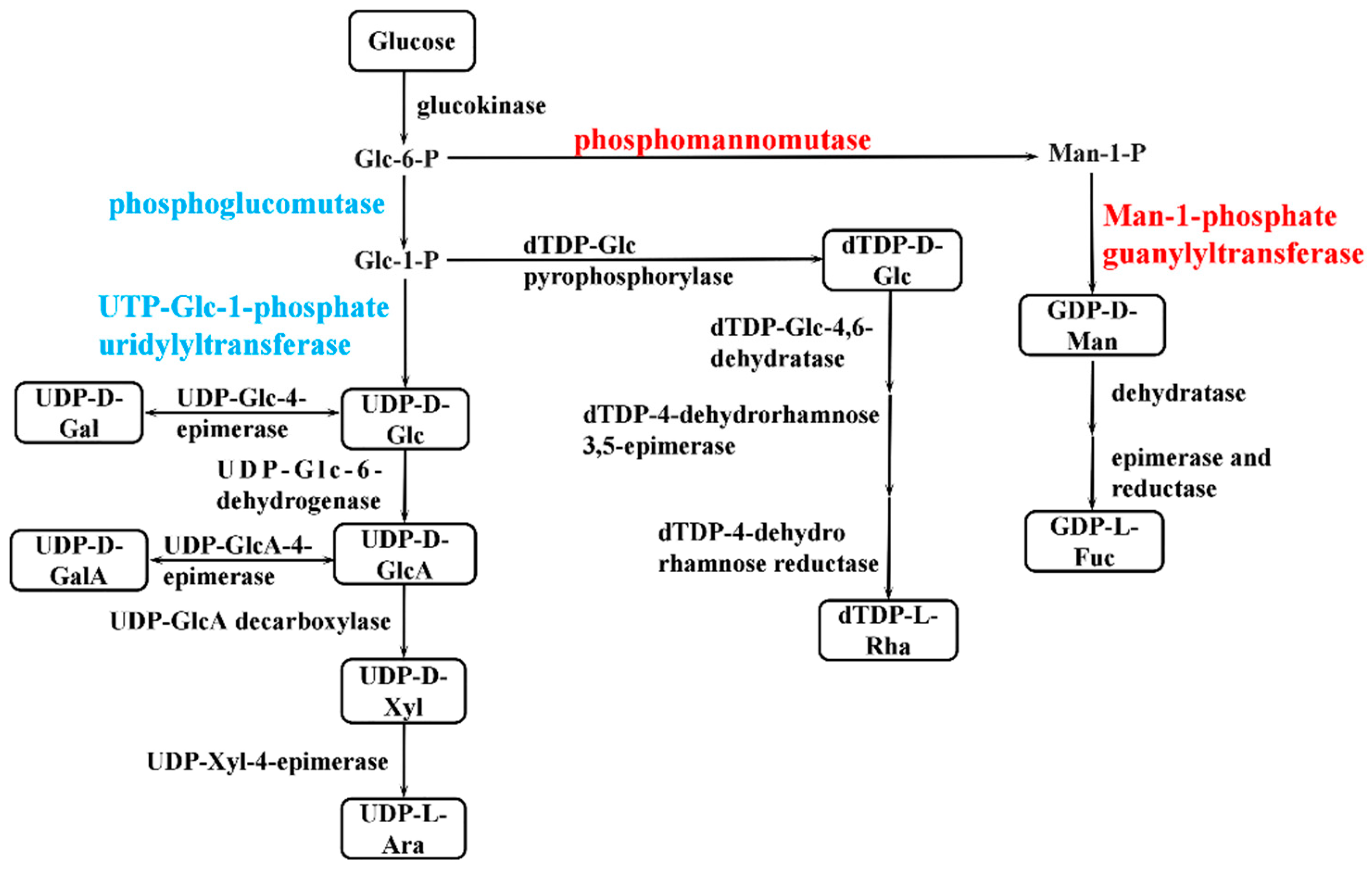
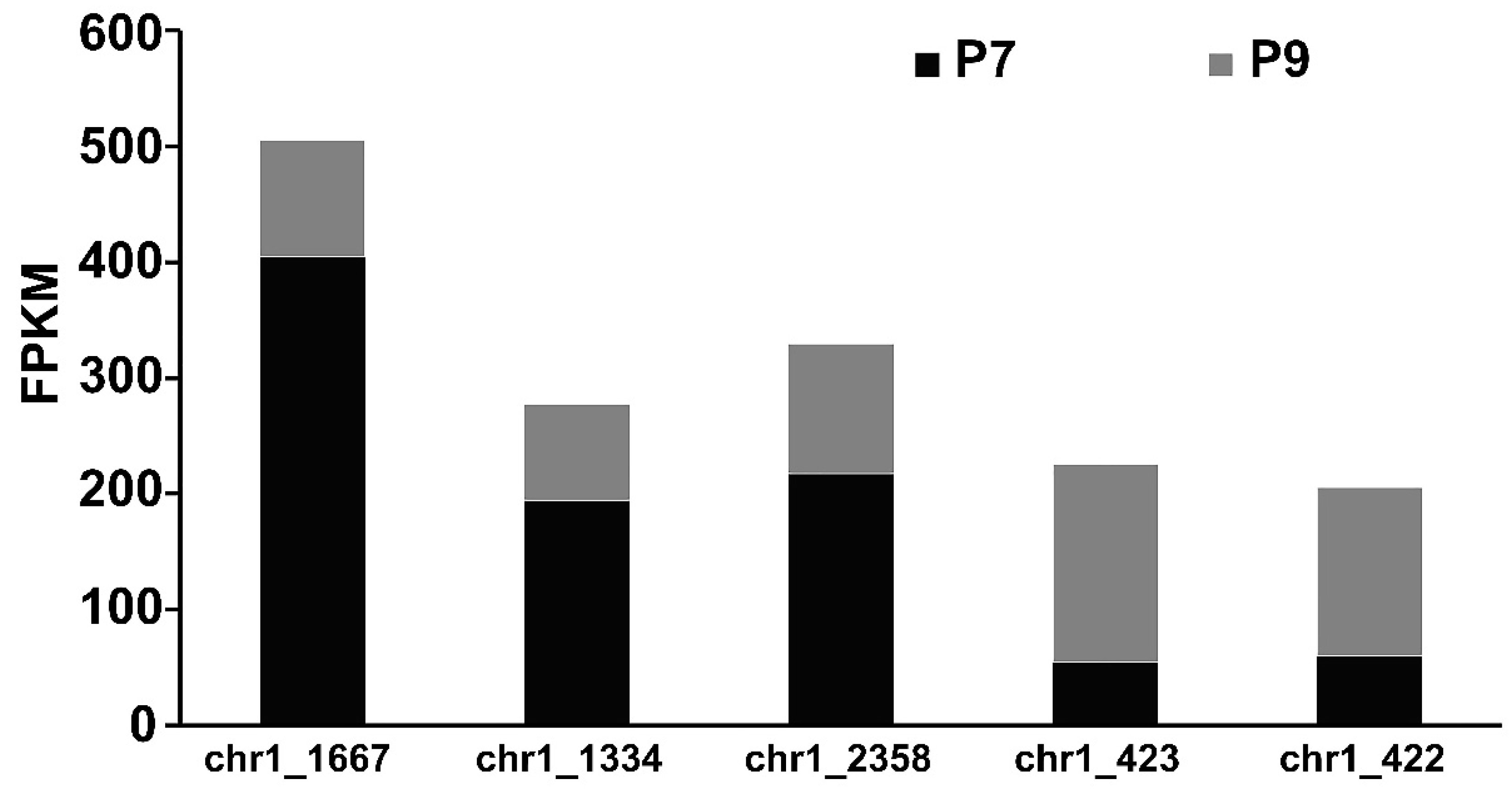
2.7. Analysis of the Expression of Genes Directly Related to EPS Synthesis
3. Discussion
4. Materials and Methods
4.1. Extraction and Purification of EPSs under Different Fermentation Conditions
4.2. Determination of EPSs via HPLC and FT-IR
4.3. Determination of Antioxidant Activity of EPSs to Scavenge OH, DPPH, and ABTS Free Radicals
4.4. Transcriptome Sequencing
4.5. Transcriptome Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harris, R.H.; Mitchell, R. The role of polymers in microbial aggregation. Annu. Rev. Microbiol. 1973, 27, 27–50. [Google Scholar] [CrossRef]
- Biddanda, B. Microbial synthesis of macroparticulate matter. Mar. Ecol. Prog. Ser. 1985, 20, 241–251. [Google Scholar] [CrossRef]
- Alldredge, A.L.; Silver, M.W. Characteristics, dynamics and significance of marine snow. Prog. Oceanogr. 1988, 20, 41–82. [Google Scholar] [CrossRef]
- Paerl, H.W. Microbial attachment to particles in marine and freshwater ecosystems. Microb. Ecol. 1975, 2, 73–83. [Google Scholar] [CrossRef]
- Marshall, K.C. Mechanisms of bacterial adhesion at solid-water interfaces. In Bacterial Adhesion; Savage, D.C., Fletcher, M., Eds.; Springer: Boston, MA, USA, 1985; pp. 133–161. ISBN 978-1-4615-6516-1. [Google Scholar]
- Vincent, P.; Pignet, P.; Talmont, F.; Bozzi, L.; Fournet, B.; Guezennec, J.; Jeanthon, C.; Prieur, D. Production and characterization of an exopolysaccharide excreted by a deep-Sea hydrothermal vent bacterium isolated from the polychaete annelid Alvinella pompejana. Appl. Environ. Microbiol. 1994, 60, 4134–4141. [Google Scholar] [CrossRef] [PubMed]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Sutherland, I.W. Biofilm Exopolysaccharides. In Microbial Extracellular Polymeric Substances; Wingender, J., Neu, T.R., Flemming, H.-C., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 73–92. ISBN 978-3-642-64277-7. [Google Scholar]
- Sutherland, I.W. Biofilm Exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef]
- Decho, A.W.; Herndl, G.J. Microbial activities and the transformation of organic matter within mucilaginous material. Sci. Total Environ. 1995, 165, 33–42. [Google Scholar] [CrossRef]
- Bitton, G.; Freihofer, V. Influence of extracellular polysaccharides on the toxicity of copper and cadmium Toward Klebsiella aerogenes. Microb. Ecol. 1977, 4, 119–125. [Google Scholar] [CrossRef]
- Decho, A.W.; Lopez, G.R. Exopolymer microenvironments of microbial flora: Multiple and interactive effects on trophic relationships. Limnol. Oceanogr. 1993, 38, 1633–1645. [Google Scholar] [CrossRef]
- Dade, W.B.; Davis, J.D.; Nichols, P.D.; Nowell, A.R.M.; Thistle, D.; Trexler, M.B.; White, D.C. Effects of bacterial exopolymer adhesion on the entrainment of sand. Geomicrobiol. J. 1990, 8, 1–16. [Google Scholar] [CrossRef]
- Uhlinger, D.J.; White, D.C. Relationship between physiological status and formation of extracellular polysaccha ride glycocalyx in Pseudomonas atlantica. Appl. Environ. Microbiol. 1983, 45, 64–70. [Google Scholar] [CrossRef]
- Llamas, I.; Mata, J.A.; Tallon, R.; Bressollier, P.; Urdaci, M.C.; Quesada, E.; Béjar, V. Characterization of the exopolysaccharide produced by Salipiger mucosus A3T, a Halophilic Species Belonging to the Alphaproteobacteria, Isolated on the Spanish Mediterranean Seaboard. Mar. Drugs 2010, 8, 2240–2251. [Google Scholar] [CrossRef]
- Wang, C.; Fan, Q.; Zhang, X.; Lu, X.; Xu, Y.; Zhu, W.; Zhang, J.; Hao, W.; Hao, L. Isolation, characterization, and pharmaceutical applications of an exopolysaccharide from Aerococcus uriaeequi. Mar. Drugs 2018, 16, 337. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Liu, W.; Liu, K.; Shan, K.; Wang, C.; Xi, C.; Liu, J.; Fan, Q.; Zhang, X.; Lu, X.; et al. Isolation, optimization of fermentation conditions, and characterization of an exopolysaccharide from Pseudoalteromonas agarivorans Hao 2018. Mar. Drugs 2019, 17, 703. [Google Scholar] [CrossRef]
- Vinroot, S.; Torzilli, A.P. Interactive effects of inoculum density, agitation, and Ph on dimorphism in a salt marsh isolate of Aureobasidium pullulans. Mycologia 1988, 80, 376–381. [Google Scholar] [CrossRef]
- Esgalhado, M.E.; Roseiro, J.C.; Collaço, M.T.A. Interactive effects of pH and temperature on cell growth and polymer production by Xanthomonas campestris. Process Biochem. 1995, 30, 667–671. [Google Scholar] [CrossRef]
- Yang, F.-C.; Liau, C.-B. The influence of environmental conditions on polysaccharide formation by Ganoderma lucidum in submerged cultures. Process Biochem. 1998, 33, 547–553. [Google Scholar] [CrossRef]
- Catley, B.J. Role of pH and nitrogen limitation in the elaboration of the extracellular polysaccharide pullulan by Pullularia pullulans. Appl. Microbiol. 1971, 22, 650–654. [Google Scholar] [CrossRef]
- Shu, C.-H.; Lung, M.-Y. Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia camphorata in batch cultures. Process Biochem. 2004, 39, 931–937. [Google Scholar] [CrossRef]
- Fang, Q.-H.; Zhong, J.-J. Effect of Initial PH on Production of Ganoderic Acid and Polysaccharide by Submerged Fermentation of Ganoderma Lucidum. Process Biochem. 2002, 37, 769–774. [Google Scholar] [CrossRef]
- Hu, S.; Xiao, X.; Wu, X.; Xia, X.; Yu, Y.; Wu, H. Comparative transcriptomic analysis by RNA-Seq of acid tolerance response (ATR) in EHEC O157:H7. LWT-Food Sci. Technol. 2017, 79, 300–308. [Google Scholar] [CrossRef]
- Shan, K.; Wang, C.; Liu, W.; Liu, K.; Jia, B.; Hao, L. Genome sequence and transcriptomic profiles of a marine bacterium, Pseudoalteromonas agarivorans Hao 2018. Sci. Data 2019, 6, 10. [Google Scholar] [CrossRef]
- Blackwell, J. Infrared and Raman Spectroscopy of polysaccharides. In Extracellular Microbial Polysaccharides; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1977; Volume 45, pp. 103–113. ISBN 978-0-8412-0372-3. [Google Scholar]
- Deng, Y.-Y.; Yi, Y.; Zhang, L.-F.; Zhang, R.-F.; Zhang, Y.; Wei, Z.-C.; Tang, X.-J.; Zhang, M.-W. Immunomodulatory activity and partial characterisation of polysaccharides from Momordica charantia. Molecules 2014, 19, 13432–13447. [Google Scholar] [CrossRef]
- Huang, K.; Li, Y.; Tao, S.; Wei, G.; Huang, Y.; Chen, D.; Wu, C. Purification, characterization and biological activity of polysaccharides from Dendrobium officinale. Molecules 2016, 21, 701. [Google Scholar] [CrossRef] [PubMed]
- Whistler, R.L.; House, L.R. Infrared spectra of sugar anomers. Anal. Chem. 1953, 25, 1463–1466. [Google Scholar] [CrossRef]
- Yi, Y.; Liao, S.T.; Zhang, M.W.; Shi, J.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C. Physicochemical characteristics and immunomodulatory activities of three polysaccharide-protein complexes of Longan pulp. Molecules 2011, 16, 6148–6164. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Zhang, H.-Y.; Li, W.-J.; Ye, J.-Z. Chemical Constituents and Structural Characterization of Polysac charides from Four Typical Bamboo Species Leaves. Molecules 2015, 20, 4162–4179. [Google Scholar] [CrossRef]
- Kačuráková, M.; Wilson, R.H. Developments in mid-infrared FT-IR spectroscopy of selected carbohydrates. Carbohydr. Polym. 2001, 44, 291–303. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, L.-M. Chemical structural and chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Polym. 2009, 76, 349–361. [Google Scholar] [CrossRef]
- Marijuán, P.C.; Navarro, J.; del Moral, R. On Prokaryotic Intelligence: Strategies for Sensing the Environment. Biosystems 2010, 99, 94–103. [Google Scholar] [CrossRef]
- Appleby, J.L.; Parkinson, J.S.; Bourret, R.B. Signal transduction via the multi-Step phosphorelay: Not necessarily a road less traveled. Cell 1996, 86, 845–848. [Google Scholar] [CrossRef]
- Jin, Q.; Yuan, Z.; Xu, J.; Wang, Y.; Shen, Y.; Lu, W.; Wang, J.; Liu, H.; Yang, J.; Yang, F.; et al. Genome sequence of Shigella flexneri 2a: Insights into pathogenicity through comparison with genomes of Escherichia coli K12 and O157. Nucleic Acids Res. 2002, 30, 4432–4441. [Google Scholar] [CrossRef]
- Stephenson, K.; Hoch, J.A. Histidine kinase-mediated signal transduction systems of pathogenic microorganisms as targets for therapeutic intervention. Curr. Drug Targets-Infect. Disord. 2002, 2, 235–246. [Google Scholar] [CrossRef]
- Watanabe, T.; Hashimoto, Y.; Yamamoto, K.; Hirao, K.; Ishihama, A.; Hino, M.; Utsumi, R. Isolation and charac terization of inhibitors of the essential histidine kinase, YycG in Bacillus subtilis and Staphylococcus aureus. J. Antibiot. 2003, 56, 1045–1052. [Google Scholar] [CrossRef][Green Version]
- Bernardini, M.L.; Fontaine, A.; Sansonetti, P.J. The two-component regulatory system OmpR-EnvZ controls the virulence of Shigella flexneri. J. Bacteriol. 1990, 172, 6274–6281. [Google Scholar] [CrossRef]
- Yang, J.; Wang, J.; Yao, Z.-J.; Jin, Q.; Shen, Y.; Chen, R. GenomeComp: A visualization tool for microbial genome comparison. J. Microbiol. Methods 2003, 54, 423–426. [Google Scholar] [CrossRef]
- Kumar, S.; Tamura, K.; Jakobsen, I.B.; Nei, M. MEGA2: Molecular evolutionary genetics analysis software. Bioinformatics 2001, 17, 1244–1245. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-L.; Zhao, F.; Shi, M.; Zhang, X.-Y.; Zhou, B.-C.; Zhang, Y.-Z.; Chen, X.-L. Characterization and biotechno logical potential analysis of a new exopolysaccharide from the Arctic marine bacterium Polaribacter sp. SM1127. Sci. Rep. 2015, 5, 18435. [Google Scholar] [CrossRef] [PubMed]
- Lo, T.C.-T.; Chang, C.A.; Chiu, K.-H.; Tsay, P.-K.; Jen, J.-F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Meng, L.; Sun, S.; Li, R.; Shen, Z.; Wang, P.; Jiang, X. Antioxidant activity of polysaccharides produced by Hirsutella sp. and relation with their chemical characteristics. Carbohydr. Polym. 2015, 117, 452–457. [Google Scholar] [CrossRef]
- Satpute, S.K.; Banat, I.M.; Dhakephalkar, P.K.; Banpurkar, A.G.; Chopade, B.A. Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol. Adv. 2010, 28, 436–450. [Google Scholar] [CrossRef]
- Marzan, L.W.; Shimizu, K. Metabolic regulation of Escherichia coli and its PhoB and PhoR genes knockout mutants under phosphate and nitrogen limitations as well as at acidic condition. Microb. Cell Factories 2011, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.Z.; Silva, A.J.; Benitez, J.A. The PhoB regulatory system modulates biofilm formation and stress response in El Tor Biotype Vibrio Cholerae. FEMS Microbiol. Lett. 2010, 302, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.W.; Tifrea, D.F.; Harwood, C.S. A Chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc. Natl. Acad. Sci. USA 2005, 102, 14422–14427. [Google Scholar] [CrossRef]
- Park, S.; Kelley, K.A.; Vinogradov, E.; Solinga, R.; Weidenmaier, C.; Misawa, Y.; Lee, J.C. Characterization of the structure and biological functions of a capsular polysaccharide produced by Staphylococcus saprophyticus. J. Bacteriol. 2010, 192, 4618–4626. [Google Scholar] [CrossRef] [PubMed]
- Shang, N.; Xu, R.; Li, P. Structure characterization of an exopolysaccharide produced by Bifidobacterium animalis RH. Carbohydr. Polym. 2013, 91, 128–134. [Google Scholar] [CrossRef]
- Papageorgiou, S.K.; Kouvelos, E.P.; Favvas, E.P.; Sapalidis, A.A.; Romanos, G.E.; Katsaros, F.K. Metal–Carboxylate interactions in metal–alginate complexes studied with FTIR spectroscopy. Carbohydr. Res. 2010, 345, 469–473. [Google Scholar] [CrossRef]
- Guo, L.; Zhu, W.; Xu, F.; Liu, M.; Xie, Y.; Zhang, J. Optimized ultrasonic-assisted extraction of polysaccharides from Cyclina sinensis and evaluation of antioxidant activities in vitro. CyTA-J. Food 2014, 12, 32–39. [Google Scholar] [CrossRef]
- Liu, S.; Ai, Z.; Qu, F.; Chen, Y.; Ni, D. Effect of steeping temperature on antioxidant and inhibitory activities of green tea extracts against α-Amylase, α-Glucosidase and intestinal glucose uptake. Food Chem. 2017, 234, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Luo, Z.; Liu, D.; Ning, Z.; Yang, J.; Ren, J. Structure characterization of a novel polysaccharide from Dictyophora Indusiata and its macrophage immunomodulatory activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, Deletions and Gene Fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-Seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, Y.; Shan, K.; Liu, W.; Xi, C.; Zhang, Y.; Wang, W.; Wang, C.; Cao, R.; Zhu, W.; Wang, H.; et al. Effect of Different Initial Fermentation pH on Exopolysaccharides Produced by Pseudoalteromonas agarivorans Hao 2018 and Identification of Key Genes Involved in Exopolysaccharide Synthesis via Transcriptome Analysis. Mar. Drugs 2022, 20, 89. https://doi.org/10.3390/md20020089
Ju Y, Shan K, Liu W, Xi C, Zhang Y, Wang W, Wang C, Cao R, Zhu W, Wang H, et al. Effect of Different Initial Fermentation pH on Exopolysaccharides Produced by Pseudoalteromonas agarivorans Hao 2018 and Identification of Key Genes Involved in Exopolysaccharide Synthesis via Transcriptome Analysis. Marine Drugs. 2022; 20(2):89. https://doi.org/10.3390/md20020089
Chicago/Turabian StyleJu, Yuhao, Kai Shan, Wenlin Liu, Chenxiang Xi, Yiling Zhang, Wei Wang, Chunlei Wang, Ruiwen Cao, Wenxing Zhu, Haiyong Wang, and et al. 2022. "Effect of Different Initial Fermentation pH on Exopolysaccharides Produced by Pseudoalteromonas agarivorans Hao 2018 and Identification of Key Genes Involved in Exopolysaccharide Synthesis via Transcriptome Analysis" Marine Drugs 20, no. 2: 89. https://doi.org/10.3390/md20020089
APA StyleJu, Y., Shan, K., Liu, W., Xi, C., Zhang, Y., Wang, W., Wang, C., Cao, R., Zhu, W., Wang, H., Zhao, Y., & Hao, L. (2022). Effect of Different Initial Fermentation pH on Exopolysaccharides Produced by Pseudoalteromonas agarivorans Hao 2018 and Identification of Key Genes Involved in Exopolysaccharide Synthesis via Transcriptome Analysis. Marine Drugs, 20(2), 89. https://doi.org/10.3390/md20020089