
Cosmetic Application of Cyanobacteria Extracts with a Sustainable Vision to Skincare: Role in the Antioxidant and Antiaging Process

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
2.1. Cytotoxicity
2.2. Phytochemical Analysis
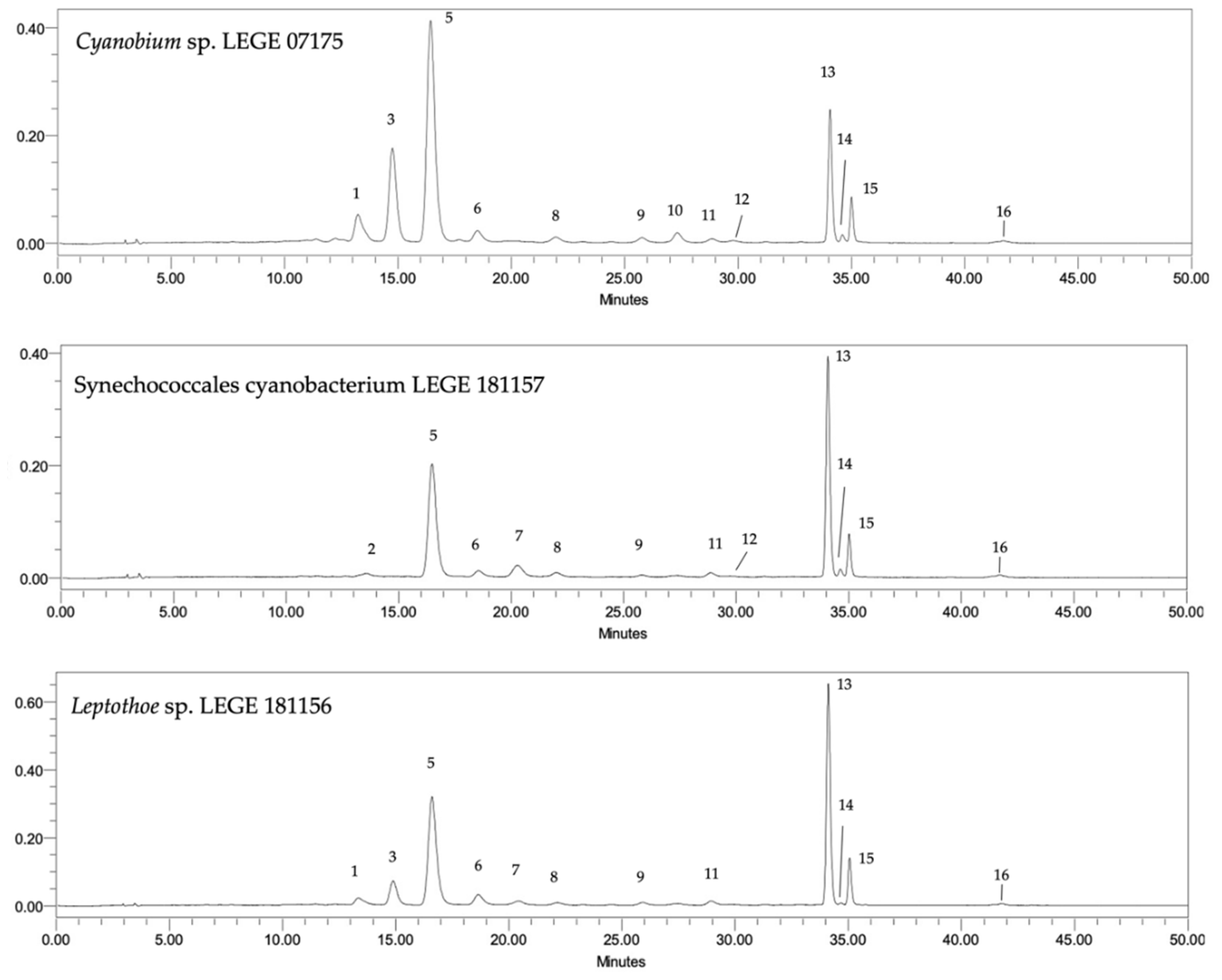
2.2.1. Carotenoids
2.2.2. Total Phenolic Content (TPC)
2.2.3. Total Proteins
2.2.4. Phycobiliproteins (PBPs)
2.3. Biological Activities
2.4. Enzymes Inhibition
2.4.1. Hyaluronidase (Hase) Inhibition
2.4.2. Elastase Inhibition
2.4.3. Tyrosinase Inhibition
2.4.4. Collagenase Inhibition
3. Discussion
3.1. Phytochemical Analysis
3.1.1. Carotenoids and Chlorophylls
3.1.2. Total Phenolic Content (TPC)
3.1.3. Proteins
3.1.4. Phycobiliproteins (PBPs)
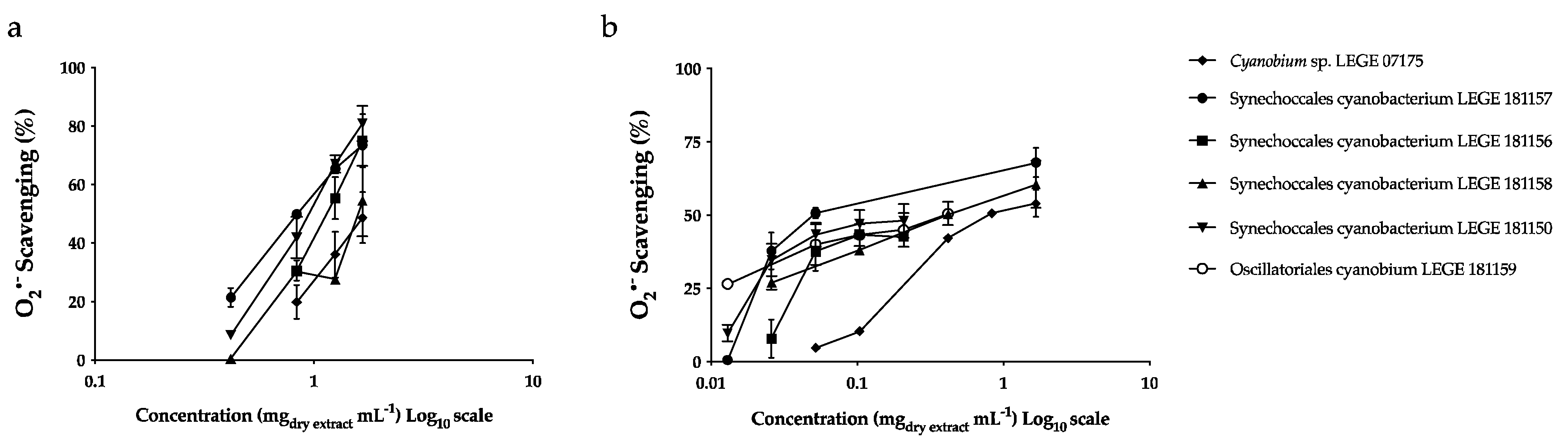
3.1.5. Antioxidant Potential
3.1.6. Inhibition of Metalloproteinases (MMPs)
4. Materials and Methods
4.1. Cyanobacteria Strains and Biomass Production
4.2. Extract Preparation
4.3. Phytochemical Analysis
4.3.1. Determination of Pigments Profile by HPLC-PDA
4.3.2. Total Phenolic Content (TPC)
4.3.3. Total Proteins
4.3.4. Phycobiliproteins
4.4. Cell Assays
4.4.1. Cell Culture
4.4.2. Cytotoxicity—MTT Assay
4.5. Biological Activities
4.5.1. Superoxide Anion Radical (O2•−) Scavenging
4.5.2. Hyaluronidase Inhibition
4.5.3. Elastase Inhibition
4.5.4. Collagenase Inhibition
4.5.5. Tyrosinase Inhibition
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morone, J.; Lopes, G.; Oliveira, B.; Vasconcelos, V.; Martins, R. Cyanobacteria in Cosmetics: A Natural Alternative for Anti-Aging Ingredients; Academic Press: Cambridge, MA, USA, 2022; pp. 257–286. [Google Scholar]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—New bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Chen, L.; Li, D.; Song, L.; Hu, C.; Wang, G.; Lui, Y. Effects of Salt Stress on Carbohydrate Metabolism in Desert Soil Alga Microcoleus vaginatus Gom. J. Integr. Plant Biol. 2006, 48, 914–919. [Google Scholar] [CrossRef]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria and microalgae bioactive compounds in skin-ageing: Potential to restore extracellular matrix filling and overcome hyperpigmentation. J. Enzym. Inhib. Med. Chem. 2021, 36, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Guedes, A.; Vasconcelos, V.; Lopes, G. Anti-inflammatory compounds from cyanobacteria. In The Pharmacological Potential of Cyanobacteria; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Assunção, J.; Amaro, H.; Malcata, F.; Guedes, A. Cyanobacterial pigments: Photosynthetic function and biotechnological purposes. In The Pharmacological Potential of Cyanobacteria; Academic Press: Cambridge, MA, USA, 2022; pp. 201–256. [Google Scholar]
- Singh, S.K.; Kaur, R.; Bansal, A.; Kapur, S.; Sundaram, S. Biotechnological exploitation of cyanobacteria and microalgae for bioactive compounds. In Biotechnological Production of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Nowruzi, B.; Sarvari, G.; Blanco, S. The cosmetic application of cyanobacterial secondary metabolites. Algal Res. 2020, 49, 101959. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef] [PubMed]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef]
- Tarasuntisuk, S.; Patipong, T.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Inhibitory effects of mycosporine-2-glycine isolated from a halotolerant cyanobacterium on protein glycation and collagenase activity. Lett Appl. Microbiol. 2018, 67, 314–320. [Google Scholar] [CrossRef]
- Keller, L.; Canuto, K.M.; Liu, C.; Suzuki, B.M.; Almaliti, J.; Sikandar, A.; Naman, C.B.; Glukhov, E.; Luo, D.; Duggan, B.M.; et al. Tutuilamides A-C: Vinyl-Chloride-Containing Cyclodepsipeptides from Marine Cyanobacteria with Potent Elastase Inhibitory Properties. ACS Chem. Biol. 2020, 15, 751–757. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Koketsu, M. Isolation and analysis of polysaccharide showing high hyaluronidase inhibitory activity in Nostochopsis lobatus MAC0804NAN. J. Biosci. Bioeng. 2016, 121, 345–348. [Google Scholar] [CrossRef]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria Secondary Metabolites as Biotechnological Ingredients in Natural Anti-Aging Cosmetics: Potential to Overcome Hyperpigmentation, Loss of Skin Density and UV Radiation-Deleterious Effects. Mar. Drugs 2022, 20, 183. [Google Scholar] [CrossRef]
- CODIF. Phormiskin Bioprotech G. Available online: https://www.codif-tn.com/en/principesactifs/phormiskin-bioprotech-g/ (accessed on 1 May 2022).
- Gao, X.; Jing, X.; Liu, X.; Lindblad, P. Biotechnological Production of the Sunscreen Pigment Scytonemin in Cyanobacteria: Progress and Strategy. Mar. Drugs 2021, 19, 129. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Silva, M.; Vasconcelos, V. The Pharmacological Potential of Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of Filamentous and Picoplanktonic Cyanobacteria for Cosmetic Applications: Potential to Improve Skin Structure and Preserve Dermal Matrix Components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Kusama, Y.; Inoue, S.; Jimbo, H.; Takaichi, S.; Sonoike, K.; Hihara, Y.; Nishiyama, Y. Zeaxanthin and Echinenone Protect the Repair of Photosystem II from Inhibition by Singlet Oxygen in Synechocystis sp. PCC 6803. Plant Cell. Physiol. 2015, 56, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Krinsky, N.I.; Landrum, J.T.; Bone, R.A. Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu. Rev. Nutr. 2003, 23, 171–201. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from Cyanobacteria: A Biotechnological Approach for the Topical Treatment of Psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef]
- Palinska, K.A.; Deventer, B.; Hariri, K.; Łotocka, M. A taxonomic study on Phormidium-group (cyanobacteria) based on morphology, pigments, RAPD molecular markers and RFLP analysis of the 16S rRNA gene fragment. Fottea 2011, 11, 41–55. [Google Scholar] [CrossRef]
- Amaro, H.M.; Fernandes, F.; Valentao, P.; Andrade, P.B.; Sousa-Pinto, I.; Malcata, F.X.; Guedes, A.C. Effect of Solvent System on Extractability of Lipidic Components of Scenedesmus obliquus (M2-1) and Gloeothece sp. on Antioxidant Scavenging Capacity Thereof. Mar. Drugs 2015, 13, 6453–6471. [Google Scholar] [CrossRef]
- Patias, L.D.; Fernandes, A.S.; Petry, F.C.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Carotenoid profile of three microalgae/cyanobacteria species with peroxyl radical scavenger capacity. Food Res. Int. 2017, 100, 260–266. [Google Scholar] [CrossRef]
- Hashtroudi, M.S.; Shariatmadari, Z.; Riahi, H.; Ghassempour, A. Analysis of Anabaena vaginicola and Nostoc calcicola from Northern Iran, as rich sources of major carotenoids. Food Chem. 2013, 136, 1148–1153. [Google Scholar] [CrossRef]
- Arnum, V.; Susan, D. Vitamin A. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons: New York, NY, USA, 2000; pp. 99–107. [Google Scholar]
- Szymanski, L.; Skopek, R.; Palusinska, M.; Schenk, T.; Stengel, S.; Lewicki, S.; Kraj, L.; Kaminski, P.; Zelent, A. Retinoic Acid and Its Derivatives in Skin. Cells 2020, 9, 2660. [Google Scholar] [CrossRef]
- Abiri, B.; Vafa, M. Antioxidant Vitamins in Acute Lymphoblastic Leukemia, 2nd ed.; Academic Press: Cambridge, MA, USA, 2021; Chapter 48. [Google Scholar]
- Steiger, S.; Schafer, L.; Sandmann, G. High-light-dependent upregulation of carotenoids and their antioxidative properties in the cyanobacterium Synechocystis PCC 6803. J. Photochem. Photobiol. 1999, 52, 14–18. [Google Scholar] [CrossRef]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Bioactive potential of Cyanobium sp. pigment-rich extracts. J. Appl. Phycol. 2020, 32, 3031–3040. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Boo, Y.C. Can Plant Phenolic Compounds Protect the Skin from Airborne Particulate Matter? Antioxidants 2019, 8, 379. [Google Scholar] [CrossRef]
- Orhan, I.E.; Khan, M.T.H. Flavonoid Derivatives as Potent Tyrosinase Inhibitors—A Survey of Recent Findings between 2008–2013. Top. Med. Chem. 2014, 14, 1486–1493. [Google Scholar] [CrossRef]
- Ahsan, H. Immunopharmacology and immunopathology of peptides and proteins in personal products. J. Immunoass. Immunochem. 2019, 40, 439–447. [Google Scholar] [CrossRef]
- Hossain, M.F.; Ratnayake, R.R.; Meerajini, K.; Wasantha Kumara, K.L. Antioxidant properties in some selected cyanobacteria isolated from fresh water bodies of Sri Lanka. Food Sci. Nutr. 2016, 4, 753–758. [Google Scholar] [CrossRef]
- Pumas, C.; Vacharapiyasophon, P.; Peerapornpisal, Y.; Leelapornpisid, P.; Boonchum, W.; Ishii, M.; Khanongnuch, C. Thermostablility of phycobiliproteins and antioxidant activity from four thermotolerant cyanobacteria. Phycol. Res. 2011, 59, 166–174. [Google Scholar] [CrossRef]
- Pez Jaeschke, D.; Rocha Teixeira, I.; Damasceno Ferreira Marczak, L.; Domeneghini Mercali, G. Phycocyanin from Spirulina: A review of extraction methods and stability. Food Res. Int. 2021, 143, 110314. [Google Scholar] [CrossRef]
- Hemlata; Fatma, T. Screening of cyanobacteria for phycobiliproteins and effect of different environmental stress on its yield. Bull. Environ. Contam. Toxicol. 2009, 83, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Rodriguez, S.; Folgueras, A.R.; Lopez-Otin, C. The role of matrix metalloproteinases in aging: Tissue remodeling and beyond. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2015–2025. [Google Scholar] [CrossRef] [PubMed]
- Del Bino, S.; Bernerd, F. Variations in skin colour and the biological consequences of ultraviolet radiation exposure. Br. J. Derm. 2013, 169, 33–40. [Google Scholar] [CrossRef]
- Pagels, F.; Almeida, C.; Vasconcelos, V.; Guedes, A.C. Cosmetic Potential of Pigments Extracts from the Marine Cyanobacterium Cyanobium sp. Mar. Drugs 2022, 20, 481. [Google Scholar] [CrossRef]
- Ramos, V.; Morais, J.; Castelo-Branco, R.; Pinheiro, A.; Martins, J.; Regueiras, A.; Pereira, A.L.; Lopes, V.R.; Frazao, B.; Gomes, D.; et al. Cyanobacterial diversity held in microbial biological resource centers as a biotechnological asset: The case study of the newly established LEGE culture collection. J. Appl. Phycol. 2018, 30, 1437–1451. [Google Scholar] [CrossRef]
- Ferreira, L.; Morais, J.; Preto, M.; Silva, R.; Urbatzka, R.; Vasconcelos, V.; Reis, M. Uncovering the Bioactive Potential of a Cyanobacterial Natural Products Library Aided by Untargeted Metabolomics. Mar. Drugs 2021, 19, 633. [Google Scholar] [CrossRef]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae; Norwegian Institute for Water Research: Oslo, Norway, 1972; Volume 15, pp. 155–164. [Google Scholar]
- Fernandes, F.; Barbosa, M.; Oliveira, A.P.; Azevedo, I.C.; Sousa-Pinto, I.; Valentão, P.; Andrade, P.B. The pigments of kelps (Ochrophyta) as part of the flexible response to highly variable marine environments. J. Appl. Phycol. 2016, 28, 3689–3696. [Google Scholar] [CrossRef]
- Barroso, M.F.; Ramalhosa, M.J.; Alves, R.C.; Dias, A.; Soares, C.M.D.; Oliva-Teles, M.T.; Delerue-Matos, C. Total antioxidant capacity of plant infusions: Assessment using electrochemical DNA-based biosensor and spectrophotometric methods. Food Control 2016, 68, 153–161. [Google Scholar] [CrossRef]
- Barbosa, M.; Fernandes, F.; Carlos, M.J.; Valentão, P.; Andrade, P.B. Adding value to marine invaders by exploring the potential of Sargassum muticum (Yendo) Fensholt phlorotannin extract on targets underlying metabolic changes in diabetes. Algal Res. 2021, 59, 102455. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentao, P. Phlorotannin extracts from fucales characterized by HPLC-DAD-ESI-MSn: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [PubMed]
- Mota, F.A.R.; Pereira, S.A.P.; Araujo, A.; Saraiva, M. Evaluation of Ionic Liquids and Ionic Liquids Active Pharmaceutical Ingredients Inhibition in Elastase Enzyme Activity. Molecules 2021, 26, 200. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.M.; Dominguez-Martin, E.M.; Nicolai, M.; Faustino, C.; Rodrigues, L.M.; Rijo, P. Screening the dermatological potential of plectranthus species components: Antioxidant and inhibitory capacities over elastase, collagenase and tyrosinase. J. Enzym. Inhib. Med. Chem. 2021, 36, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Devkota, H.P.; Takano, A.; Masudaà, K.; Nakaneà, T.; Basnet, P.; Skalko-Basnet, N. Screening of Nepalese crude drugs traditionally used to treat hyperpigmentation: In vitro tyrosinase inhibition. Int. J. Cosmet. Sci. 2008, 30, 353–360. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Peak | Compound | RT (min) | Cyanobium sp. LEGE 07175 | Synechococcales cyanobacterium LEGE 181150 | Leptothoe sp. LEGE 181156 | Synechococcales cyanobacterium LEGE 181157 | Synechococcales cyanobacterium LEGE 181158 | Oscillatoriales cyanobacterium LEGE 191159 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aqueous Extract | Acetonic Extract | Aqueous Extract | Acetonic Extract | Aqueous Extract | Acetonic Extract | Aqueous Extract | Acetonic Extract | Aqueous Extract | Acetonic Extract | Aqueous Extract | |||
| 1 | Unidentified carotenoid | 13.35 | nq | 7.28 ± 0.03 a | nd | 2.55 ± 0.04 c | nd | 3.31 ± 0.07 b | nd | nd | nd | nd | nd |
| 2 | Lutein | 13.44 | nd | nd | nd | nd | nd | nd | nd | 2.07 ± 0.70 | nd | 1.96 ± 0.15 | nd |
| 3 | Unidentified carotenoid | 14.81 | 0.53 ± 0.05 | 21.03 ± 0.40 a | 0.04 ± 0.02 | 7.75 ± 0.39 b | nq | 8.53 ± 0.03 b | nd | nd | nd | 0.77 ± 0.12 c | nd |
| 4 | Chlorophyll−a | 17.02 | nd | nd | 14.77 ± 1.21 A | 32.19 ± 1.02 | 0.81 ± 0.06 C | nd | 0.06 ± 0.002 | nd | 9.42 ± 1.01 B | 31.96 ± 2.04 | 8.69 ± 0.07 B |
| 5 | Zeaxanthin | 17.15 | 1.58 ± 0.08 | 53.08 ± 1.48 a | nd | nd | 39.85 ± 2.69 b | 24.38 ± 0.33 c | nd | nd | |||
| 6 | Chlorophyll−a derivative | 17.98 | nd | 25.27 ± 1.17 b | nd | 29.23 ± 0.69 a,b | nd | 34.81 ± 1.52 a | nd | 12.69 ± 2.04 c | nq | 21.90 ± 3.55 b | nd |
| 7 | Myxoxanthophyll | 21.07 | nd | nd | 0.29 ± 0.03 A | 8.22 ± 0.37 a | nd | 4.07 ± 0.38 b | 0.01 ± 0.002 C | 5.41 ± 0.28 a,b | 0.17 ± 0.05 A,B | 8.43 ± 1.41 a | 0.16 ± 0.01 B |
| 8 | Unidentified carotenoid | 22.18 | nd | 1.40 ± 0.18 a | nd | 2.13 ± 0.07 a | nd | 1.27 ± 0.02 b | nd | 1.15 ± 0.03 b | nd | 1.76 ± 0.41 a | nd |
| 9 | Unidentified carotenoid | 25.89 | nq | 1.21 ± 0.15 a | nd | 0.70 ± 0.01 b | nd | 1.37 ± 0.17 a | nd | 0.26 ± 0.08 b | nd | nd | nd |
| 10 | Echinenone | 27.01 | nd | 1.17 ± 0.07 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| 11 | Unidentified carotenoid | 28.93 | nd | 1.10 ± 0.04 b | nd | nd | nd | 2.02 ± 0.24 a | nd | 1.15 ± 0.14 b | nd | 1.00 ± 0.08 b | nd |
| 12 | Unidentified carotenoid | 29.7 | nq | 0.41 ± 0.16 b | nq | 1.15 ± 0.07 a | nd | nd | nd | 0.26 ± 0.09 b | nd | nd | nq |
| 13 | β−Carotene | 34.32 | 0.88 ± 0.11 B | 19.58 ± 0.64 c | 1.22 ± 0.11 A | 25.13 ± 0.75 b,c | 0.05 ± 0.002 D | 47.89 ± 2.81 a | 0.10 ± 0.002 D | 29.30 ± 0.56 b | 0.54 ± 0.05 C | 30.57 ± 1.63 b | 0.68 ± 0.01 B,C |
| 14 | Phaeophytin-a | 34.69 | nq | 5.81 ± 1.08 a | nd | 2.25 ± 0.61 b | nd | 1.81 ± 0.11 b | nd | 5.23 ± 0.27 a | nd | 6.31 ± 0.33 a | nd |
| 15 | Unidentified carotenoid | 35.05 | nq | 3.92 ± 0.18 c | nq | 5.23 ± 0.64 a,b | nq | 6.51 ± 0.24 a | nq | 3.37 ± 0.11 c | nq | 4.53 ± 0.60 b,c | nq |
| 16 | γ−Carotene | 41.78 | nd | nq | nd | 35.96 ± 3.00 a | nd | 33.52 ± 3.79 a | nd | 32.11 ± 1.47 a | nd | 26.51 ± 1.99 b | nd |
| Total carotenoids | 3.00 ±≤ 0.01 A | 115.98 ± 0.48 c | 1.55 ±≤ 0.01 B | 123.26 ± 0.85 b | 0.05 ±≤ 0.01 F | 150.15 ± 1.40 a | 0.18 ±≤ 0.01 E | 104.71 ± 0.42 d | 0.71 ±≤ 0.01 D | 113.81 ± 0.81 c | 0.84 ±≤ 0.01 C | ||
| Total chlorophylls | nd | 25.27 ± 1.17 a,b | 14.77 ± 1.21 A | 29.23 ± 0.69 a,b | 0.81 ± 0.06 C | 34.81 ± 1.52 a | nd | 12.69 ± 2.04 c | 9.42 ± 1.01 B | 21.90 ± 3.55 b,c | 8.69 ± 0.07 B | ||
| Strains | TPC (μg (GAE) mg−1 dry extract) | Total Proteins (μg (BSA) mg−1dry extract) | ||
|---|---|---|---|---|
| Aqueous Extracts | Acetonic Extracts | Aqueous Extracts | Acetonic Extracts | |
| Cyanobium sp. LEGE 07175 | 14.79 a,b ± 0.20 | 23.11 a,b ± 0.69 | 309.63 e ± 4.26 | 167.49 b ± 4.93 |
| Synechococcales cyanobacterium LEGE 181150 | 14.49 a,b ± 0.48 | 20.14 b,c ± 0.16 | 695.25 b ± 10.35 | 199.37 a ± 6.38 |
| Leptothoe sp. LEGE 181156 | 14.93 a,b ± 0.27 | 27.13 a ± 0.07 | 463.70 d ± 8.78 | 158.67 ± 2.58 |
| Synechococcales cyanobacterium LEGE 181157 | 14.92 a,b ± 0.66 | 17.86 c ± 0.06 | 760.81 a ± 15.72 | 128.84 c ± 7.12 |
| Synechococcales cyanobacterium LEGE 181158 | 15.67 a ± 0.23 | 24.71 a,b ± 0.53 | 579.99 c ± 12.36 | 209.31 a ± 4.36 |
| Oscillatoriales cyanobacterium LEGE 181159 | 11.46 b ± 0.67 | − | 550.59 c ± 18.14 | − |
| Strains | Phycobiliproteins (μg mg−1 dry aqueous extract) | ||
|---|---|---|---|
| Phycocyanin | Allophycocyanin | Phycoerythrin | |
| Cyanobium sp. LEGE 07175 | 18.21 f ± 1.36 | 11.26 e ± 1.20 | 8.06 e ± 1.17 |
| Synechococcales cyanobacterium LEGE 181150 | 222.76 a ± 0.79 | 57.29 b ± 0.17 | 17.31 d ± 0.11 |
| Leptothoe sp. LEGE 181156 | 76.10 e ± 0.13 | 37.05 d ± 0.28 | 159.05 b ± 0.14 |
| Synechococcales cyanobacterium LEGE 181157 | 117.94 d ± 0.12 | 44.57 c ± 0.28 | 275.04 a ± 0.30 |
| Synechococcales cyanobacterium LEGE 181158 | 194.43 c ± 0.39 | 44.52 c ± 0.34 | 17.42 d ± 0.22 |
| Oscillatoriales cyanobacterium LEGE 181159 | 201.15 b ± 0.78 | 65.90 a ± 0.67 | 26.33 c ± 0.16 |
| Strains | O2•− (μg mL−1) | |||
|---|---|---|---|---|
| Aqueous Extracts | Acetonic Extracts | |||
| IC25 | IC50 | IC25 | IC50 | |
| Cyanobium sp. LEGE 07175 | 248.31 c ± 1.97 | 816.22 c ± 11.48 | 921.46 b ± 77.09 | 1549.10 b ± 65.28 |
| Synechococcales cyanobacterium LEGE 181150 | 25.48 a,b ± 9.46 | 112.18 a ± 12.46 | 538.01 a ± 67.00 | 943.45 a ± 45.48 |
| Leptothoe sp. LEGE 181156 | 41.10 a,b ± 5.24 | nd | 647.24 a ± 8.95 | 1097.05 a ± 2.41 |
| Synechococcales cyanobacterium LEGE 181157 | 25.48 a,b ± 6.81 | 63.24 a ± 24.67 | 467.26 a ± 41.66 | 847.65 a ± 30.70 |
| Synechococcales cyanobacterium LEGE 181158 | 51.22 b ± 24.20 | 404.59 b ± 14.30 | 1032.15 b ± 139.10 | 1463.69 b ± 271.87 |
| Oscillatoriales cyanobacterium LEGE 181159 | 12.27 a ± 0.70 | 332.40 b ± 77.41 | − | − |
| Strains | Hyaluronidase (μg mL−1) | Elastase (μg mL−1) | Tyrosinase (μg mL−1) | |||||
|---|---|---|---|---|---|---|---|---|
| Aqueous Extracts | Acetonic Extracts | Acetonic Extracts | Acetonic Extracts | |||||
| IC25 | IC50 | IC25 | IC50 | IC25 | IC50 | IC25 | IC50 | |
| Cyanobium sp. LEGE 07175 | 488.39 a,b ± 97.20 | 894.59 c ± 78.14 | 796.94 a ± 284.55 | nd | nd | nd | 795.57 b ± 198.79 | nd |
| Synechococcales cyanobacterium LEGE 181150 | 257.05 a,b ± 1.39 | 483.86 a ± 86.71 | 595.11 a ± 4.77 | 726.29 a ± 16.56 | nd | nd | nd | nd |
| Leptothoe sp. LEGE 181156 | 800.00 c ± 152.73 | nd | nd | nd | 378.26 b ± 24.50 | nd | 133.47 a ± 35.40 | 465.92 a ± 37.59 |
| Synechococcales cyanobacterium LEGE 181157 | 188.89 a ± 86.14 | 645.06 a,b ± 118.73 | 613.47 a ± 82.241 | 859.83 a ± 122.08 | 51.43 a ± 20.02 | 380.50 ± 19.72 | 381.33 a ± 14.378 | 849.48 b ± 75.815 |
| Synechococcales cyanobacterium LEGE 181158 | 397.72 a,b ± 70.00 | 624.51 a,b ± 19.84 | 442.63 a ± 36.54 | 738.88 a ± 83.79 | 233.47 b ± 89.26 | nd | nd | nd |
| Oscillatoriales cyanobacterium LEGE 181159 | 518.69 b,c ± 15.68 | 801.30 b,c ± 116.04 | − | − | − | − | − | − |
| Strains | % Collagenase Inhibition | |||
|---|---|---|---|---|
| Aqueous Extracts | Acetonic Extracts | |||
| 0.75 mg mL−1 | 1 mg mL−1 | 0.75 mg mL−1 | 1 mg mL−1 | |
| Cyanobium sp. LEGE 07175 | nd | 8.91 b ± 2.1 | 32.88 a ± 1.70 | nd |
| Synechococcales cyanobacterium LEGE 181150 | nd | nd | 9.81 b ± 2.28 | 44.40 a ± 3.87 |
| Leptothoe sp. LEGE 181156 | nd | nd | 36.61 a ± 2.72 | 36.74 a,b ± 7.82 |
| Synechococcales cyanobacterium LEGE 181157 | nd | nd | nd | nd |
| Synechococcales cyanobacterium LEGE 181158 | nd | 25.97 a ± 6.22 | 7.30 b ± 1.58 | 24.82 b ± 4.01 |
| Oscillatoriales cyanobacterium LEGE 181159 | nd | nd | − | − |
| Strains | Solvents | |
|---|---|---|
| Acetone | Water | |
| Cyanobium sp. LEGE 07175 | 1.2 ± 0.3 | 17.2 ± 1.4 |
| Synechococcales cyanobacterium LEGE 181150 | 1.4 ± 0.4 | 39.0 ± 5.2 |
| Leptothoe sp. LEGE 181156 | 1.9 ± 0.6 | 26.6 ± 5.3 |
| Synechococcales cyanobacterium LEGE 181157 | 1.6 ± 0.6 | 31.7 ± 8.4 |
| Synechococcales cyanobacterium LEGE 181158 | 1.5 ± 0.3 | 23.4 ± 5.6 |
| Oscillatoriales cyanobacterium LEGE 181159 | − | 38.9 ± 1.0 |
| Standards | Calibration Curve | r2 |
|---|---|---|
| Lutein | y = 31,188,975x + 81,368 | 0.9987 |
| Chlorophyll-a | y =5,647,422x + 14,838 | 0.9989 |
| Zeaxanthin | y = 40,108,171x + 97,810 | 0.9994 |
| Cantaxanthin | y = 39,997,059x + 197,655 | 0.9992 |
| Myxoxantophyll | y = 30,518,380x + 5976 | 0.9993 |
| Echinenone | y = 74,770,292x + 126,878 | 0.9997 |
| β-Carotene | y = 31,852,521x + 16,127 | 0.9999 |
| γ-Carotene | y = 452,252x + 17,574 | 0.9913 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morone, J.; Lopes, G.; Morais, J.; Neves, J.; Vasconcelos, V.; Martins, R. Cosmetic Application of Cyanobacteria Extracts with a Sustainable Vision to Skincare: Role in the Antioxidant and Antiaging Process. Mar. Drugs 2022, 20, 761. https://doi.org/10.3390/md20120761
Morone J, Lopes G, Morais J, Neves J, Vasconcelos V, Martins R. Cosmetic Application of Cyanobacteria Extracts with a Sustainable Vision to Skincare: Role in the Antioxidant and Antiaging Process. Marine Drugs. 2022; 20(12):761. https://doi.org/10.3390/md20120761
Chicago/Turabian StyleMorone, Janaína, Graciliana Lopes, João Morais, Jorge Neves, Vítor Vasconcelos, and Rosário Martins. 2022. "Cosmetic Application of Cyanobacteria Extracts with a Sustainable Vision to Skincare: Role in the Antioxidant and Antiaging Process" Marine Drugs 20, no. 12: 761. https://doi.org/10.3390/md20120761
APA StyleMorone, J., Lopes, G., Morais, J., Neves, J., Vasconcelos, V., & Martins, R. (2022). Cosmetic Application of Cyanobacteria Extracts with a Sustainable Vision to Skincare: Role in the Antioxidant and Antiaging Process. Marine Drugs, 20(12), 761. https://doi.org/10.3390/md20120761