
Functional Diversification of Oyster Big Defensins Generates Antimicrobial Specificity and Synergy against Members of the Microbiota
 , , , , , , , , , and add
Show full author list
, , , , , , , , , and add
Show full author list
Abstract
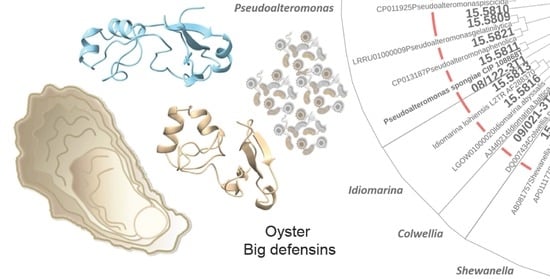
1. Introduction
2. Results
2.1. In Vivo Activity of Cg-BigDef1 on Oyster Commensal Microbiota
2.2. Establishment of a Collection of Culturable Bacteria from C. gigas Microbiota
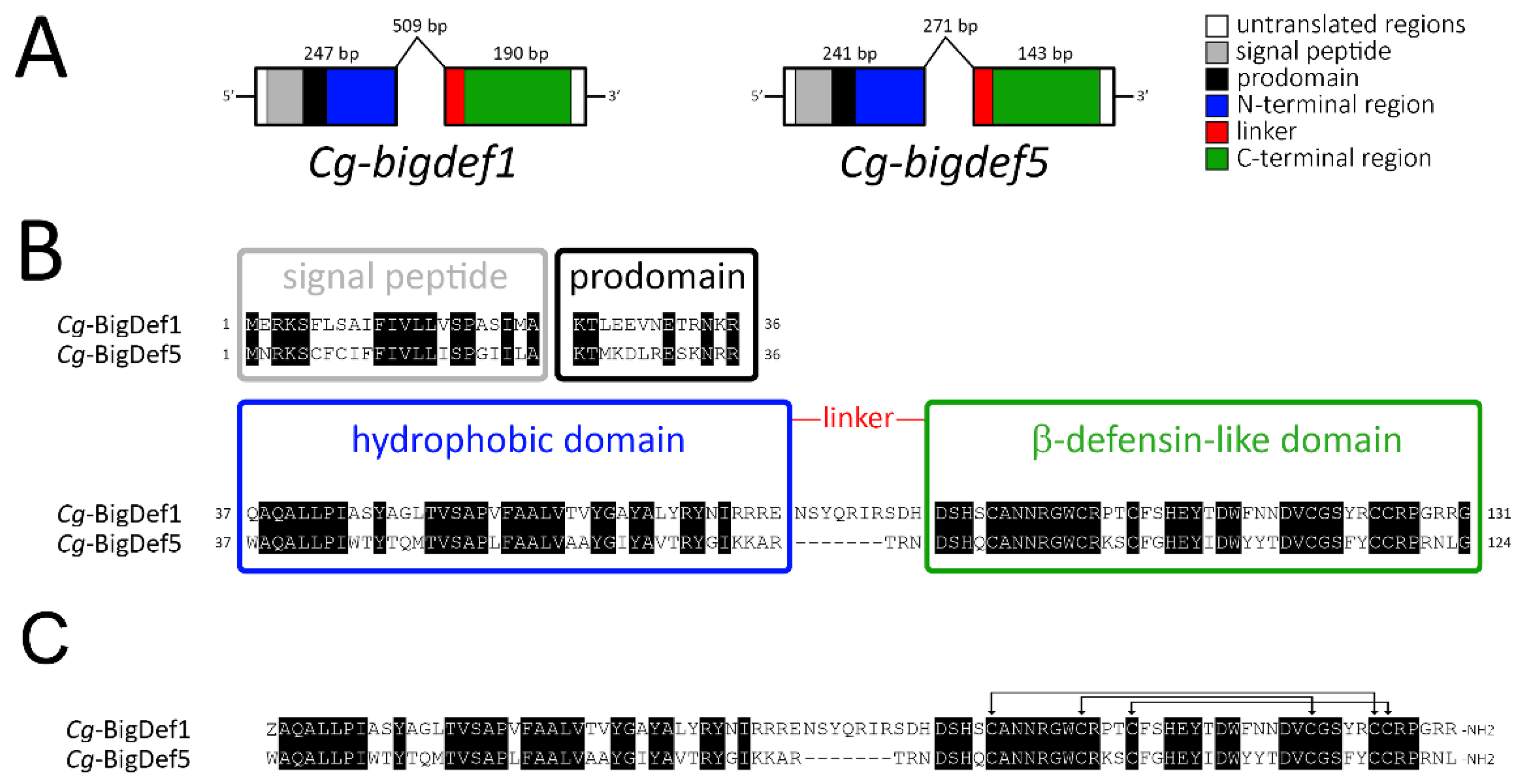
2.3. Gene Cloning and Chemical Synthesis of Cg-BigDef1 and Cg-BigDef5
2.4. Specificity, Synergy, and Complementary Broad-Spectrum Activity of Cg-BigDef1 and Cg-BigDef5 against Bacteria from the Oyster Microbiota
3. Discussion
Conclusions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Oysters
4.3. Bacterial Strains and Culture Conditions
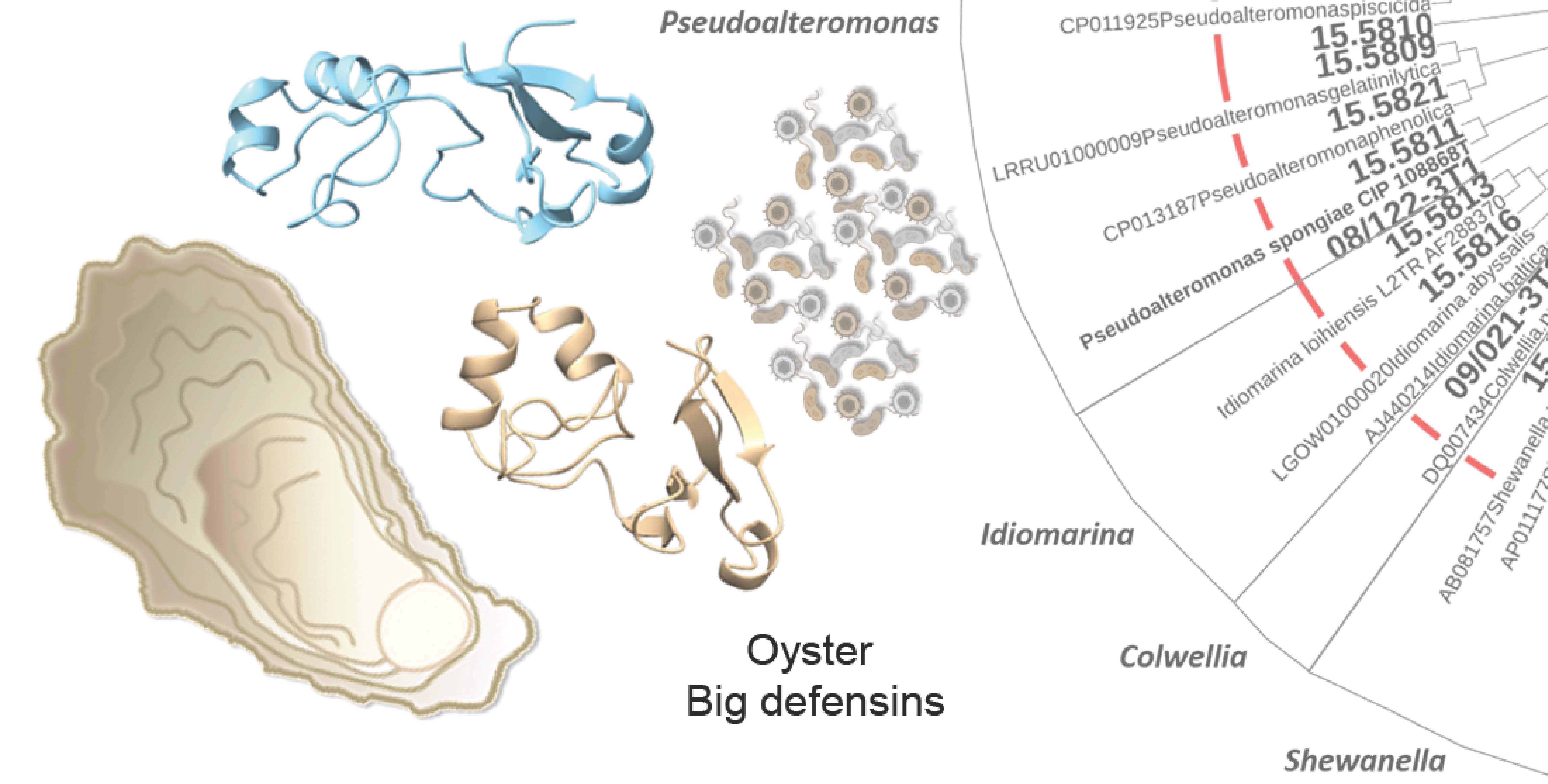
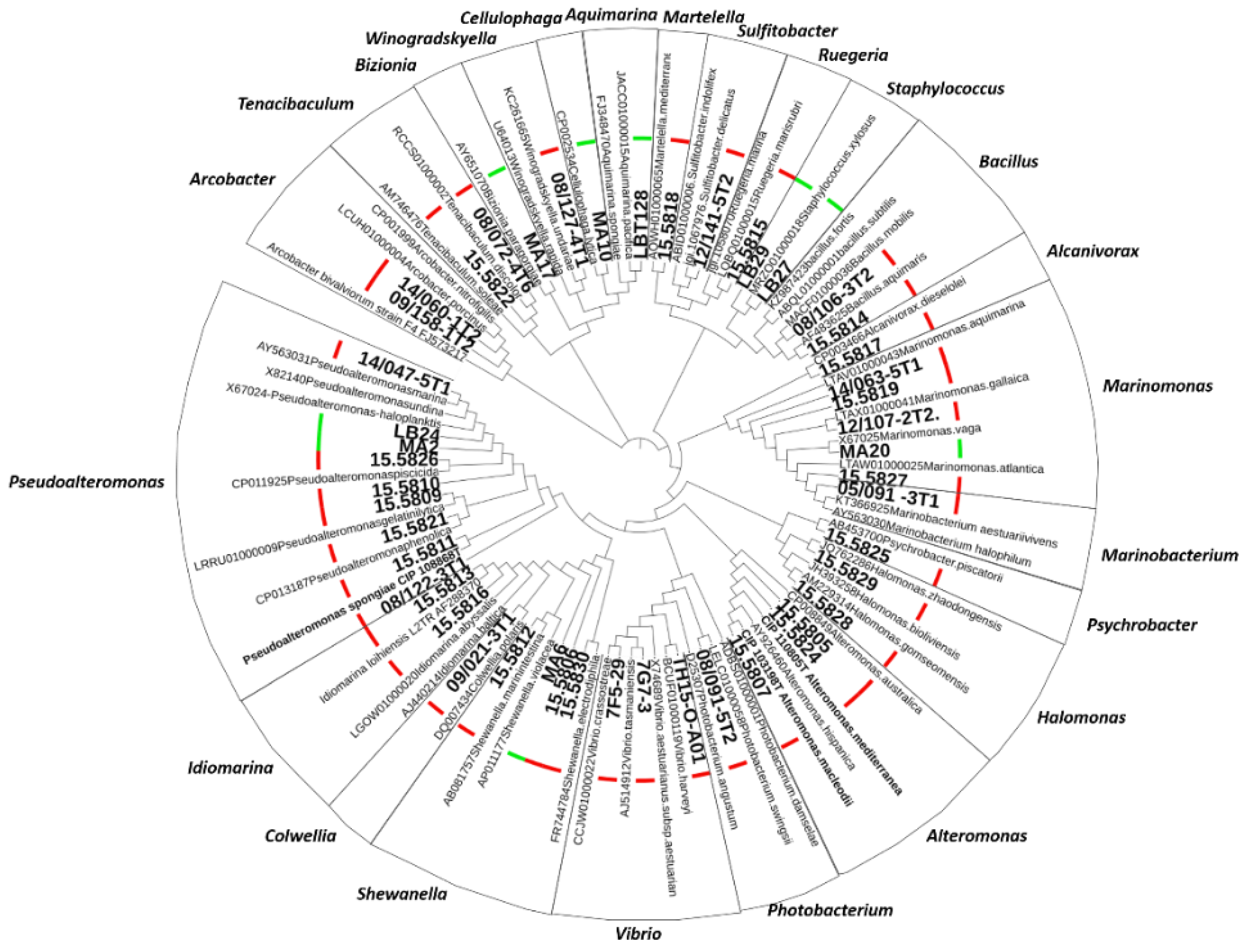
4.4. Molecular Phylogeny Based on 16S RNA
4.5. Identification of Bacterial Isolates by Matrix-Assisted Laser Desorption Ionization Mass Spectrometry (MALDI Biotyping)
4.6. Molecular Cloning and Sequence Data Analysis
4.7. Peptide Synthesis and Net Charge Calculation
4.8. Determination of the Minimum Inhibitory Concentrations (MIC), Minimum Bactericidal Concentrations (MBC), and Synergy
4.9. Microbiota Modifications Induced by Cg-BigDef1 In Vivo
4.10. 16S rRNA Metabarcoding
4.11. Quantification of Total 16S Bacterial DNA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchon, N.; Broderick, N.A.; Lemaitre, B. Gut Homeostasis in a Microbial World: Insights from Drosophila melanogaster. Nat. Rev. Microbiol. 2013, 11, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the Microbiota and the Immune System. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial Peptides and Gut Microbiota in Homeostasis and Pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Salzman, N.H. Paneth Cells, Antimicrobial Peptides and Maintenance of Intestinal Homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.G.; Zasloff, M. Antimicrobial Peptides—Or How Our Ancestors Learned to Control the Microbiome. MBio 2021, 12, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Hanson, M.A.; Kondo, S.; Erkosar, B.; Lemaitre, B. Drosophila Antimicrobial Peptides and Lysozymes Regulate Gut Microbiota Composition and Abundance. MBio 2021, 12, e0082421. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial Peptides of Innate Immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-Microbial Peptides: From Invertebrates to Vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The Immunology of Host Defence Peptides: Beyond Antimicrobial Activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Reassessing the Host Defense Peptide Landscape. Front. Chem. 2019, 7, 1–22. [Google Scholar] [CrossRef]
- Dierking, K.; Pita, L. Receptors Mediating Host-Microbiota Communication in the Metaorganism: The Invertebrate Perspective. Front. Immunol. 2020, 11, 1251. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-W.; Xu, J.-D.; Zhao, X.-F.; Vasta, G.R.; Wang, J.-X. A Shrimp C-Type Lectin Inhibits Proliferation of the Hemolymph Microbiota by Maintaining the Expression of Antimicrobial Peptides. J. Biol. Chem. 2014, 289, 11779–11790. [Google Scholar] [CrossRef]
- Login, F.H.; Balmand, S.; Vallier, A.; Vincent-Monegat, C.; Vigneron, A.; Weiss-Gayet, M.; Rochat, D.; Heddi, A. Antimicrobial Peptides Keep Insect Endosymbionts Under Control. Science 2011, 334, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Franzenburg, S.; Walter, J.; Künzel, S.; Wang, J.; Baines, J.F.; Bosch, T.C.G.; Fraune, S. Distinct Antimicrobial Peptide Expression Determines Host Species-Specific Bacterial Associations. Proc. Natl. Acad. Sci. USA 2013, 110, E3730–E3738. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, P.; Rosa, R.D.; Duperthuy, M.; de Lorgeril, J.; Bachère, E.; Destoumieux-Garzón, D. The Antimicrobial Defense of the Pacific Oyster, Crassostrea gigas. How Diversity May Compensate for Scarcity in the Regulation of Resident/Pathogenic Microflora. Front. Microbiol. 2012, 3, 160. [Google Scholar] [CrossRef]
- Bachère, E.; Rosa, R.D.; Schmitt, P.; Poirier, A.C.; Merou, N.; Charrière, G.M.; Destoumieux-Garzón, D. The New Insights into the Oyster Antimicrobial Defense: Cellular, Molecular and Genetic View. Fish Shellfish Immunol. 2015, 46, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial Peptides in Marine Invertebrate Health and Disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150300. [Google Scholar] [CrossRef] [PubMed]
- Lokmer, A.; Mathias Wegner, K. Hemolymph Microbiome of Pacific Oysters in Response to Temperature, Temperature Stress and Infection. ISME J. 2015, 9, 670–682. [Google Scholar] [CrossRef]
- Dupont, S.; Lokmer, A.; Corre, E.; Auguet, J.-C.; Petton, B.; Toulza, E.; Montagnani, C.; Tanguy, G.; Pecqueur, D.; Salmeron, C.; et al. Oyster Hemolymph Is a Complex and Dynamic Ecosystem Hosting Bacteria, Protists and Viruses. Anim. Microbiome 2020, 2, 12. [Google Scholar] [CrossRef]
- Petton, B.; Bruto, M.; James, A.; Labreuche, Y.; Alunno-Bruscia, M.; Le Roux, F. Crassostrea gigas Mortality in France: The Usual Suspect, a Herpes Virus, May Not Be the Killer in This Polymicrobial Opportunistic Disease. Front. Microbiol. 2015, 6, 686. [Google Scholar] [CrossRef]
- de Lorgeril, J.; Lucasson, A.; Petton, B.; Toulza, E.; Montagnani, C.; Clerissi, C.; Vidal-Dupiol, J.; Chaparro, C.; Galinier, R.; Escoubas, J.-M.; et al. Immune-Suppression by OsHV-1 Viral Infection Causes Fatal Bacteraemia in Pacific Oysters. Nat. Commun. 2018, 9, 4215. [Google Scholar] [CrossRef] [PubMed]
- González, R.; Gonçalves, A.T.; Rojas, R.; Brokordt, K.; Rosa, R.D.; Schmitt, P. Host Defense Effectors Expressed by Hemocytes Shape the Bacterial Microbiota From the Scallop Hemolymph. Front. Immunol. 2020, 11, 599625. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Santini, A.; Fievet, J.; Bulet, P.; Destoumieux-Garzón, D.; Bachère, E. Big Defensins, a Diverse Family of Antimicrobial Peptides That Follows Different Patterns of Expression in Hemocytes of the Oyster Crassostrea gigas. PLoS ONE 2011, 6, e25594. [Google Scholar] [CrossRef]
- Gerdol, M.; Schmitt, P.; Venier, P.; Rocha, G.; Rosa, R.D.; Destoumieux-Garzón, D. Functional Insights From the Evolutionary Diversification of Big Defensins. Front. Immunol. 2020, 11, 758. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Gao, B. Evolutionary Origin of Beta-Defensins. Dev. Comp. Immunol. 2012, 39, 79–84. [Google Scholar] [CrossRef]
- Terrier, V.P.; Adihou, H.; Arnould, M.; Delmas, A.F.; Aucagne, V. A Straightforward Method for Automated Fmoc-Based Synthesis of Bio-Inspired Peptide Crypto-Thioesters. Chem. Sci. 2015, 7, 339–345. [Google Scholar] [CrossRef]
- Loth, K.; Vergnes, A.; Barreto, C.; Voisin, S.N.; Meudal, H.; Silva, D.; Bressan, A.; Bulet, P.; Touqui, L.; Delmas, A.F.; et al. The Ancestral N-Terminal Domain of Big Defensins Drives Bacterially Triggered Assembly into Antimicrobial Nanonets Karine. MBio 2019, 10, e01821-19. [Google Scholar] [CrossRef]
- Lokmer, A.; Kuenzel, S.; Baines, J.F.; Wegner, K.M. The Role of Tissue-Specific Microbiota in Initial Establishment Success of Pacific Oysters. Environ. Microbiol. 2016, 18, 970–987. [Google Scholar] [CrossRef]
- Wegner, K.M.; Volkenborn, N.; Peter, H.; Eiler, A. Disturbance Induced Decoupling between Host Genetics and Composition of the Associated Microbiome. BMC Microbiol. 2013, 13, 252. [Google Scholar] [CrossRef]
- Lucasson, A.; Luo, X.; Mortaza, S.; de Lorgeril, J.; Toulza, E.; Petton, B.; Escoubas, J.M.; Clerissi, C.; Dégremont, L.; Gueguen, Y.; et al. A Core of Functionally Complementary Bacteria Colonizes Oysters in Pacific Oyster Mortality Syndrome. bioRxiv 2020. [Google Scholar] [CrossRef]
- Clerissi, C.; de Lorgeril, J.; Petton, B.; Lucasson, A.; Escoubas, J.M.; Gueguen, Y.; Dégremont, L.; Mitta, G.; Toulza, E. Microbiota Composition and Evenness Predict Survival Rate of Oysters Confronted to Pacific Oyster Mortality Syndrome. Front. Microbiol. 2020, 11, 311. [Google Scholar] [CrossRef] [PubMed]
- Rahman, F.U.; Andree, K.B.; Salas-Massó, N.; Fernandez-Tejedor, M.; Sanjuan, A.; Figueras, M.J.; Furones, M.D. Improved Culture Enrichment Broth for Isolation of Arcobacter-like Species from the Marine Environment. Sci. Rep. 2020, 10, 14547. [Google Scholar] [CrossRef] [PubMed]
- Travers, M.A.; Boettcher Miller, K.; Roque, A.; Friedman, C.S. Bacterial Diseases in Marine Bivalves. J. Invertebr. Pathol. 2015, 131, 11–31. [Google Scholar] [CrossRef]
- Asokan, A.; Meudal, H.; de San Nicolas, N.; Loth, K.; Destoumieux-Garzón, D.; Delmas, A.F.; Aucagne, V. Overcoming Challenges of Total Synthesis and 3D Structure Determination of Highly Hydrophobic Big Defensins. in prep.
- Schmitt, P.; De Lorgeril, J.; Gueguen, Y.; Destoumieux-Garzón, D.; Bachère, E. Expression, Tissue Localization and Synergy of Antimicrobial Peptides and Proteins in the Immune Response of the Oyster Crassostrea gigas. Dev. Comp. Immunol. 2012, 37, 363–370. [Google Scholar] [CrossRef]
- Schmitt, P.; Wilmes, M.; Pugnière, M.; Aumelas, A.; Bachère, E.; Sahl, H.G.; Schneider, T.; Destoumieux-Garzón, D. Insight into Invertebrate Defensin Mechanism of Action: Oyster Defensins Inhibit Peptidoglycan Biosynthesis by Binding to Lipid II. J. Biol. Chem. 2010, 285, 29208–29216. [Google Scholar] [CrossRef]
- Gonzalez, M.; Gueguen, Y.; Destoumieux-Garzón, D.; Romestand, B.; Fievet, J.; Pugnière, M.; Roquet, F.; Escoubas, J.-M.; Vandenbulcke, F.; Levy, O.; et al. Evidence of a Bactericidal Permeability Increasing Protein in an Invertebrate, the Crassostrea gigas Cg-BPI. Proc. Natl. Acad. Sci. USA 2007, 104, 17759–17764. [Google Scholar] [CrossRef] [PubMed]
- Salzman, N.H.; Ghosh, D.; Huttner, K.M.; Paterson, Y.; Bevins, C.L. Protection against Enteric Salmonellosis in Transgenic Mice Expressing a Human Intestinal Defensin. Nature 2003, 422, 522–526. [Google Scholar] [CrossRef]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventós, D.S.; et al. Plectasin, a Fungal Defensin, Targets the Bacterial Cell Wall Precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef]
- Turner, J.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a Cathelin-Associated Antimicrobial Peptide of Human Neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaître, B. Synergy and Remarkable Specificity of Antimicrobial Peptides In Vivo Using a Systematic Knockout Approach (ELife (2019) 8 PII: E48778). Elife 2019, 8, e44341. [Google Scholar] [CrossRef] [PubMed]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting Modern Challenges in Visualization and Analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure Visualization for Researchers, Educators, and Developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Saulnier, D.; Avarre, J.C.; Le Moullac, G.; Ansquer, D.; Levy, P.; Vonau, V. Rapid and Sensitive PCR Detection of Vibrio Penaeicida, the Putative Etiological Agent of Syndrome 93 in New Caledonia. Dis. Aquat. Organ. 2000, 40, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Moussa, M.; Cauvin, E.; Le Piouffle, A.; Lucas, O.; Bidault, A.; Paillard, C.; Benoit, F.; Thuillier, B.; Treilles, M.; Travers, M.A.; et al. A MALDI-TOF MS Database for Fast Identification of Vibrio Spp. Potentially Pathogenic to Marine Mollusks. Appl. Microbiol. Biotechnol. 2021, 105, 2527–2539. [Google Scholar] [CrossRef]
- Da Silva, P.; Strzepa, A.; Jouvensal, L.; Rahioui, I.; Gressent, F.; Delmas, A.F. A Folded and Functional Synthetic PA1b: An Interlocked Entomotoxic Miniprotein. Biopolymers 2009, 92, 436–444. [Google Scholar] [CrossRef]
- Kozlowski, L.P. IPC—Isoelectric Point Calculator. Biol. Direct 2016, 11, 55. [Google Scholar] [CrossRef]
- Suquet, M.; De Kermoysan, G.; Araya, R.G.; Queau, I.; Lebrun, L.; Souchu, P.L.; Mingant, C. Anesthesia in Pacific Oyster, Crassostrea gigas. Aquat. Living Resour. 2009, 22, 29–34. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- DI Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow Enables Reproducible Computational Workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.; Simpson, G.L. Vegan: Community Ecology Package, R Package Version 1.17–18. 2011. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 1 September 2022).
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
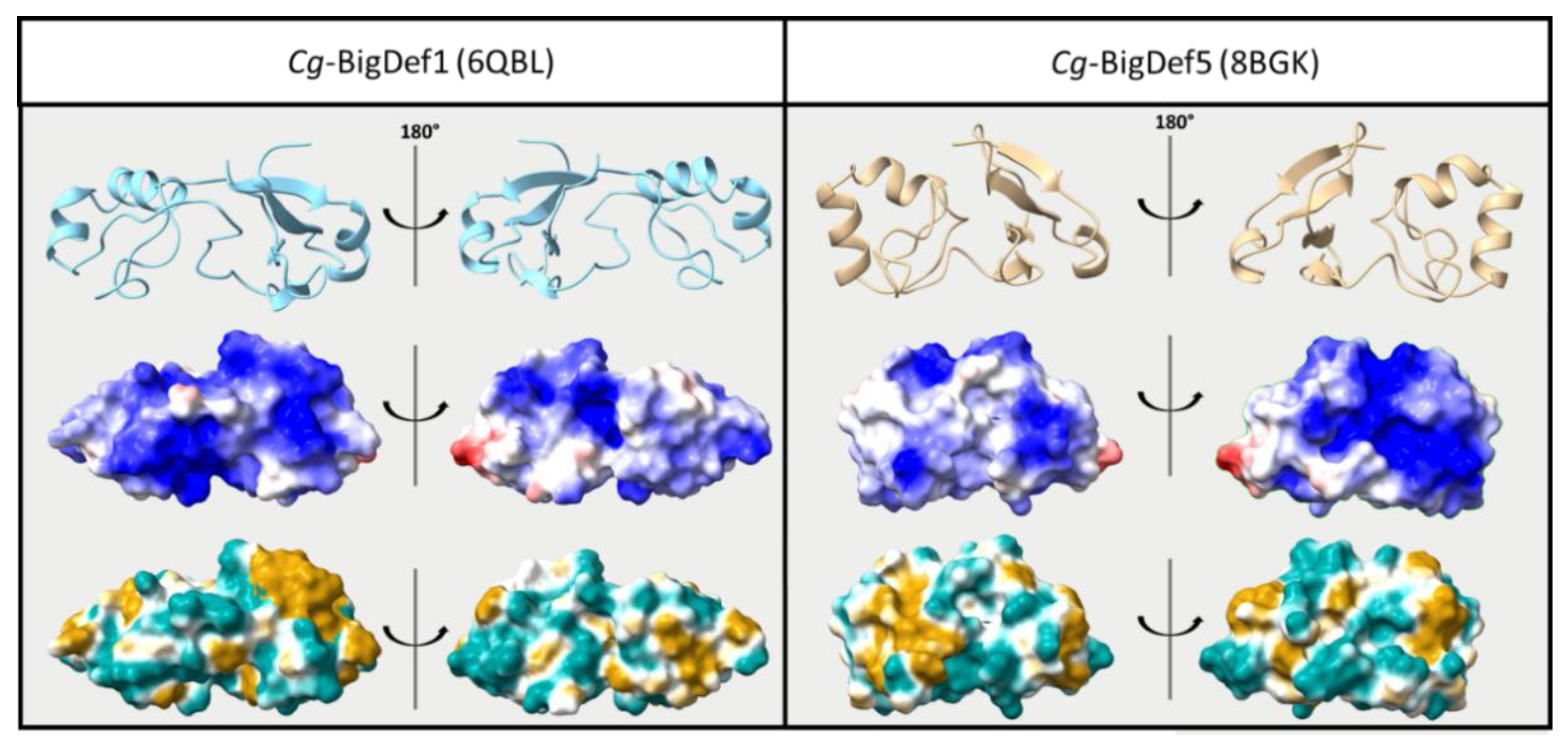{kind=link}
| Strain | Cg-BigDef1 (1–93) | Cg-BigDef5 (1–86) | FIC Index | ||
|---|---|---|---|---|---|
| MIC (µM) | MBC (µM) | MIC (µM) | MBC (µM) | ||
| Alcanivorax sp. 15.5817 | 2.50 | 5 | >10 | >10 | nt |
| Alteromonas sp. 15.5805 | 1.25 | 5 | 2.50 | >10 | 1 |
| Aquimarina sp. LTB 128 | 5 | >10 | 2.50 | >10 | nt |
| Bacillus sp. 15.5814 | 0.04 | 0.31 | 1.25 | >10 | 0.35 |
| Halomonas sp. 15.5829 | 2.50 | 10 | >10 | >10 | nt |
| Marinobacterium sp. 05.091-3T1 | >10 | >10 | 2.50 | >10 | nt |
| Marinomonas sp. 14.063 | >10 | >10 | 0.60 | >10 | nt |
| Marinomonas sp. 15.5827 | 2.50 | >10 | >10 | >10 | nt |
| Martellela sp. 15.5818 | 2.50 | >10 | 1.25 | >10 | nt |
| Pseudoalteromonas sp. 15.5809 | 10 | 10 | >10 | >10 | nt |
| Ruegeria sp. 15.5815 | >10 | >10 | 10 | >10 | nt |
| Shewanella sp. 15.5830 | 1.25 | >10 | 10 | >10 | nt |
| Sulfitobacter sp. 12.141-5T2 | >10 | >10 | 2.50 | >10 | nt |
| Tenacibaculum sp. 08.072-4T6 | 1.25 | >10 | >10 | >10 | nt |
| Winogradskyella sp. 08.27-4T1 | 1.25 | 5 | >10 | >10 | nt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De San Nicolas, N.; Asokan, A.; Rosa, R.D.; Voisin, S.N.; Travers, M.-A.; Rocha, G.; Dantan, L.; Dorant, Y.; Mitta, G.; Petton, B.; et al. Functional Diversification of Oyster Big Defensins Generates Antimicrobial Specificity and Synergy against Members of the Microbiota. Mar. Drugs 2022, 20, 745. https://doi.org/10.3390/md20120745
De San Nicolas N, Asokan A, Rosa RD, Voisin SN, Travers M-A, Rocha G, Dantan L, Dorant Y, Mitta G, Petton B, et al. Functional Diversification of Oyster Big Defensins Generates Antimicrobial Specificity and Synergy against Members of the Microbiota. Marine Drugs. 2022; 20(12):745. https://doi.org/10.3390/md20120745
Chicago/Turabian StyleDe San Nicolas, Noémie, Aromal Asokan, Rafael D. Rosa, Sébastien N. Voisin, Marie-Agnès Travers, Gustavo Rocha, Luc Dantan, Yann Dorant, Guillaume Mitta, Bruno Petton, and et al. 2022. "Functional Diversification of Oyster Big Defensins Generates Antimicrobial Specificity and Synergy against Members of the Microbiota" Marine Drugs 20, no. 12: 745. https://doi.org/10.3390/md20120745
APA StyleDe San Nicolas, N., Asokan, A., Rosa, R. D., Voisin, S. N., Travers, M.-A., Rocha, G., Dantan, L., Dorant, Y., Mitta, G., Petton, B., Charrière, G. M., Escoubas, J.-M., Boulo, V., Pouzadoux, J., Meudal, H., Loth, K., Aucagne, V., Delmas, A. F., Bulet, P., ... Destoumieux-Garzón, D. (2022). Functional Diversification of Oyster Big Defensins Generates Antimicrobial Specificity and Synergy against Members of the Microbiota. Marine Drugs, 20(12), 745. https://doi.org/10.3390/md20120745