


κ- and λ-Carrageenans from Marine Alga Chondrus armatus Exhibit Anticancer In Vitro Activity in Human Gastrointestinal Cancers Models

 ,
, 

Abstract
1. Introduction
2. Results
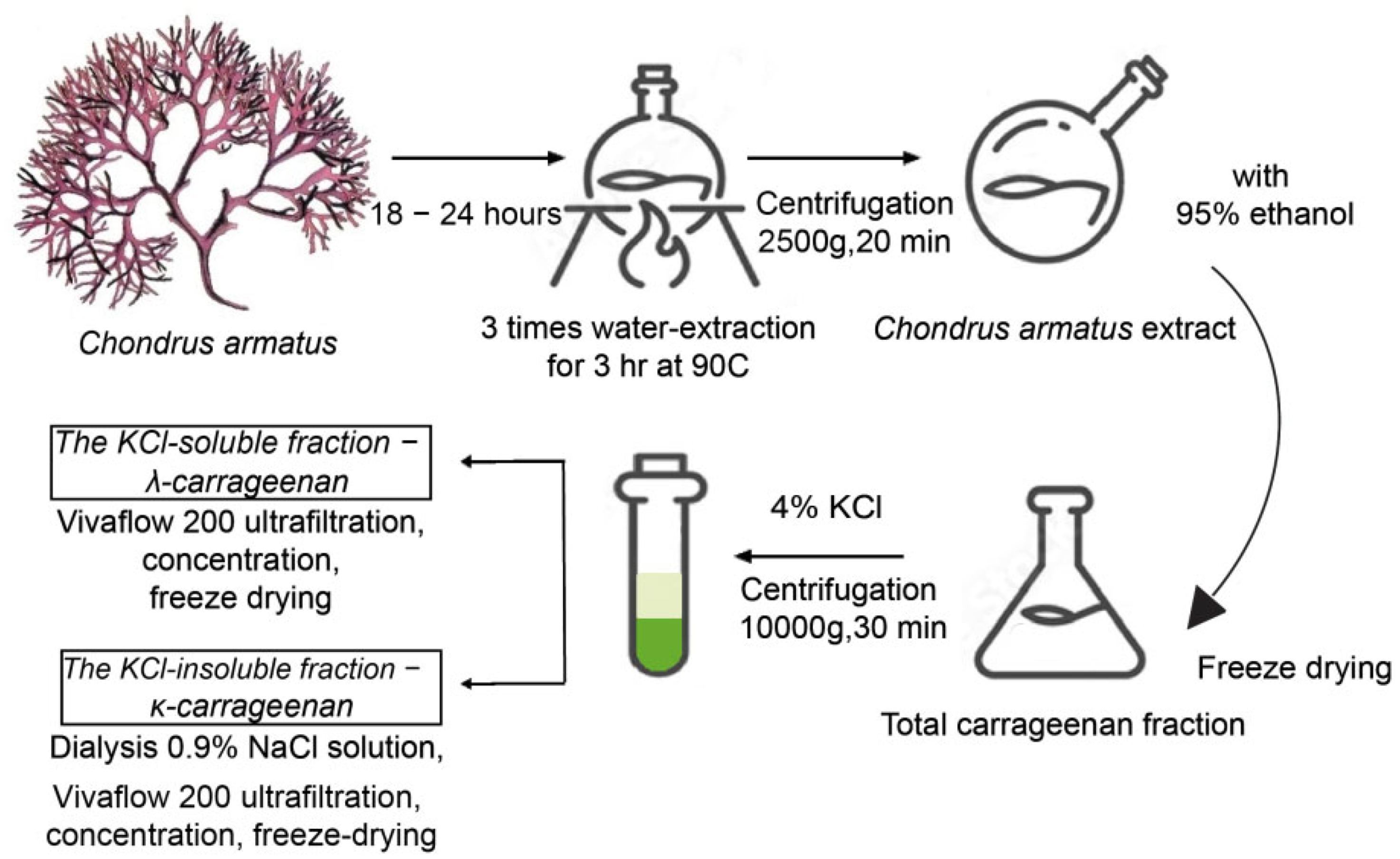
2.1. Isolation, Purification, and Structural Characterization of κ- and λ-Carrageenans from Chondrus armatus
2.2. Cytotoxicity of κo- and λo-Carrageenans
2.3. Effect of κo- and λo-Carrageenans on the Cell Cycle
2.4. κo- and λo-Carrageenans Induce Apoptosis on Human Colon Cancer and Esophageal Carcinoma Cell Lines
2.5. Gastrointestinal Cancer Cells Treated with κo- and λo-Carrageenans Demonstrated a Reduction of CDK2 and E2F2
3. Discussion
4. Materials and Methods
4.1. Obtaining Polysaccharides and Their Low Molecular Weight Derivatives
4.2. Structural Features and Molecular Weight
4.3. Cell Cultures
4.4. IC50 Concentration
4.5. Cell Cycle Assay
4.6. Detection of S-G2-M Phase with Cell-Cycle Censor GEMNN_GFP
4.7. Apoptosis Assay
4.8. Western Blot
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neergheen-Bhujun, V.; Awan, A.T.; Baran, Y.; Bunnefeld, N.; Chan, K.; Dela Cruz, T.E.; Egamberdieva, D.; Elsässer, S.; Johnson, M.V.; Komai, S.; et al. Biodiversity, drug discovery, and the future of global health: Introducing the biodiversity to biomedicine consortium, a call to action. J. Glob. Health 2017, 7, 020304. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: Updates 2020. Mar. Drugs 2020, 18, 643. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 16, 8–53. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From marine origin to therapeutics: The Antitumor Potential of marine algae-derived compounds. Front. Pharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A. Recent Updates on Marine Cancer-Preventive Compounds. Mar. Drugs 2021, 19, 558. [Google Scholar] [CrossRef] [PubMed]
- Lins, K.O.; Bezerra, D.P.; Alves, A.P.; Alencar, N.M.; Lima, M.W.; Torres, V.M.; Farias, W.R.; Pessoa, C.; de Moraes, M.O.; Costa-Lotufo, L.V. Antitumor properties of a sulfated polysaccharide from the red seaweed Champia feldmannii (Diaz-Pifferer). J. Appl. Toxicol. 2009, 29, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Campo, V.L.; Kawano, D.F.; da Silva, D.B.; Carvalho, I., Jr. Carrageenans: Biological properties, chemical modifications and structural analysis—A review. Carbohydr. Polym. 2009, 77, 167–180. [Google Scholar] [CrossRef]
- Ghanbarzadeh, M.; Golmoradizadeh, A.; Homaei, A. Carrageenans and carrageenases: Versatile polysaccharides and promising marine enzymes. Phytochem. Rev. 2018, 17, 535–571. [Google Scholar] [CrossRef]
- Hu, X.; Jiang, X.; Aubree, E.; Boulenguer, P.; Critchley, A.T. Preparation and in vivo antitumor activity of kappa-carrageenan oligosaccharides. Pharm. Biol. 2006, 44, 646–650. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Mao, X.; Peng, X.H.; Tang, S.Q. Effects of sulfate group in red seaweed polysaccharides on anticoagulant activity and cytotoxicity. Carbohydr. Polym. 2014, 101, 776–785. [Google Scholar] [CrossRef]
- Tang, F.; Chen, F.; Li, F. Preparation and potential in vivo anti-influenza virus activity of low molecular-weight kappa-carrageenans and their derivatives. J. Appl. Polym. Sci. 2013, 127, 2110–2115. [Google Scholar] [CrossRef]
- Cosenza, V.A.; Navarro, D.A.; Pujol, C.A.; Damonte, E.B.; Stortz, C.A. Partial and total C-6 oxidation of gelling carrageenans. Modulation of the antiviral activity with the anionic character. Carbohydr. Polym. 2015, 128, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos-Fidencio, G.C.; Goncalves, A.G.; Noseda, M.D.; Duarte, M.E.R.; Ducatti, D.R.B. Effects of carboxyl group on the anticoagulant activity of oxidized carrageenans. Carbohydr. Polym. 2019, 214, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Sun, Y.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xin, H.; Sheng, W.; Sun, Y.; Li, Z.; Xu, Z. In vivo growth-inhibition of S180 tumor by mixture of 5-Fu and low molecular λ-carrageenan from Chondrus ocellatus. Pharmacol. Res. 2005, 51, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Song, J. Preparation, structural characterization and in vitro anti- tumor activity of kappa-carrageenan oligosaccharide fraction from Kappaphycus striatum. J. Appl. Phycol. 2005, 17, 7–13. [Google Scholar] [CrossRef]
- de Souza, M.C.R.; Marques, C.T.; Dore, C.M.G.; da Silva, F.R.F.; Rocha, H.A.O.; Leite, E.L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef]
- Calvo, G.H.; Cosenza, V.A.; Sáenz, D.A.; Navarro, D.A.; Stortz, C.A.; Cespedes, M.A.; Mamone, L.A.; Gasas, A.G.; Di Venosa, G.M. Disaccharides obtained from carrageenans as potential antitumor agents. Sci. Rep. 2019, 9, 6654. [Google Scholar] [CrossRef] [PubMed]
- Suganya, A.M.; Sanjivkumar, M.; Chandran, M.N.; Palavesam, A.; Immanuel, G. Pharmacological importance of sulphated polysaccharide carrageenan from red seaweed Kappaphycus alvarezii in comparison with commercial carrageenan. Biomed. Pharm. 2016, 84, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Cicinskas, E.; Begun, M.A.; Vikhareva, V.V.; Karetin, Y.A.; Kalitnik, A.A. Immunological effects of Chondrus armatus carrageenans and their low molecular weight degradation products. J. Biomed. Mater. Res. Part A 2021, 109, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Kalitnik, A.A.; Marcov, P.A.; Anastyuk, S.D.; Barabanova, A.O.; Glazunov, V.P.; Popov, S.V.; Ovodov, Y.S.; Yermak, I.M. Gelling polysaccharide from Chondrus armatus and its oligosaccharides: The structural peculiarities and anti-inflammatory activity. Carbohydr. Polym. 2015, 115, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Cicinskas, E.; Begun, M.A.; Tiasto, V.A.; Belousov, A.S.; Vikhareva, V.V.; Mikhailova, V.A.; Kalitnik, A.A. In vitro antitumor and immunotropic activity of carrageenans from red algae Chondrus armatus and their low-molecular weight degradation products. J. Biomed. Mater. Res. A 2020, 108, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Vikhareva, V.V.; Leontieva, E.A.; Begun, M.A.; Cicinskas, E.; Kalitnik, A.A.; Khotimchenko, M.Y. Antitumor activity of sulfated polysaccharides of Chondrus armatus alga of the Sea of Japan. Sib. Med. Rev. 2020, 3, 78–85. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Internat. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Yermak, I.M.; Kim, Y.H.; Titlynov, E.A.; Isakov, V.V.; Solov’eva, T.F. Chemical structure and gel properties of carrageenans from algae belonging to the Gigartinaceae and Tichocarpaceae, collected from the Russian Pacific coast. J. Appl. Phycol. 1999, 11, 41–48. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FTRaman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef]
- van de Velde, F.; Knutsen, S.H.; Usov, A.I.; Rollemay, H.S.; Cerezo, A.S. 1H and 13C high resolution NMR spectroscopy of carrageenans: Application in research and industry. Trends Food Sci. Technol. 2002, 13, 73–92. [Google Scholar] [CrossRef]
- Kalitnik, A.A. Low Molecular Weight Derivatives of Ionic Polysaccharides. Structure and Properties. Ph.D. Thesis, Pacific Institute of Bioorganic Chemistry, Far Eastern Branch of the Russian Academy of Sciences, Moscow, Russia, 2013. Available online: https://static.freereferats.ru/_avtoreferats/01006573944.pdf (accessed on 7 November 2022). (In Russian).
- Sakaue-Sawano, A.; Ohtawa, K.; Hama, H.; Kawano, M.; Ogawa, M.; Miyawaki, A. Tracing the silhouette of individual cells in S/G2/M phases with fluorescence. ACS Chem. Biol. 2008, 15, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Tecson, M.G.; Abad, L.V.; Ebajo, V.D.; Camacho, D.H., Jr. Ultrasound-assisted depolymerization of kappa-carrageenan and characterization of degradation product. Ultrason Sonochem. 2021, 73, 105540. [Google Scholar] [CrossRef]
- Ariffin, S.H.; Yeen, W.W.; Abidin, I.Z.; Wahab, R.M.A.; Ariffin, Z.Z.; Senafi, S. Cytotoxicity effect of degraded and undegraded kappa and iota carrageenan in human intestine and liver cell lines. BMC Complement Altern. Med. 2014, 14, 508. [Google Scholar] [CrossRef]
- Jin, Z.; Han, Y.X.; Han, X.R. Degraded iota-carrageenan can induce apoptosis in human osteosarcoma cells via the Wnt/β-catenin signaling pathway. Nutr. Cancer 2013, 65, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, S.N.; Ermakova, S.P.; Zvyagintseva, T.N.; Stonik, V.A. Anticancer and cancer preventive properties of marine polysaccharides: Some results and prospects. Mar. Drugs 2013, 11, 4876–4901. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Miyake, M.; Kobayashi, D.; Hazama, A. Carrageenan delays cell cycle progression in human cancer cells in vitro demonstrated by FUCCI imaging. BMC Complement Altern. Med. 2016, 4, 16–270. [Google Scholar] [CrossRef]
- Ling, N. Growth inhibition and cell cycle arrest of Kappa-Selenocarrageenan and paclitaxel on HepG2 cells. Adv. Mat. Res. 2012, 343–344, 535–537. [Google Scholar] [CrossRef]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.T.; Zhou, T.T.; Liu, B. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Anand, J.; Sathuvan, M.; Babu, G.V.; Sakthivel, M.; Palani, P.; Nagaraj, S. Bioactive potential and composition analysis of sulfated polysaccharide from Acanthophora specifera (Vahl) Borgeson. Int. J. Biol. Macromol. 2018, 111, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Jazzara, M.; Ghannam, A.; Soukkarieh, C.; Murad, H. Anti-Proliferative Activity of λ-Carrageenan through the Induction of Apoptosis in Human Breast Cancer Cells. Iran. J. Cancer Prev. 2016, 9, 3836. [Google Scholar] [CrossRef] [PubMed]
- Donnellan, R.; Chetty, R. Cyclin E in human cancers. FASEB J. 1999, 13, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.G.; Schneider-Broussard, R. Role of E2F in cell cycle control and cancer. Front. Biosci.-Landmark 1998, 3, 447–448. [Google Scholar] [CrossRef]
- Jurisic, V.; Bumbasirevic, V. In vitro assays for cell death determination. Arch. Onchol. 2008, 16, 49–54. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Carrageenan | Monosaccharide Unit | 13C Signals (ppm.) | |||||
|---|---|---|---|---|---|---|---|
| C-1 | C-2 | C-3 | C-4 | C-5 | C-6 | ||
| κ [28] | G4S | 102.5 | 69.6 | 78.9 | 74.1 | 74.8 | 61.3 |
| κo [29] | 103.1 | 70.09 | 79.04 | 74.2 | 75.5 | 61.9 | |
| κ [28] | DA | 95.3 | 69.9 | 79.2 | 78.3 | 76.8 | 69.5 |
| κo [29] | 95.82 | 70.48 | 79.83 | 77.8 | 77.4 | 70.2 | |
| λ [28] | G2S | 103.4 | 77.4 | 75.8 | 64.2 | 74.2 | 61.3 |
| λo | 101.1, 102.5, 103.1, 104.0 | 78.7, 76.4 | 76.3 | 64.7 | 74.4, 74.2 | 60.4, 60.6 | |
| λ [28] | D2S,6S | 91.6 | 74.8 | 69.5 | 80.3 | 68.7 | 68.1 |
| λo | 91.6 | 75.1 | 69.6 | 80.4 | 68.9, 68.6 | 68.1 | |
| Mn | Mw | Mw/Mn | |
|---|---|---|---|
| κ-polysaccharide | 678.8 kDa | 783.9 kDa | 3.09 |
| κo-oligosaccharide | 0.9 kDa | 3.93 kDa | 3.96 |
| λ-polysaccharide | 844.2 kDa | 851.4 kDa | 4.19 |
| λo-oligosaccharide | 21.4 kDa | 69.4 kDa | 3.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiasto, V.A.; Goncharov, N.V.; Romanishin, A.O.; Zhidkov, M.E.; Khotimchenko, Y.S. κ- and λ-Carrageenans from Marine Alga Chondrus armatus Exhibit Anticancer In Vitro Activity in Human Gastrointestinal Cancers Models. Mar. Drugs 2022, 20, 741. https://doi.org/10.3390/md20120741
Tiasto VA, Goncharov NV, Romanishin AO, Zhidkov ME, Khotimchenko YS. κ- and λ-Carrageenans from Marine Alga Chondrus armatus Exhibit Anticancer In Vitro Activity in Human Gastrointestinal Cancers Models. Marine Drugs. 2022; 20(12):741. https://doi.org/10.3390/md20120741
Chicago/Turabian StyleTiasto, Vladlena A., Nikolay V. Goncharov, Alexander O. Romanishin, Maxim E. Zhidkov, and Yuri S. Khotimchenko. 2022. "κ- and λ-Carrageenans from Marine Alga Chondrus armatus Exhibit Anticancer In Vitro Activity in Human Gastrointestinal Cancers Models" Marine Drugs 20, no. 12: 741. https://doi.org/10.3390/md20120741
APA StyleTiasto, V. A., Goncharov, N. V., Romanishin, A. O., Zhidkov, M. E., & Khotimchenko, Y. S. (2022). κ- and λ-Carrageenans from Marine Alga Chondrus armatus Exhibit Anticancer In Vitro Activity in Human Gastrointestinal Cancers Models. Marine Drugs, 20(12), 741. https://doi.org/10.3390/md20120741