



A New Perspective: Revealing the Algicidal Properties of Bacillus subtilis to Alexandrium pacificum from Bacterial Communities and Toxins


Abstract
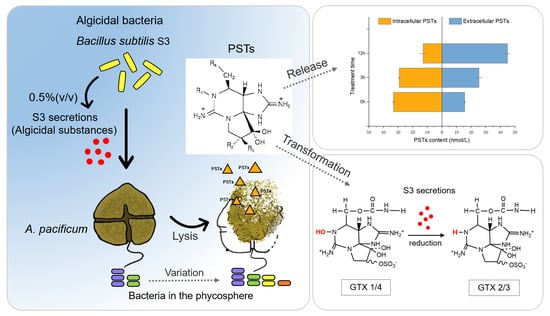
1. Introduction
2. Results
2.1. Algicidal Bacteria Screening
2.2. Algicidal Effect of B. subtilis S3 on A. pacificum
2.3. Observation of the A. pacificum Cell Morphology
2.4. Changes in the Bacterial Community during the Algicidal Process
2.5. Changes in the Contents and Toxicity of PSTs during the Algicidal Process
2.5.1. Extracellular and Intracellular PSTs Contents
2.5.2. Total PSTs Content, Toxicity, and Profile
3. Discussion
3.1. Algicidal Effect of B. subtilis S3 on A. pacificum
3.2. Impact on Bacterial Community during the Algicidal Process
3.3. Impact on PSTs during the Algicidal Process
3.3.1. Extracellular and Intracellular PSTs
3.3.2. Total PSTs Content, Toxicity, and Profile
4. Materials and Methods
4.1. Cultivation of Algae and Bacteria
4.2. Screening of Algicidal Bacteria
4.3. Algicidal Effects of B. subtilis S3
4.4. Observation of Algal Cell Morphology
4.5. Determination of Bacterial Community Structure
4.5.1. DNA Extraction for Sequencing
4.5.2. Processing of 16S rDNA Genes Sequence Data
4.6. Detection of PSTs
4.6.1. Extraction of PSTs
4.6.2. LC-MS/MS Analysis
4.7. Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [PubMed]
- Balaji-Prasath, B.; Wang, Y.; Su, Y.P.; Hamilton, D.P.; Lin, H.; Zheng, L.; Zhang, Y. Methods to control harmful algal blooms: A review. Environ. Chem. Lett. 2022, 20, 3133–3152. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal Revision of the Alexandrium tamarense Species Complex (Dinophyceae) Taxonomy: The Introduction of Five Species with Emphasis on Molecular-based (rDNA) Classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, A.; Nagai, S.; Kudo, I.; Tanizawa, K. Viability of Alexandrium tamarense cysts in the sediment of Funka Bay, Hokkaido, Japan: Over a hundred year survival times for cysts. Harmful Algae 2012, 16, 81–88. [Google Scholar] [CrossRef]
- Oyaneder-Terrazas, J.; Figueroa, D.; Araneda, O.F.; García, C. Saxitoxin Group Toxins Accumulation Induces Antioxidant Responses in Tissues of Mytilus chilensis, Ameghinomya antiqua, and Concholepas concholepas during a Bloom of Alexandrium pacificum. Antioxidants 2022, 11, 392. [Google Scholar] [CrossRef]
- Fertouna-Bellakhal, M.; Dhib, A.; Fathalli, A.; Bellakhal, M.; Chomérat, N.; Masseret, E.; Laabir, M.; Turki, S.; Aleya, L. Alexandrium pacificum Litaker sp. nov (Group IV): Resting cyst distribution and toxin profile of vegetative cells in Bizerte Lagoon (Tunisia, Southern Mediterranean Sea). Harmful Algae 2015, 48, 69–82. [Google Scholar] [CrossRef]
- Gu, H.; Zeng, N.; Liu, T.; Yang, W.; Müller, A.; Krock, B. Morphology, toxicity, and phylogeny of Alexandrium (Dinophyceae) species along the coast of China. Harmful Algae 2013, 27, 68–81. [Google Scholar] [CrossRef]
- Han, M.; Lee, H.; Anderson, D.M.; Kim, B. Paralytic shellfish toxin production by the dinoflagellate Alexandrium pacificum (Chinhae Bay, Korea) in axenic, nutrient-limited chemostat cultures and nutrient-enriched batch cultures. Mar. Pollut. Bull. 2016, 104, 34–43. [Google Scholar] [CrossRef]
- Kao, C.; Nishiyama, A. Actions of Saxitoxin on Peripheral Neuromuscular Systems. J. Physiol.-Lond. 1965, 180, 50–66. [Google Scholar]
- Raposo, M.I.C.; Gomes, M.T.S.R.; Botelho, M.J.; Rudnitskaya, A. Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review. Toxins 2020, 12, 344. [Google Scholar] [CrossRef]
- Hadjadji, I.; Laabir, M.; Frihi, H.; Collos, Y.; Shao, Z.J.; Berrebi, P.; Abadie, E.; Amzil, Z.; Chomérat, N.; Rolland, J.L.; et al. Unsuspected intraspecific variability in the toxin production, growth and morphology of the dinoflagellate Alexandrium pacificum R.W. Litaker (Group IV) blooming in a South Western Mediterranean marine ecosystem, Annaba Bay (Algeria). Toxicon 2020, 180, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Shumway, S.E.; Barter, J.; Sherman-Caswell, S. Auditing the Impact of Toxic Algal Blooms on Oysters. Environ. Audit. 1990, 2, 41–56. [Google Scholar]
- Suleiman, M.; Jelip, J.; Rundi, C.; Chua, T.H. Case Report: Paralytic Shellfish Poisoning in Sabah, Malaysia. Am. J. Trop. Med. Hyg. 2017, 97, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Mayali, X.; Azam, F. Algicidal bacteria in the sea and their impact on algal blooms. Eukaryot. Microbiol. 2002, 51, 139–144. [Google Scholar] [CrossRef]
- Park, S.C.; Lee, J.K.; Kim, S.W.; Park, Y.; Terence, E. Selective Algicidal Action of Peptides against Harmful Algal Bloom Species. PLoS ONE 2011, 6, e26733. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Chi, Y.; Wu, D.; Dai, X.; Zhang, X.; Igarashi, Y.; Luo, F. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake. J. Basic Microbiol. 2019, 59, 1112–1124. [Google Scholar] [CrossRef]
- Umetsu, S.; Kanda, M.; Imai, I.; Sakai, R.; Fujita, M.J. Questiomycins, Algicidal Compounds Produced by the Marine Bacterium Alteromonas sp. D and Their Production Cue. Molecules 2019, 24, 4522. [Google Scholar] [CrossRef]
- Schut, F.; Vries, E.D.; Gottschal, J.C.; Robertson, B.R.; Harder, W.; Prins, R.A.; Button, D.K. Isolation of Typical Marine Bacteria by Dilution Culture: Growth, Maintenance, and Characteristics of Isolates under Laboratory Conditions. Appl. Environ. Microbiol. 1993, 59, 2150–2160. [Google Scholar] [CrossRef]
- Yaylacı, E.U. Isolation and characterization of Bacillus spp. from aquaculture cage water and its inhibitory effect against selected Vibrio spp. Arch. Microbiol. 2022, 204, 26. [Google Scholar] [CrossRef]
- Kawser, A.Q.M.R.; Islam, T.; Alam, M.S.; Rahman, M.M.; Salam, M.A. Mechanisms of the Beneficial Effects of Probiotic Bacillus spp. In Aquaculture. In Bacilli in Agrobiotechnology; Springer: Cham, Switzerland, 2022; pp. 453–486. [Google Scholar] [CrossRef]
- Kamilya, D.; Devi, W.M. Bacillus Probiotics and Bioremediation: An Aquaculture Perspective. In Bacilli in Agrobiotechnology; Springer: Cham, Switzerland, 2022; pp. 335–347. [Google Scholar] [CrossRef]
- Nicholson, W.; Munakata, N.; Horneck, G.; Melosh, J.; Setlow, P. Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments. Microbiol. Mol. Biol. Rev. MMBR 2000, 64, 548–572. [Google Scholar] [CrossRef]
- Bedoshvili, Y.; Bayramova, E.; Sudakov, N.; Klimenkov, I.; Kurilkina, M.; Likhoshway, Y.; Zakharova, Y. Impact of Algicidal Bacillus mycoides on Diatom Ulnaria acus from Lake Baikal. Diversity 2021, 13, 469. [Google Scholar] [CrossRef]
- Hou, S.; Shu, W.; Tan, S.; Zhao, L.; Yin, P. Exploration of the Antioxidant System and Photosynthetic System of a Marine Algicidal Bacillus and Its Effect on Four Harmful Algal Bloom Species. Can. J. Microbiol. 2016, 62, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, X.; Lu, J.; Zhou, Y.; Su, J.; Tian, Y.; Zhang, J.; Wang, G.; Zheng, T. A marine bacterium producing protein with algicidal activity against Alexandrium tamarense. Harmful Algae 2012, 13, 83–88. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Wang, Y.; Sun, P.; Ma, S.; Chen, T. Algicidal Effects of a High-Efficiency Algicidal Bacterium Shewanella Y1 on the Toxic Bloom-Causing Dinoflagellate Alexandrium pacificum. Mar. Drugs 2022, 20, 239. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Trainer, V.L.; Scholz, N.L. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat. Toxicol. 2004, 66, 159–170. [Google Scholar] [CrossRef]
- Samson, J.C.; Shumway, S.E.; Weis, J.S. Effects of the toxic dinoflagellate, Alexandrium fundyense on three species of larval fish: A food-chain approach. J. Fish Biol. 2008, 72, 168–188. [Google Scholar] [CrossRef]
- Costa, P.R.; Botelho, M.J.; Lefebvre, K.A. Characterization of paralytic shellfish toxins in seawater and sardines (Sardina pilchardus) during blooms of Gymnodinium catenatum. Hydrobiologia 2010, 655, 89–97. [Google Scholar] [CrossRef]
- Kim, J.D.; Kim, J.Y.; Park, J.K.; Lee, C.G. Selective Control of the Prorocentrum minimum Harmful Algal Blooms by a Novel Algal-Lytic Bacterium Pseudoalteromonas haloplanktis AFMB-008041. Mar. Biotechnol. 2009, 11, 463–472. [Google Scholar] [CrossRef]
- Ko, S.-R.; Jeong, Y.; Cho, S.-H.; Lee, E.; Jeong, B.-S.; Baek, S.H.; Oh, B.-H.; Ahn, C.-Y.; Oh, H.-M.; Cho, B.-K.; et al. Functional role of a novel algicidal compound produced by Pseudoruegeria sp. M32A2M on the harmful algae Alexandrium catenella. Chemosphere 2022, 300, 134535. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.; Xie, S.; Zang, H.; Borriss, R.; Gao, X. Bacilysin from Bacillus amyloliquefaciens FZB42 Has Specific Bactericidal Activity against Harmful Algal Bloom Species. Appl. Environ. Microbiol. 2014, 80, 7512–7520. [Google Scholar] [CrossRef]
- Ahn, C.-Y.; Joung, S.-H.; Jeon, J.-W.; Kim, H.-S.; Yoon, B.-D.; Oh, H.-M. Selective control of cyanobacteria by surfactin-containing culture broth of Bacillus subtilis C1. Biotechnol. Lett. 2003, 25, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-J.; Du, X.-P.; Zhu, J.-M.; Meng, C.-X.; Zhou, J.; Zuo, P. The complete genome sequence of the algicidal bacterium Bacillus subtilis strain JA and the use of quorum sensing to evaluate its antialgal ability. Biotechnol. Rep. 2020, 25, e00421. [Google Scholar] [CrossRef] [PubMed]
- Imai, I. Algicidal ranges in killer bacteria of direct attack type for marine phytoplankton. Bull. Plankton. Soc. Jpn. 1997, 44, 3–9. [Google Scholar]
- Uronen, P.; Kuuppo, P.; Legrand, C.; Tamminen, T. Allelopathic Effects of Toxic Haptophyte Prymnesium parvum Lead to Release of Dissolved Organic Carbon and Increase in Bacterial Biomass. Microb. Ecol. 2007, 54, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.L.; Ten-Hage, L.; Leflaive, J. Allelopathic interactions involving benthic phototrophic microorganisms. Environ. Microbiol. Rep. 2016, 8, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Salehizadeh, H.; Shojaosadati, S.A. Extracellular biopolymeric flocculants: Recent trends and biotechnological importance. Biotechnol. Adv. 2001, 19, 371–385. [Google Scholar] [CrossRef]
- Roth, P.B.; Twiner, M.J.; Mikulski, C.M.; Barnhorst, A.B.; Doucette, G.J. Comparative analysis of two algicidal bacteria active against the red tide dinoflagellate Karenia brevis. Harmful Algae 2008, 7, 682–691. [Google Scholar] [CrossRef]
- Jeong, S.-Y.; Ishida, K.; Ito, Y.; Okada, S.; Murakami, M. Bacillamide, a novel algicide from the marine bacterium, Bacillus sp. SY-1, against the harmful dinoflagellate, Cochlodinium polykrikoides. Tetrahedron Lett. 2003, 44, 8005–8007. [Google Scholar] [CrossRef]
- Xu, Y.; Ge, F.; Wang, N.; Zhu, R.; Tao, N. Selective Algicidal Activity of Surfactant and Its Mechanism. Pract. Period. Hazard. Toxic Radioact. Waste Manag. 2011, 15, 21–25. [Google Scholar] [CrossRef]
- Cho, J.Y. Algicidal activity of marine Alteromonas sp. KNS-16 and isolation of active compounds. J. Agric. Chem. Soc. Jpn. 2012, 76, 1452–1458. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Bravo, I.; Isabel Figueroa, R.; Garcés, E.; Fraga, S.; Massanet, A. The intricacies of dinoflagellate pellicle cysts: The example of Alexandrium minutum cysts from a bloom-recurrent area (Bay of Baiona, NW Spain). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 166–174. [Google Scholar] [CrossRef]
- Haberkorn, H.; Hégaret, H.; Marie, D.; Lambert, C.; Soudant, P. Flow cytometric measurements of cellular responses in a toxic dinoflagellate, Alexandrium minutum, upon exposure to thermal, chemical and mechanical stresses. Harmful Algae 2011, 10, 463–471. [Google Scholar] [CrossRef]
- Jensen, M.Ø.; Moestrup, Ø. Autecology of the toxic dinoflagellate Alexandrium ostenfeldii: Life history and growth at different temperatures and salinities. Eur. J. Phycol. 1997, 32, 9–18. [Google Scholar] [CrossRef]
- Nagasaki, K.; Yamaguchi, M.; Imai, I. Algicidal activity of a killer bacterium against the harmful red tide dinoflagellate Heterocapsa circularisquama isolated from Ago Bay, Japan. Nippon.-Suisan-Gakkaish 2000, 66, 666–673. [Google Scholar] [CrossRef]
- Bell, W.; Mitchell, R. Chemotactic and Growth Responses of Marine Bacteria to Algal Extracellular Products. Biol. Bull. 1972, 143, 265–277. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2015, 80, 91–138. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Su, J.; Tian, Y.; Ning, X.; Hong, H.; Zheng, T. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere. Biol. Control 2010, 52, 123–130. [Google Scholar] [CrossRef]
- Shi, X.; Liu, L.; Li, Y.; Xiao, Y.; Ding, G.; Lin, S.; Chen, J. Isolation of an algicidal bacterium and its effects against the harmful-algal- bloom dinoflagellate Prorocentrum donghaiense (Dinophyceae). Harmful Algae 2018, 80, 72–79. [Google Scholar] [CrossRef]
- Alpermann, T.J.; Tillmann, U.; Beszteri, B.; Cembella, A.D.; John, U. Phenotypic variation and genotypic diversity in a planktonic population of the toxigenic marine dinoflagellate Alexandrium tamarense (Dinophyceae). J. Phycol. 2010, 46, 18–32. [Google Scholar] [CrossRef]
- Soto-Liebe, K.; López-Cortés, X.A.; Fuentes-Valdes, J.J.; Stucken, K.; Gonzalez-Nilo, F.; Vásquez, M. In Silico Analysis of Putative Paralytic Shellfish Poisoning Toxins Export Proteins in Cyanobacteria. PLoS ONE 2013, 8, e55664. [Google Scholar] [CrossRef]
- Jansson, D.; Åstot, C. Analysis of paralytic shellfish toxins, potential chemical threat agents, in food using hydrophilic interaction liquid chromatography–mass spectrometry. J. Chromatogr. A 2015, 1417, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics and physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar] [CrossRef]
- Hwang, D.F.; Lu, Y.H. Influence of environmental and nutritional factors on growth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum. Toxicon 2000, 38, 1491–1503. [Google Scholar] [CrossRef]
- Nam, K.T.; Oh, S.J. Influence of Water Temperature and Salinity on the Production of Paralytic Shellfish Poisoning by Toxic Dinoflagellate Alexandrium catenella (Group I). J. Korean Soc. Mar. Environ. Saf. 2021, 27, 119–126. [Google Scholar] [CrossRef]
- Maas, E.W.; Brooks, H.J.L. Is photosynthesis a requirement for paralytic shellfish toxin production in the dinoflagellate Alexandrium minutum algal–bacterial consortium? J. Appl. Phycol. 2010, 22, 293–296. [Google Scholar] [CrossRef]
- Plumley, F.G.; Wei, Z.; Toivanen, T.B.; Doucette, G.J.; Franca, S. Tn5 mutagenesis of Pseudomonas stutzerisf/ps, a bacterium associated with Alexandrium lusitanicum (dinophyceae) and paralytic shellfish poisoning. J. Phycol. 1999, 35, 1390–1396. [Google Scholar] [CrossRef]
- Baker, T.R.; Doucette, G.J.; Powell, C.L.; Boyer, G.L.; Plumley, F.G. GTX4 imposters: Characterization of fluorescent compounds synthesized by Pseudomonas stutzeri SF/PS and Pseudomonas/Alteromonas PTB-1, symbionts of saxitoxin-producing Alexandrium spp. Toxicon 2003, 41, 339–347. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, S.-G.; Gu, H.-F.; Lai Chan, L.; Hong, H.-S. Paralytic shellfish toxin profiles and toxin variability of the genus Alexandrium (Dinophyceae) isolated from the Southeast China Sea. Toxicon 2006, 48, 138–151. [Google Scholar] [CrossRef]
- Flynn, K.; Franco, J.M.; Fernandez, P.; Reguera, B.; Wood, G.J. Changes in toxin content, biomass and pigments of the dinoflagellate Alexandrium minutum during nitrogen refeeding and growth into nitrogen or phosphorus stress. Mar. Ecol. Prog. Ser. 1994, 111, 99–109. [Google Scholar] [CrossRef]
- Lian, Z.; Li, F.; He, X.; Chen, J.; Yu, R.-C. Rising CO2 will increase toxicity of marine dinoflagellate Alexandrium minutum. J. Hazard. Mater. 2022, 431, 128627. [Google Scholar] [CrossRef] [PubMed]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. Biol. Sci. 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Hansen, P.J. Allelopathic effects of Alexandrium tamarense on other algae: Evidence from mixed growth experiments. Aquat. Microb. Ecol. 2009, 57, 101–112. [Google Scholar] [CrossRef]
- Oshima, Y. Chemical and enzymatic transformation of paralytic shellfish toxins in marine organisms. In Harmful Marine Algal Blooms; Lavoisier: Paris, France, 1995; pp. 475–480. [Google Scholar]
- Lin, H.-P.; Cho, Y.; Yashiro, H.; Yamada, T.; Oshima, Y. Purification and characterization of paralytic shellfish toxin transforming enzyme from Mactra chinensis. Toxicon 2004, 44, 657–668. [Google Scholar] [CrossRef]
- Donovan, C.J.; Ku, J.C.; Quilliam, M.A.; Gill, T.A. Bacterial degradation of paralytic shellfish toxins. Toxicon 2008, 52, 91–100. [Google Scholar] [CrossRef]
- Fast, M.D.; Cembella, A.D.; Ross, N.W. In vitro transformation of paralytic shellfish toxins in the clams Mya arenaria and Protothaca staminea. Harmful Algae 2006, 5, 79–90. [Google Scholar] [CrossRef]
- Smith, E.A.; Mackintosh, F.H.; Grant, F.; Gallacher, S. Sodium channel blocking (SCB) activity and transformation of paralytic shellfish toxins (PST) by dinoflagellate-associated bacteria. Aquat. Microb. Ecol. 2002, 29, 1–9. [Google Scholar] [CrossRef]
- Smith, E.A.; Grant, F.; Ferguson, C.M.J.; Gallacher, S. Biotransformations of Paralytic Shellfish Toxins by Bacteria Isolated from Bivalve molluscs. Appl. Environ. Microbiol. 2001, 67, 2345–2353. [Google Scholar] [CrossRef]
- Tian, H.; Gao, C.; Wang, Z.; Sun, P.; Fan, S.; Zhu, M. Comparative study on in vitro transformation of paralytic shellfish poisoning (PSP) toxins in different shellfish tissues. Acta Oceanol. Sin. 2010, 29, 120–126. [Google Scholar] [CrossRef]
- Andres, J.K.; Yñiguez, A.T.; Maister, J.M.; Turner, A.D.; Olano, D.E.B.; Mendoza, J.; Salvador-Reyes, L.; Azanza, R.V. Paralytic Shellfish Toxin Uptake, Assimilation, Depuration, and Transformation in the Southeast Asian Green-Lipped Mussel (Perna viridis). Toxins 2019, 11, 468. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Pan, L.; Huang, J.J.; Chen, J.; He, X.; Wang, Y.; Wang, J.; Wang, B. Trace determination of multiple hydrophilic cyanotoxins in freshwater by off- and on-line solid phase extraction coupled to liquid chromatography-tandem mass spectrometry. Sci. Total Environ. 2022, 853, 158545. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, S.-F.; Zhang, Y.; Lin, L. Paralytic shellfish toxin biosynthesis in cyanobacteria and dinoflagellates: A molecular overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Abundance (107 Cells/mL) | Shannon | Simpson | |
|---|---|---|---|
| CK-12 | 1.01 ± 0.02 | 2.80 ± 0.38 | 0.73 ± 0.03 |
| A-12 | 2.06 ± 0.11 ** | 3.43 ± 0.41 ** | 0.83 ± 0.03 ** |
| B-12 | 1.46 ± 0.22 * | 3.31 ± 0.39 ** | 0.82 ± 0.03 ** |
| C-12 | 1.30 ± 0.07 | 2.94 ± 0.34 | 0.75 ± 0.03 |
| D-12 | 1.79 ± 0.19 ** | 2.91 ± 0.29 | 0.77 ± 0.02 * |
| PSTs | STX | C1 | C2 | GTX1 | GTX4 | GTX2 | GTX3 | GTX5 | dcGTX2 | dcGTX3 | NEO |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TEF | 1 | 0.006 | 0.096 | 0.99 | 0.73 | 0.36 | 0.64 | 0.06 | 0.833 | 0.724 | 0.801 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, R.; Song, X.; Song, W.; Yu, Z. A New Perspective: Revealing the Algicidal Properties of Bacillus subtilis to Alexandrium pacificum from Bacterial Communities and Toxins. Mar. Drugs 2022, 20, 624. https://doi.org/10.3390/md20100624
Cheng R, Song X, Song W, Yu Z. A New Perspective: Revealing the Algicidal Properties of Bacillus subtilis to Alexandrium pacificum from Bacterial Communities and Toxins. Marine Drugs. 2022; 20(10):624. https://doi.org/10.3390/md20100624
Chicago/Turabian StyleCheng, Ruihong, Xiuxian Song, Weijia Song, and Zhiming Yu. 2022. "A New Perspective: Revealing the Algicidal Properties of Bacillus subtilis to Alexandrium pacificum from Bacterial Communities and Toxins" Marine Drugs 20, no. 10: 624. https://doi.org/10.3390/md20100624
APA StyleCheng, R., Song, X., Song, W., & Yu, Z. (2022). A New Perspective: Revealing the Algicidal Properties of Bacillus subtilis to Alexandrium pacificum from Bacterial Communities and Toxins. Marine Drugs, 20(10), 624. https://doi.org/10.3390/md20100624