


Potential of Epidermal Growth Factor-like Peptide from the Sea Cucumber Stichopus horrens to Increase the Growth of Human Cells: In Silico Molecular Docking Approach

 and
and
Abstract
1. Introduction
2. Results
2.1. Selection of Sh-EGFl-1 Peptide from Contig 498513 Protein Sequence
2.2. Modeling of Sh-EGFl-1 Peptide
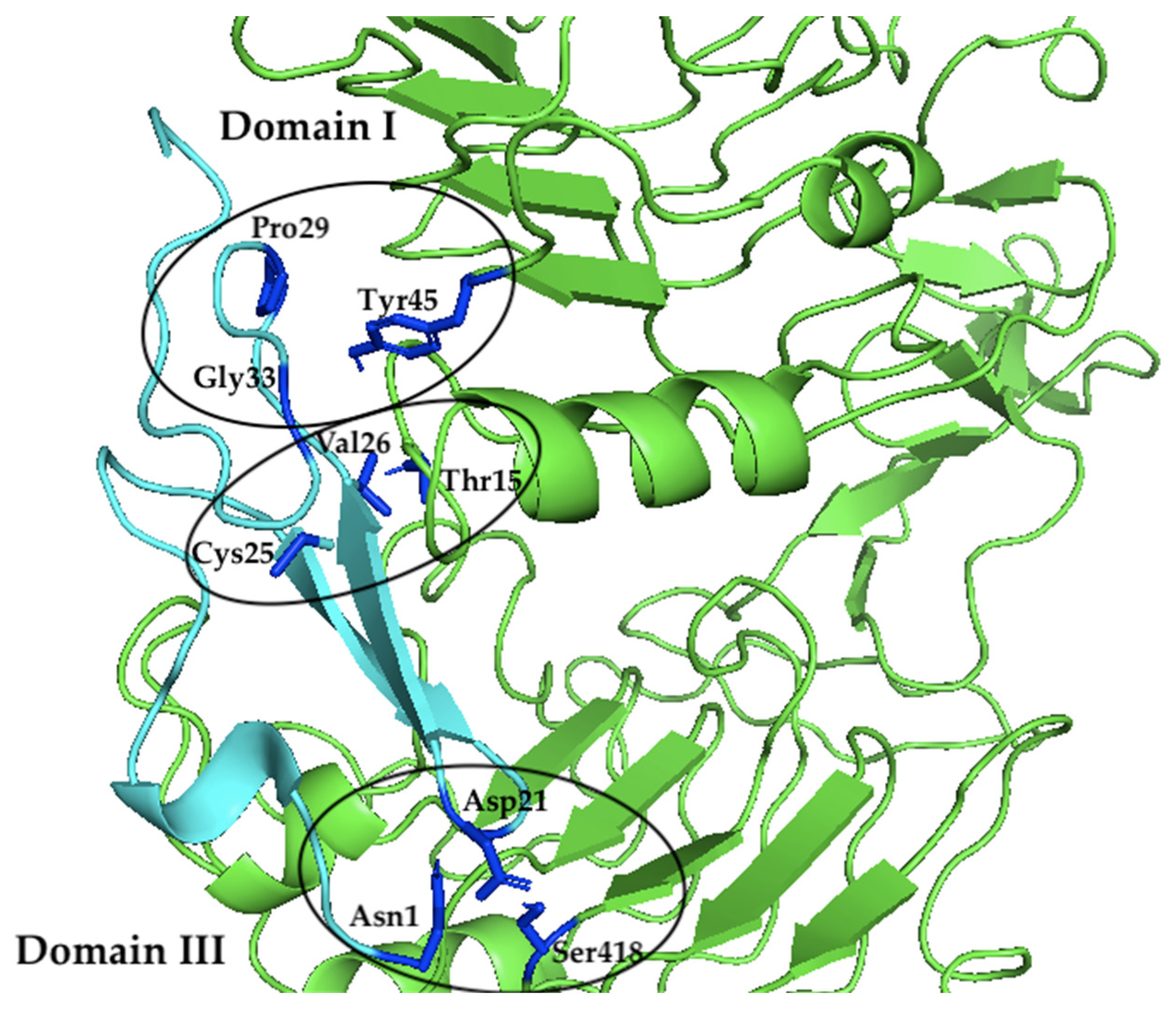
2.3. Molecular Docking Model between Sh-EGFl-1 Peptide and Human EGFR
2.4. Sh-EGFl-1 Peptide–EGFR Binding Assay
2.5. Effect of Sh-EGFl-1 Peptide on Cell Proliferation and Cell Morphology
2.6. Effect of Sh-EGFl-1 on EGF Pathway
3. Discussion
4. Materials and Methods
4.1. S. horrens Protein Modeling and Interaction Study through Molecular Docking
4.2. Sh-EGFl-1 Peptide Modeling and Interaction Study through Molecular Docking
4.3. Preparation of Peptide
4.4. Cell Culture
4.5. Cell Proliferation Assay and Cell Morphology Observation
4.6. Binding Affinity Assay
4.7. Gene Expression Analysis of EGF Pathway
4.8. RNA Extraction
4.9. cDNA Synthesis
4.10. Quantitative Real-Time PCR Containing Array of EGF Pathway Genes
5. Conclusions and Future Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yaacob, H.; Kim, K.; Shahimi, M.; Aziz, N.; Sahil, S. Malaysian sea cucumber (gamat): A prospect in health food and therapeutic. In Proceedings of the Asian Food Technology Seminar, Kuala Lumpur, Malaysia, 6 December 1997. [Google Scholar]
- Patruno, M.; Thorndyke, M.C.; Candia Carnevali, M.D.; Bonasoro, F.; Beesley, P.W. Growth factors, heat-shock proteins and regeneration in echinoderms. J. Exp. Biol. 2001, 204, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, H.; Friedman, K.; Solofa, A.; Mulipola, A.T. A pilot study to investigate the survival of Stichopus horrens after viscera harvest in Samoa. SPC Bêche-de-Mer Inf. Bull. 2007, 26, 2–4. [Google Scholar] [CrossRef]
- Mashanov, V.; García-Arrarás, J. Gut regeneration in holothurians: A snapshot of recent developments. Biol. Bull. 2011, 221, 93–109. [Google Scholar] [CrossRef]
- Siahaan, E.A.; Pangestuti, R.; Munandar, H.; Kim, S.-K. Cosmeceuticals properties of sea cucumbers: Prospects and trends. Cosmetics 2017, 4, 26. [Google Scholar] [CrossRef]
- Aydın, M.; Sevgili, H.; Tufan, B.; Emre, Y.; Köse, S. Proximate composition and fatty acid profile of three different fresh and dried commercial sea cucumbers from Turkey. Int. J. Food Sci. 2011, 46, 500–508. [Google Scholar] [CrossRef]
- Fredalina, B.D.; Ridzwan, B.H.; Abidin, A.A.; Kaswandi, M.A.; Zaiton, H.; Zali, I.; Kittakoop, P.; Jais, A.M. Fatty acid compositions in local sea cucumber, Stichopus chloronotus, for wound healing. Gen. Pharmacol. 1999, 33, 337–340. [Google Scholar] [CrossRef]
- Chen, J. Overview of sea cucumber farming and sea ranching practices in China. SPC Beche-de-Mer Inf. Bull. 2003, 18, 18–23. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea cucumbers metabolites as potent anti-cancer agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef]
- Correia-da-Silva, M.; Sousa, E.; Pinto, M.M.M.; Kijjoa, A. Anticancer and cancer preventive compounds from edible marine organisms. Semin. Cancer Biol. 2017, 46, 55–64. [Google Scholar] [CrossRef]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J. Epidermal growth factor and epidermal growth factor receptor: The yin and yang in the treatment of cutaneous wounds and cancer. Adv. Wound Care 2013, 2, 24–29. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Yang, F.; Li, G. Epidermal growth factor promotes proliferation and migration of follicular outer root sheath cells via Wnt/β-Catenin signaling. Cell Physiol. Biochem. 2016, 39, 360–370. [Google Scholar] [CrossRef]
- Fatimah, S.S.; Tan, G.C.; Chua, K.H.; Tan, A.E.; Hayati, A.R. Effects of epidermal growth factor on the proliferation and cell cycle regulation of cultured human amnion epithelial cells. J. Biosci. Bioeng. 2012, 114, 220–227. [Google Scholar] [CrossRef]
- Hyder, A.; Ehnert, S.; Hinz, H.; Nüssler, A.K.; Fändrich, F.; Ungefroren, H. EGF and HB-EGF enhance the proliferation of programmable cells of monocytic origin (PCMO) through activation of MEK/ERK signaling and improve differentiation of PCMO-derived hepatocyte-like cells. Cell Commun. Signal. 2012, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Bhora, F.; Dunkin, B.; Batzri, S.; Aly, H.; Bass, B.L.; Sidawy, A.; Harmon, J. Effect of growth factors on cell proliferation and epithelialization in human skin. J. Surg. Res. 1995, 59, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Nagate, T.; Matsuda, H. Acceleration of wound healing by gelatin film dressings with epidermal growth factor. J. Vet. Med. Sci. 2005, 67, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Goswami, P. A redox cycle within the cell cycle: Ring in the old with the new. Oncogene 2007, 26, 1101. [Google Scholar] [CrossRef]
- Chiu, J.; Dawes, I.W. Redox control of cell proliferation. Trends Cell Biol. 2012, 22, 592–601. [Google Scholar] [CrossRef]
- Wee, P.; Wang, Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Patar, A.; Jamalullail, S.M.S.S.; Jaafar, H.; Abdullah, J.M. The effect of water extract of sea cucumber Stichopus variegatus on rat spinal astrocytes cell lines. Curr. Neurobiol. 2012, 3, 11–16. [Google Scholar]
- Arundina, I.; Yuliati, Y.; Soesilawati, P.; Damaiyanti, D.W.; Maharani, D. The effects of golden sea cucumber extract (Stichopus hermanii) on the number of lymphocytes during the healing process of traumatic ulcer on wistar rat’s oral mucous. Dent. J. 2015, 48, 100–103. [Google Scholar] [CrossRef]
- Masre, S.F.; Yip, G.W.; Sirajudeen, K.N.; Ghazali, F.C. Wound healing potential of total sulfated glycosaminoglycan (GAG) from Malaysian sea cucumber, Stichopus vastus coelomic fluid. Res. J. Med. Sci. 2015, 9, 67–72. [Google Scholar] [CrossRef]
- Mazliadiyana, M.; Nazrun, A.; Isa, N. Optimum dose of sea cucumber (Stichopus chloronotus) extract for wound healing. Med. Health 2017, 12, 83–89. [Google Scholar] [CrossRef]
- Subramaniam, B.S.; Amuthan, A.; D’Almeida, P.M.; Arunkumar, H. Efficacy of gamat extract in wound healing in albino wistar rats. Int. J. Pharm. Sci. Rev. Res. 2013, 20, 142–145. [Google Scholar]
- Jia, M.; Shi, Z.; Yan, X.; Xu, L.; Dong, L.; Li, J.; Wang, Y.; Yang, S.; Yuan, F. Insulin and heparin-binding epidermal growth factor-like growth factor synergistically promote astrocyte survival and proliferation in serum-free medium. J. Neurosci. Methods 2018, 307, 240–247. [Google Scholar] [CrossRef]
- Rao, Z.; Handford, P.; Mayhew, M.; Knott, V.; Brownlee, G.G.; Stuart, D. The structure of a Ca2+-binding epidermal growth factor-like domain: Its role in protein-protein interactions. Cell 1995, 82, 131–141. [Google Scholar] [CrossRef]
- de Vries, S.J.; Bonvin, A.M. CPORT: A consensus interface predictor and its performance in prediction-driven docking with HADDOCK. PLoS ONE 2011, 6, e17695. [Google Scholar] [CrossRef]
- Ogiso, H.; Ishitani, R.; Nureki, O.; Fukai, S.; Yamanaka, M.; Kim, J.-H.; Saito, K.; Sakamoto, A.; Inoue, M.; Shirouzu, M. Crystal structure of the complex of human epidermal growth factor and receptor extracellular domains. Cell 2002, 110, 775–787. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. Molprobity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- van Zundert, G.C.; Bonvin, A.M. Modeling protein–protein complexes using the HADDOCK webserver “modeling protein complexes with HADDOCK”. Methods Mol. Biol. 2014, 1137, 163–179. [Google Scholar] [CrossRef]
- Panigrahi, S.K. Strong and weak hydrogen bonds in protein-ligand complexes of kinases: A comparative study. Amino Acids 2008, 34, 617–633. [Google Scholar] [CrossRef]
- Bissantz, C.; Kuhn, B.; Stahl, M. A medicinal chemist’s guide to molecular interactions. J. Med. Chem. 2010, 53, 5061–5084. [Google Scholar] [CrossRef]
- Janin, J.; Henrick, K.; Moult, J.; Eyck, L.T.; Sternberg, M.J.; Vajda, S.; Vakser, I.; Wodak, S.J. CAPRI: A critical assessment of predicted interactions. Proteins: Struct. Funct. Genet. 2003, 52, 2–9. [Google Scholar] [CrossRef]
- Tadaki, D.K.; Niyogi, S. The functional importance of hydrophobicity of the tyrosine at position 13 of human epidermal growth factor in receptor binding. J. Biol. Chem. 1993, 268, 10114–10119. [Google Scholar] [CrossRef]
- Kuo, W.-T.; Lin, W.-C.; Chang, K.-C.; Huang, J.-Y.; Yen, K.-C.; Young, I.-C.; Sun, Y.-J.; Lin, F.-H. Quantitative analysis of ligand-EGFR interactions: A platform for screening targeting molecules. PLoS ONE 2015, 10, e0116610. [Google Scholar] [CrossRef]
- Björkelund, H.; Gedda, L.; Andersson, K. Comparing the epidermal growth factor interaction with four different cell lines: Intriguing effects imply strong dependency of cellular context. PLoS ONE 2011, 6, e16536. [Google Scholar] [CrossRef] [PubMed]
- van Zundert, G.C.P.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.S.J.; van Dijk, M.; de Vries, S.J.; Bonvin, A. The HADDOCK2.2 web server: User-friendly integrative modeling of biomolecular complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Das, S.; Stanley, A.; Yadav, L.; Sudhakar, A.; Varma, A.K. Optimized hydrophobic interactions and hydrogen bonding at the target-ligand interface leads the pathways of drug-designing. PLoS ONE 2010, 5, e12029. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Oezguen, N.; Urvil, P.; Ferguson, C.; Dann, S.M.; Savidge, T.C. Regulation of protein-ligand binding affinity by hydrogen bond pairing. Sci. Adv. 2016, 2, e1501240. [Google Scholar] [CrossRef]
- King, I.C.; Catino, J.J. Nonradioactive ligand binding assay for epidermal growth factor receptor. Anal. Biochem. 1990, 188, 97–100. [Google Scholar] [CrossRef]
- Broecker, M.; Hammer, J.; Derwahl, M. Excessive activation of tyrosine kinases leads to inhibition of proliferation in a thyroid carcinoma cell line. Life Sci. 1998, 63, 2373–2386. [Google Scholar] [CrossRef]
- Selander-Sunnerhagen, M.; Ullner, M.; Persson, E.; Teleman, O.; Stenflo, J.; Drakenberg, T. How an epidermal growth factor (EGF)-like domain binds calcium. High resolution NMR structure of the calcium form of the NH2-terminal EGF-like domain in coagulation Factor X. J. Biol. Chem. 1992, 267, 19642–19649. [Google Scholar] [CrossRef]
- Baker, S.J.; Reddy, E.P. CDK4: A key player in the cell cycle, development, and cancer. Genes Cancer 2012, 3, 658–669. [Google Scholar] [CrossRef]
- Terada, Y.; Inoshita, S.; Nakashima, O.; Kuwahara, M.; Sasaki, S.; Marumo, F. Regulation of cyclin D1 expression and cell cycle progression by mitogen-activated protein kinase cascade. Kidney Int. 1999, 56, 1258–1261. [Google Scholar] [CrossRef]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef]
- Blumenberg, M. Profiling and metaanalysis of epidermal keratinocytes responses to epidermal growth factor. BMC Genomics 2013, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- David, M.; Petit, D.; Bertoglio, J. Cell cycle regulation of Rho signaling pathways. Cell Cycle 2012, 11, 3003–3010. [Google Scholar] [CrossRef] [PubMed]
- Soares-Silva, M.; Diniz, F.F.; Gomes, G.N.; Bahia, D. The mitogen-activated protein kinase (MAPK) pathway: Role in immune evasion by trypanosomatids. Front. Microbiol. 2016, 7, 183. [Google Scholar] [CrossRef]
- Ibrahim, N.I.; Wong, S.K.; Mohamed, I.N.; Mohamed, N.; Chin, K.-Y.; Ima-Nirwana, S.; Shuid, A.N. Wound healing properties of selected natural products. Int. J. Environ. Res. 2018, 15, 2360. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H. Targeting the PDGF signaling pathway in tumor treatment. Cell Commun. Signal 2013, 11, 97. [Google Scholar] [CrossRef]
- Keshet, Y.; Seger, R. The MAP kinase signaling cascades: A system of hundreds of components regulates a diverse array of physiological functions. Methods Mol. Biol. 2010, 661, 3–38. [Google Scholar] [CrossRef]
- Xu, F.; Na, L.; Li, Y.; Chen, L. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci. 2020, 10, 54. [Google Scholar] [CrossRef]
- Zeke, A.; Misheva, M.; Remenyi, A.; Bogoyevitch, M.A. JNK signaling: Regulation and functions based on complex protein-protein partnerships. Microbiol. Mol. Biol. Rev. 2016, 80, 793–835. [Google Scholar] [CrossRef]
- Zohdi, R.M.; Zakaria, Z.A.B.; Yusof, N.; Mustapha, N.M.; Abdullah, M.N.H. Sea cucumber (Stichopus hermanii) based hydrogel to treat burn wounds in rats. J. Biomed. Mater. Res. 2011, 98, 30–37. [Google Scholar] [CrossRef]
- Wen, A.P.Y.; Halim, A.S.; Saad, A.Z.M.; Nor, F.M.; Sulaiman, W.A.W. A prospective study evaluating wound healing with sea cucumber gel compared with hydrogel in treatment of skin graft donor sites. Complement. Ther. Med. 2018, 41, 261–266. [Google Scholar] [CrossRef]
- Bryan, P.J.; McClintock, J.B.; Watts, S.A.; Marion, K.R.; Hopkins, T.S. Antimicrobial activity of ethanolic body-wall extracts of echinoderms from the Northern Gulf of Mexico. In Echinoderms through Time, 1st ed.; David, B., Guille, A., Feral, J., Roux, M., Eds.; CRC Press: London, UK, 1994; pp. 17–23. [Google Scholar]
- Bang, F.B.; Lemma, A. Bacterial infection and reaction to injury in some echinoderms. J. Insect Pathol. 1962, 4, 401–414. [Google Scholar]
- Brem, H.; Stojadinovic, O.; Diegelmann, R.F.; Entero, H.; Lee, B.; Pastar, I.; Golinko, M.; Rosenberg, H.; Tomic-Canic, M. Molecular markers in patients with chronic wounds to guide surgical debridement. Mol. Med. 2007, 13, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, M.C. Regeneration in echinoderms: Repair, regrowth, cloning. Inverteb. Surviv. J. 2006, 3, 64–76. [Google Scholar]
- Zhang, X.; Sun, L.; Yuan, J.; Sun, Y.; Gao, Y.; Zhang, L.; Li, S.; Dai, H.; Hamel, J.-F.; Liu, C. The sea cucumber genome provides insights into morphological evolution and visceral regeneration. PLoS Biol. 2017, 15, e2003790. [Google Scholar] [CrossRef]
- Logan, C.Y.; Nusse, R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef]
- Maruyama, I.N. Mechanisms of activation of receptor tyrosine kinases: Monomers or dimers. Cells 2014, 3, 304–330. [Google Scholar] [CrossRef]
- Ferguson, K.M.; Berger, M.B.; Mendrola, J.M.; Cho, H.-S.; Leahy, D.J.; Lemmon, M.A. EGF activates its receptor by removing interactions that autoinhibit ectodomain dimerization. Mol. Cell 2003, 11, 507–517. [Google Scholar] [CrossRef]
- Dawson, J.P.; Berger, M.B.; Lin, C.-C.; Schlessinger, J.; Lemmon, M.A.; Ferguson, K.M. Epidermal growth factor receptor dimerization and activation require ligand-induced conformational changes in the dimer interface. Mol. Cell. Biol. 2005, 25, 7734–7742. [Google Scholar] [CrossRef]
- Macdonald-Obermann, J.L.; Pike, L.J. The intracellular juxtamembrane domain of the epidermal growth factor (EGF) receptor is responsible for the allosteric regulation of EGF binding. J. Biol. Chem. 2009, 284, 13570–13576. [Google Scholar] [CrossRef]
- Miaczynska, M.; Christoforidis, S.; Giner, A.; Shevchenko, A.; Uttenweiler-Joseph, S.; Habermann, B.; Wilm, M.; Parton, R.G.; Zerial, M. APPL proteins link Rab5 to nuclear signal transduction via an endosomal compartment. Cell 2004, 116, 445–456. [Google Scholar] [CrossRef]
- Rocher-Ros, V.; Marco, S.; Mao, J.; Gines, S.; Metzger, D.; Chambon, P.; Balmain, A.; Saura, C. Presenilin modulates EGFR signaling and cell transformation by regulating the ubiquitin ligase Fbw7. Oncogene 2010, 29, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.H.; Furnari, F.B.; Cavenee, W.K.; Bögler, O. Epidermal growth factor receptor signaling intensity determines intracellular protein interactions, ubiquitination, and internalization. Proc. Natl. Acad. Sci. USA 2003, 100, 6505–6510. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Ishii, S.; Richert, N.; Merlino, G.T.; Pastan, I. Epidermal growth factor regulates the expression of its own receptor. Proc. Natl. Acad. Sci. USA 1985, 82, 8374–8378. [Google Scholar] [CrossRef]
- Zhao, X.; Dai, W.; Zhu, H.; Zhang, Y.; Cao, L.; Ye, Q.; Lei, P.; Shen, G. Epidermal growth factor (EGF) induces apoptosis in a transfected cell line expressing EGF receptor on its membrane. Cell Biol. Int. 2006, 30, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, A.; Kornilova, E.; Teslenko, L.; Sorokin, A.; Nikolsky, N. Recycling of epidermal growth factor-receptor complexes in A431 cells. Biochim. Biophys. Acta 1989, 1011, 88–96. [Google Scholar] [CrossRef]
- Yamamoto, T.; Ebisuya, M.; Ashida, F.; Okamoto, K.; Yonehara, S.; Nishida, E. Continuous ERK activation downregulates antiproliferative genes throughout G1 phase to allow cell-cycle progression. Curr. Biol. 2006, 16, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gustafson-Brown, C.; Hanks, S.K.; Nason, K.; Arbeit, J.M.; Pogliano, K.; Wisdom, R.M.; Johnson, R.S. c-Jun is essential for organization of the epidermal leading edge. Dev. Cell 2003, 4, 865–877. [Google Scholar] [CrossRef]
- Andasari, V.; Lü, D.; Swat, M.; Feng, S.; Spill, F.; Chen, L.; Luo, X.; Zaman, M.; Long, M. Computational model of wound healing: EGF secreted by fibroblasts promotes delayed re-epithelialization of epithelial keratinocytes. Integr. Biol. 2018, 10, 605–634. [Google Scholar] [CrossRef]
- Vangone, A.; Bonvin, A.M.J.J. Contacts-based prediction of binding affinity in protein–protein complexes. Elife 2015, 4, e07454. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model 2011, 10, 2778–2786. [Google Scholar] [CrossRef]
- Land, H.; Humble, M.S. YASARA: A tool to obtain structural guidance in biocatalytic investigations. Methods Mol. Biol. 2018, 1685, 43–67. [Google Scholar] [CrossRef] [PubMed]
- Hebert, T.L.; Wu, X.; Yu, G.; Goh, B.C.; Halvorsen, Y.D.C.; Wang, Z.; Moro, C.; Gimble, J.M. Culture effects of epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) on cryopreserved human adipose-derived stromal/stem cell proliferation and adipogenesis. J. Tissue Eng. Regen. Med. 2009, 3, 553–561. [Google Scholar] [CrossRef]
- Kolkmann, A.; Post, M.; Rutjens, M.; van Essen, A.; Moutsatsou, P. Serum-free media for the growth of primary bovine myoblasts. Cytotechnology 2020, 72, 111–120. [Google Scholar] [CrossRef]
- Yamakawa, S.; Hayashida, K. Advances in surgical applications of growth factors for wound healing. Burns Trauma 2019, 7, s41038-019-0148-1. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Hwang, S.R.; Yoon, I.-S. Advanced growth factor delivery systems in wound management and skin regeneration. Molecules 2017, 22, 1259. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, F.; Momeni-Moghaddam, M.; Naderi-Meshkin, H. Regeneration and repair of skin wounds: Various strategies for treatment. Int. J. Low Extrem. Wounds 2019, 18, 247–261. [Google Scholar] [CrossRef]
- Ogino, S.; Morimoto, N.; Sakamoto, M.; Jinno, C.; Sakamoto, Y.; Taira, T.; Suzuki, S. Efficacy of the dual controlled release of HGF and bFGF impregnated with a collagen/gelatin scaffold. J. Surg. Res. 2018, 221, 173–182. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Docking Results | Human EGF-EGFR (1IVO) | Sh-EGFl-1-Human EGFR |
|---|---|---|
| Binding affinity (kcal/mol) | −15.4 | −15.1 |
| Dissociation constant, Kd (M) | 5.2 × 10−12 | 7.8 × 10−12 |
| Number of interacting residues in EGF | 25 | 23 |
| Number of hydrogen bonds | 12 | 10 |
| Number of residues aligned with EGF 1IVO | - | 8 |
| Number of residues aligned with EGFR 1IVO | - | 14 |
| RMSD (Å) (<3 Å) | - | 0.852 |
| Ligand-RMSD (Å) (<10 Å) | - | 7.873 |
| 10 nM Sh-EGFl-1 | 10 nM rhEGF | Negative Control | |||
|---|---|---|---|---|---|
| MAPK signaling | |||||
| CBL | − | (1.00) | − | (0.96) | − |
| SHC1 | + | (0.94) | − | (0.80) | + |
| HRAS | + | (0.93) | − | (0.62) | + |
| RASA1 (p120GAP) | + | (1.07) | − | (0.83) | + |
| RAP1A | + | (1.14) | − | (0.80) | unc. |
| ATF1 | + | (0.04) | − | (0.87) | + |
| ATF2 | + | (0.89) | − | (0.67) | + |
| NFATC3 | + | (1.22) | − | (0.96) | − |
| CASP3 | + | (1.05) | − | (0.70) | + |
| CASP9 | + | (0.93) | − | (0.63) | + |
| MAP2K1 (MEK1) | unc. | (0.91) | − | (0.74) | + |
| MAP2K4 (MKK4) | unc. | (0.84) | − | (0.51) | + |
| MAP2K7 (MKK7) | + | (1.0) | + | (0.95) | + |
| MAP3K2 (MEKK2) | + | (1.27) | − | (0.82) | − |
| MAPK1 (ERK2) | + | (1.03) | − | (0.76) | − |
| MAPK10 | − | (0.55) | + | (1.24) | + |
| MAPK3 (ERK1) | + | (0.98) | − | (0.79) | + |
| MAPK8 | + | (0.99) | − | (0.73) | + |
| MAPK9 | unc. | (0.80) | − | (0.54) | + |
| MKNK1 | − | (0.82) | − | (0.54) | + |
| RPS6KA5 (p70S6K) | + | (0.97) | unc. | (0.75) | + |
| RPS6KB1 (p70S6K) | + | (1.00) | − | (0.68) | + |
| TP53 | + | (0.96) | unc. | (0.82) | + |
| DUSP1 | + | (0.97) | − | (0.50) | + |
| DUSP6 | + | (1.05) | − | (0.74) | + |
| IKBKB | + | (1.05) | − | (0.73) | + |
| PI3K signaling | |||||
| SHC1 | + | (0.94) | − | (0.78) | + |
| GRB2 | + | (1.07) | − | (0.76) | + |
| GAB1 | + | (1.00) | + | (0.88) | + |
| P13KCA | + | (1.47) | − | (1.08) | − |
| PIK3R1 | + | (0.98) | − | (0.80) | + |
| PIK3R2 | + | (1.17) | − | (0.75) | − |
| E1F4E | + | (1.28) | + | (1.24) | − |
| PDPK1 (PDK1) | unc. | (0.95) | − | (0.86) | + |
| AKT1 | + | (0.95) | − | (0.69) | + |
| AKT3 | + | (1.11) | − | (0.87) | unc. |
| TP53 | + | (0.96) | unc. | (0.82) | + |
| IKBKB (IKK) | + | (1.06) | − | (0.73) | + |
| GSK3A | unc. | (0.87) | − | (0.75) | − |
| GSK3B | + | (1.06) | − | (0.84) | unc. |
| RPS6KA5 (p70S6K) | + | (0.97) | unc. | (0.75) | − |
| RPS6KB1 (p70S6K) | + | (1.00) | − | (0.68) | − |
| NFKB | + | (1.02) | − | (0.90) | − |
| CCND1 | − | (1.17) | − | (0.83) | − |
| JAK-STAT signaling | |||||
| SRC | + | (1.18) | + | (1.16) | + |
| STAT3 | + | (1.06) | − | (0.89) | unc. |
| STAT5 | + | (1.06) | − | (0.89) | − |
| JAK1 | + | (1.02) | − | (0.77) | − |
| FOS | + | (1.50) | − | (1.02) | − |
| JUN | + | (1.32) | unc. | (0.98) | unc. |
| ELK1 | + | (1.08) | − | (0.77) | − |
| PLC gamma signaling | |||||
| PLCG1 (PLC) | + | (1.18) | − | (0.89) | − |
| PPP2CA (TSC2) | + | (0.94) | − | (0.69) | − |
| PRKCA | + | (1.10) | − | (0.78) | unc. |
| RPS6KA5 (p70S6K) | + | (0.97) | unc. | (0.75) | − |
| RPS6KB1 (p70S6K) | + | (1.00) | − | (0.68) | − |
| IKBKB (IKK) | + | (1.06) | − | (0.73) | − |
| NFKB | + | (1.02) | − | (0.90) | − |
| Rho signaling | |||||
| RHOA | + | (1.08) | − | (0.84) | unc. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilus, N.S.M.; Muhamad, A.; Shahidan, M.A.; Yusof, N.Y.M. Potential of Epidermal Growth Factor-like Peptide from the Sea Cucumber Stichopus horrens to Increase the Growth of Human Cells: In Silico Molecular Docking Approach. Mar. Drugs 2022, 20, 596. https://doi.org/10.3390/md20100596
Pilus NSM, Muhamad A, Shahidan MA, Yusof NYM. Potential of Epidermal Growth Factor-like Peptide from the Sea Cucumber Stichopus horrens to Increase the Growth of Human Cells: In Silico Molecular Docking Approach. Marine Drugs. 2022; 20(10):596. https://doi.org/10.3390/md20100596
Chicago/Turabian StylePilus, Nur Shazwani Mohd, Azira Muhamad, Muhammad Ashraf Shahidan, and Nurul Yuziana Mohd Yusof. 2022. "Potential of Epidermal Growth Factor-like Peptide from the Sea Cucumber Stichopus horrens to Increase the Growth of Human Cells: In Silico Molecular Docking Approach" Marine Drugs 20, no. 10: 596. https://doi.org/10.3390/md20100596
APA StylePilus, N. S. M., Muhamad, A., Shahidan, M. A., & Yusof, N. Y. M. (2022). Potential of Epidermal Growth Factor-like Peptide from the Sea Cucumber Stichopus horrens to Increase the Growth of Human Cells: In Silico Molecular Docking Approach. Marine Drugs, 20(10), 596. https://doi.org/10.3390/md20100596