
Unveiling the Chemical Diversity of the Deep-Sea Sponge Characella pachastrelloides

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
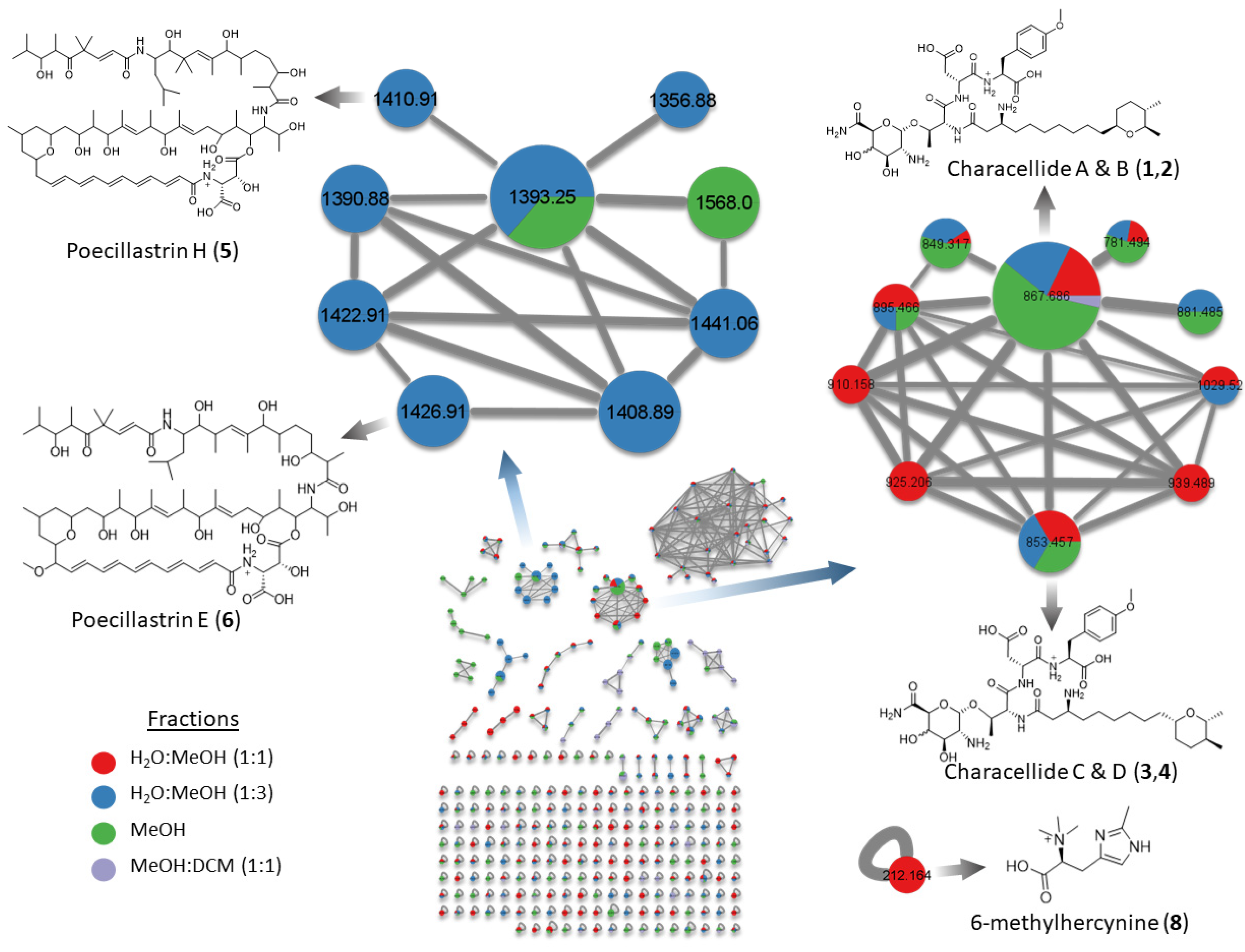
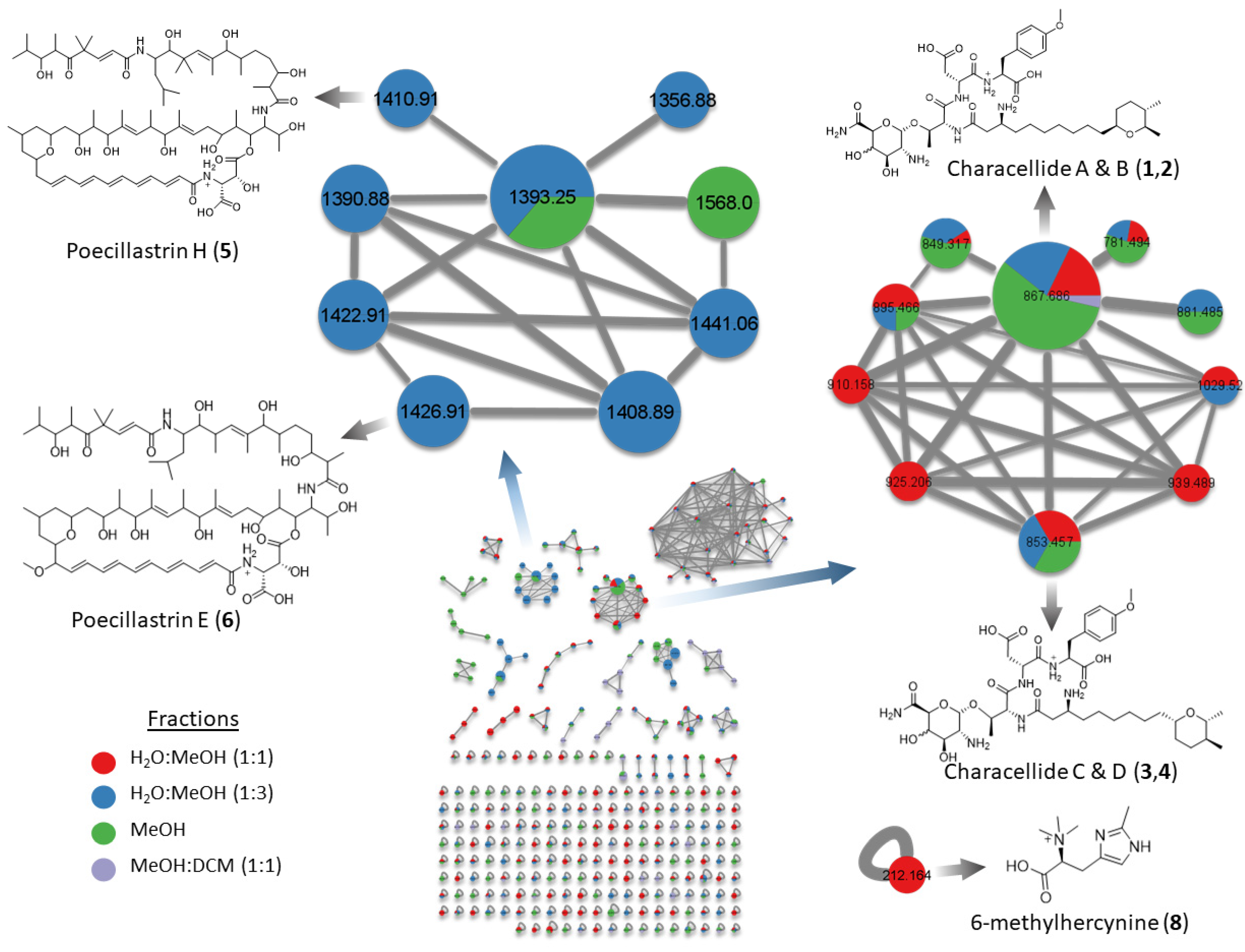
2.1. Molecular Networking
2.2. Glycolipopeptides: Characellides
2.3. Polyketides: Poecillastrins
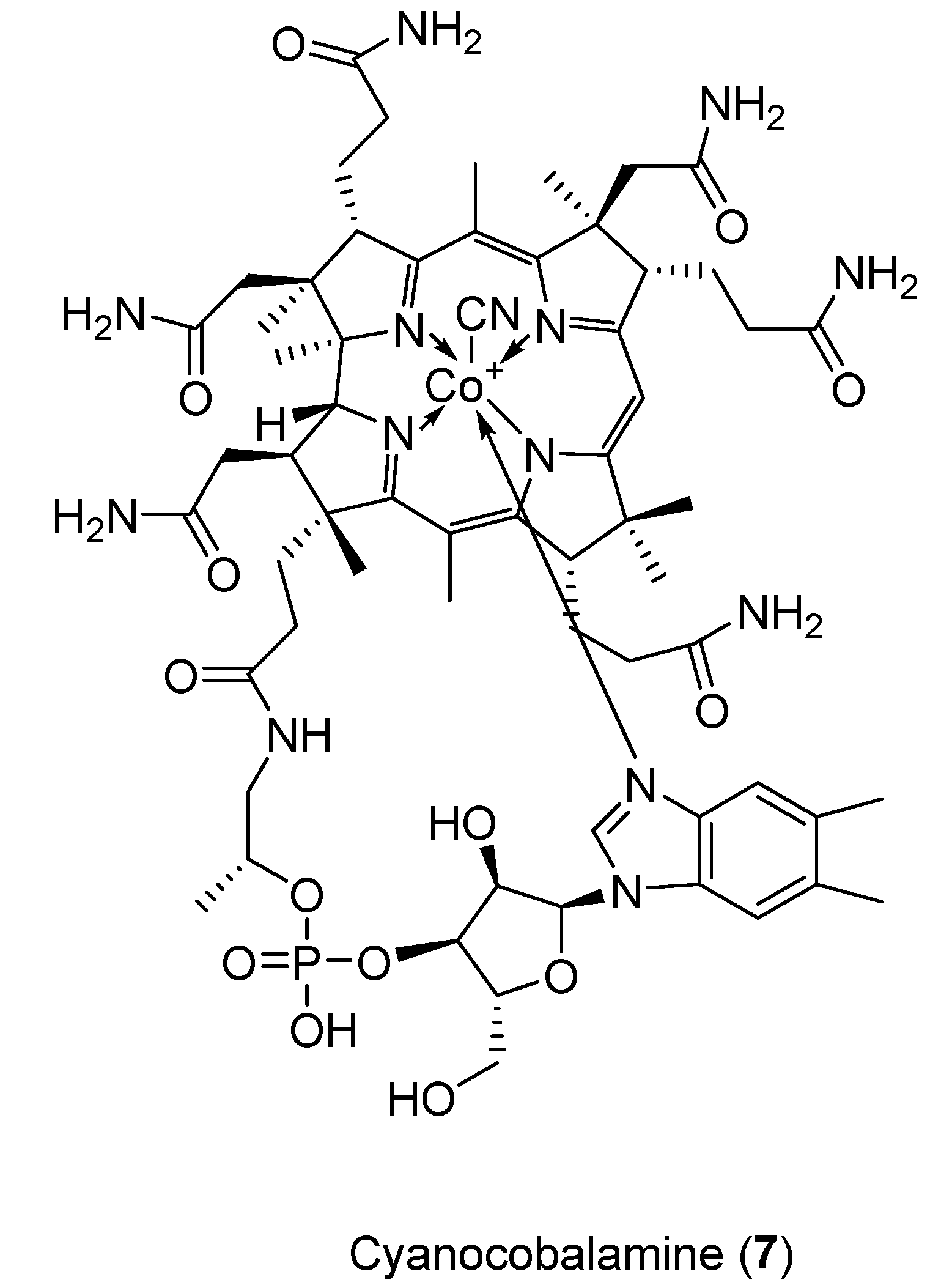
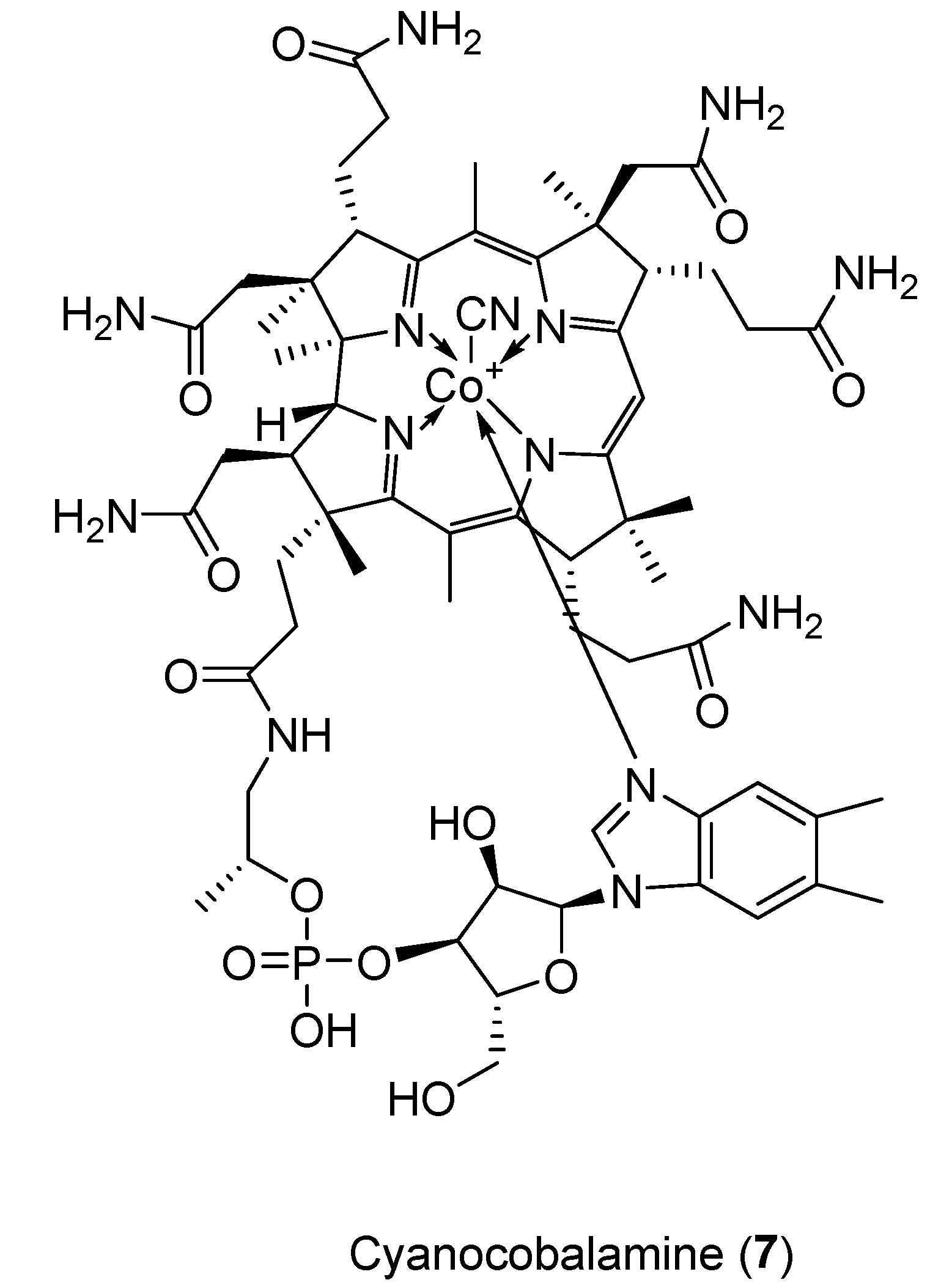
2.4. Cyanocobalamin
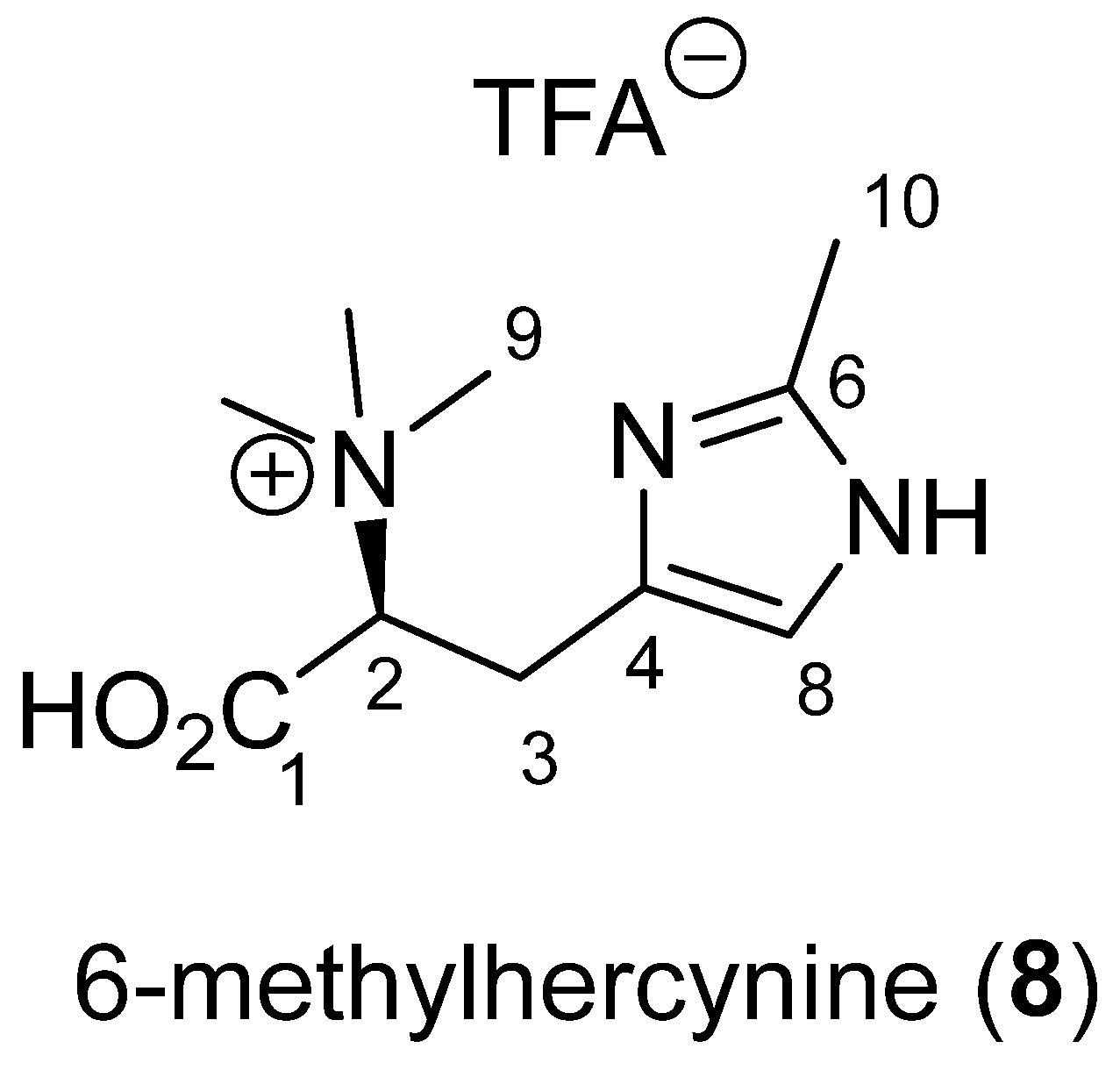
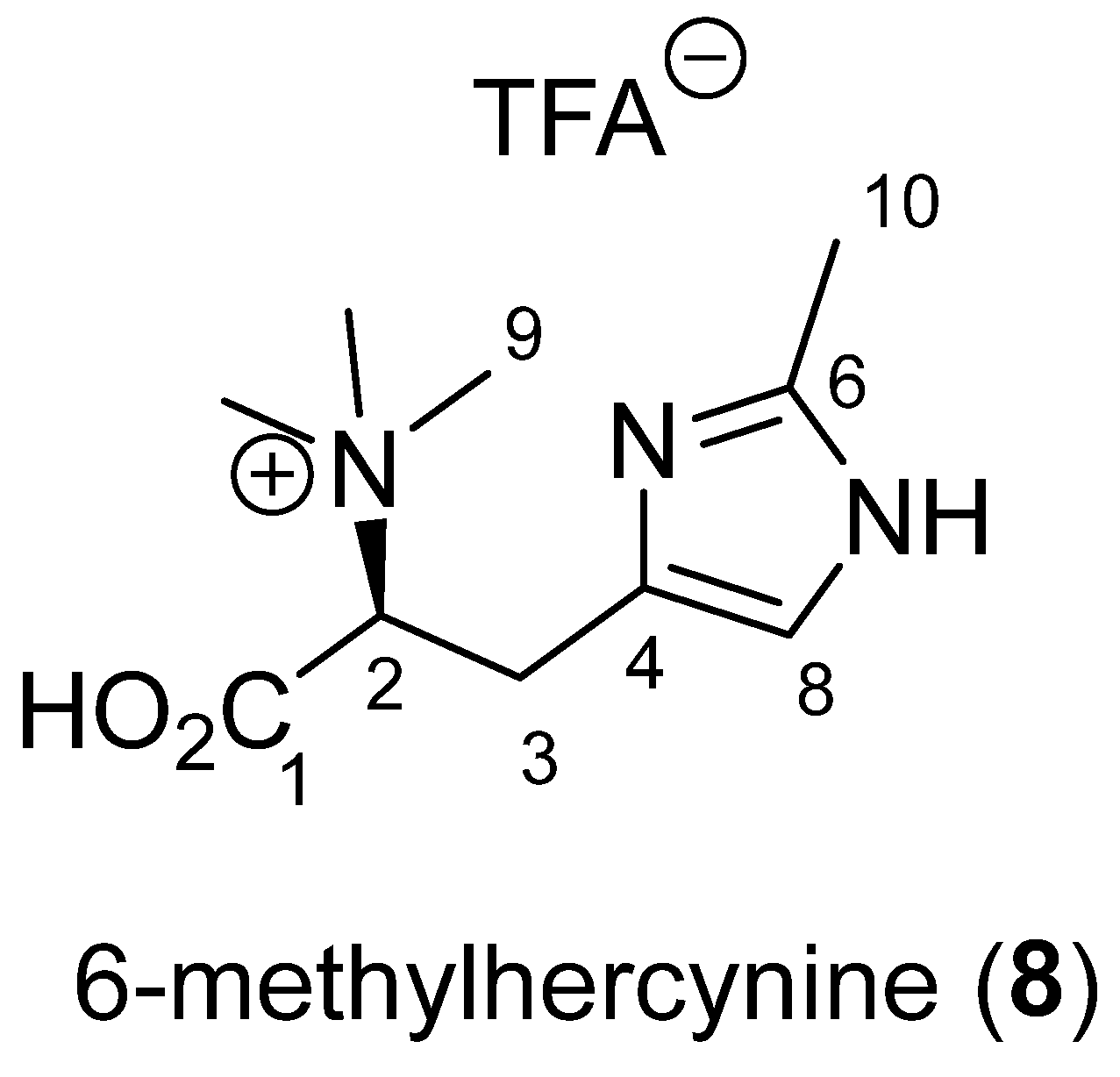
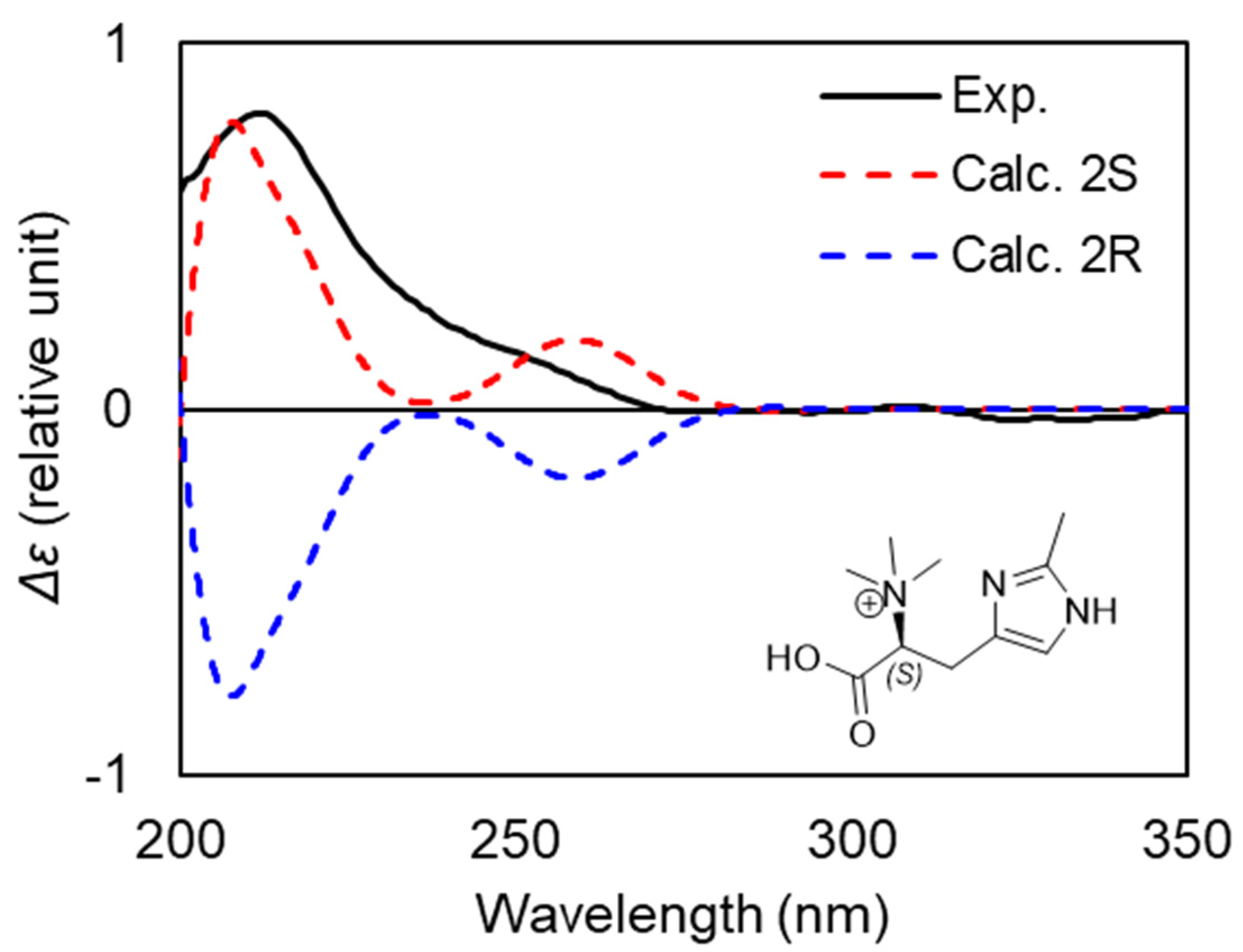
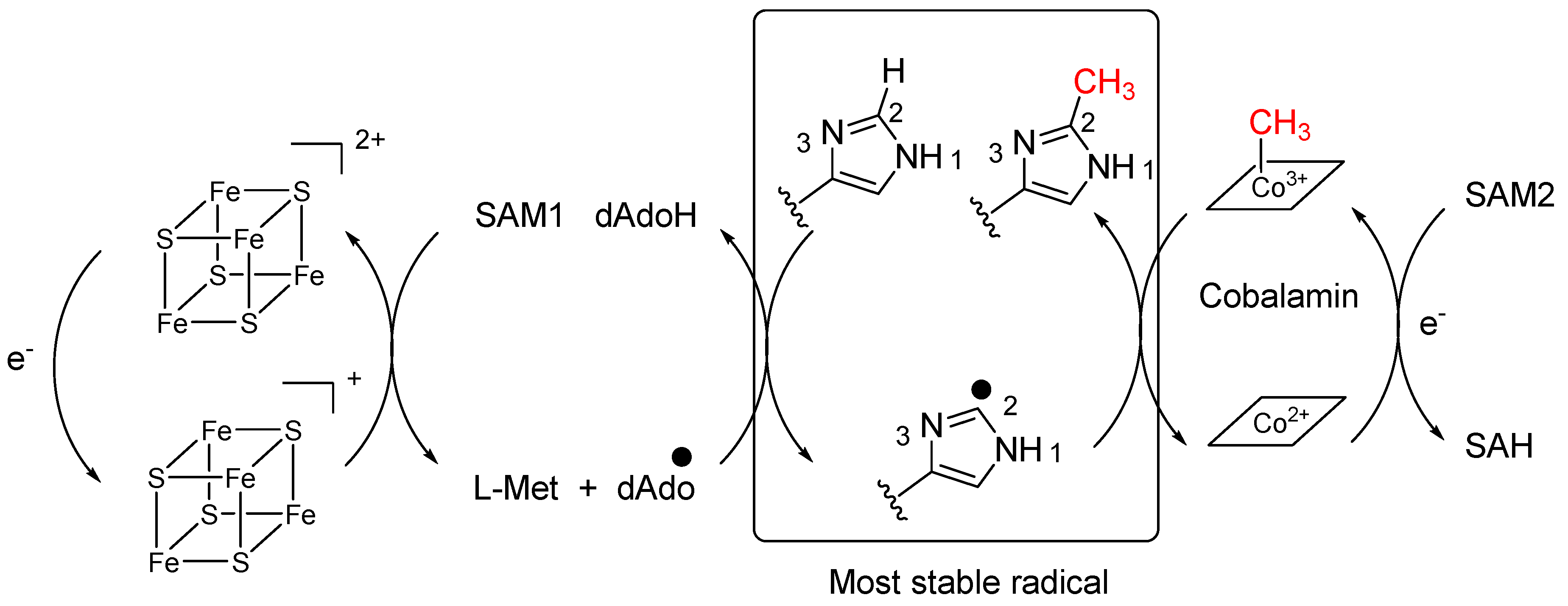
2.5. Betaine: 6-Methylhercynine
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Extraction and Isolation
3.4. Computational Methods
3.5. Molecular Network
3.6. Biological Activities
Clonogenic Survival Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, M.P.; White, M.; Wilson, A.; Würzberg, L.; Schwabe, E.; Folch, H.; Allcock, A.L. A Vertical Wall Dominated by Acesta excavata and Neopycnodonte zibrowii, Part of an Undersampled Group of Deep-Sea Habitats. PLoS ONE 2013, 8, e79917. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Arcaya, U.; Ramirez-Llodra, E.; Aguzzi, J.; Allcock, A.L.; Davies, J.S.; Dissanayake, A.; Harris, P.; Howell, K.; Huvenne, V.A.I.; Macmillan-Lawler, M.; et al. Ecological Role of Submarine Canyons and Need for Canyon Conservation: A Review. Front. Mar. Sci. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Pilkington, L.I. A Chemometric Analysis of Deep-Sea Natural Products. Molecules 2019, 24, 3942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, M.C.; Puga, J.; Serôdio, J.; Gomes, N.C.M.; Calado, R. Trends in the Discovery of New Marine Natural Products from Invertebrates over the Last Two Decades—Where and What Are We Bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef] [PubMed]
- Genta-Jouve, G.; Thomas, O.P. Chapter four—Sponge Chemical Diversity: From Biosynthetic Pathways to Ecological Roles. In Advances in Marine Biology; Becerro, M.A., Uriz, M.J., Maldonado, M., Turon, X., Eds.; Academic Press: Cambridge, MA, USA, 2012; Volume 62, pp. 183–230. [Google Scholar]
- Wright, P.C.; Westacott, R.E.; Burja, A.M. Piezotolerance as a metabolic engineering tool for the biosynthesis of natural products. Biomol. Eng. 2003, 20, 325–331. [Google Scholar] [CrossRef]
- Marinlit. Royal Society of Chemistry. Available online: https://marinlit.rsc.org/ (accessed on 12 July 2021).
- Afoullouss, S.; Calabro, K.; Genta-Jouve, G.; Gegunde, S.; Alfonso, A.; Nesbitt, R.; Morrow, C.; Alonso, E.; Botana, L.M.; Allcock, A.L.; et al. Treasures from the Deep: Characellides as Anti-Inflammatory Lipoglycotripeptides from the Sponge Characella pachastrelloides. Org. Lett. 2019, 21, 246–251. [Google Scholar] [CrossRef]
- Marcella, S.; Afoullouss, S.; Thomas, O.P.; Allcock, A.L.; Murphy, P.V.; Loffredo, S. Immunomodulatory properties of characellide A on human peripheral blood mononuclear cells. Inflammopharmacology 2021, 29, 1201–1210. [Google Scholar] [CrossRef]
- Ragini, K.; Piggott, A.M.; Karuso, P. Bisindole Alkaloids from a New Zealand Deep-Sea Marine Sponge Lamellomorpha strongylata. Mar. Drugs 2019, 17, 683. [Google Scholar] [CrossRef] [Green Version]
- Nakamukai, S.; Takada, K.; Furihata, K.; Ise, Y.; Okada, S.; Morii, Y.; Yamawaki, N.; Takatani, T.; Arakawa, O.; Gustafson, K.R.; et al. Stellatolide H, a cytotoxic peptide lactone from a deep-sea sponge Discodermia sp. Tetrahedron Lett. 2018, 59, 2532–2536. [Google Scholar] [CrossRef]
- Irie, R.; Hitora, Y.; Ise, Y.; Okada, S.; Takada, K.; Matsunaga, S. Poecillastrin E, F, and G, cytotoxic chondropsin-type macrolides from a marine sponge Poecillastra sp. Tetrahedron 2018, 74, 1430–1434. [Google Scholar] [CrossRef]
- Wright, A.E.; Roberts, J.C.; Guzmán, E.A.; Pitts, T.P.; Pomponi, S.A.; Reed, J.K. Analogues of the Potent Antitumor Compound Leiodermatolide from a Deep-Water Sponge of the Genus Leiodermatium. J. Nat. Prod. 2017, 80, 735–739. [Google Scholar] [CrossRef]
- Dumdei, E.J.; Blunt, J.W.; Munro, M.H.G.; Pannell, L.K. Isolation of Calyculins, Calyculinamides, and Swinholide H from the New Zealand Deep-Water Marine Sponge Lamellomorpha strongylata. J. Org. Chem. 1997, 62, 2636–2639. [Google Scholar] [CrossRef] [PubMed]
- Calabro, K.; Kalahroodi, E.L.; Rodrigues, D.; Díaz, C.; Cruz, M.D.L.; Cautain, B.; Laville, R.; Reyes, F.; Pérez, T.; Soussi, B.; et al. Poecillastrosides, Steroidal Saponins from the Mediterranean Deep-Sea Sponge Poecillastra compressa (Bowerbank, 1866). Mar. Drugs 2017, 15, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanashi, E.; Takada, K.; Hashimoto, M.; Itoh, Y.; Ise, Y.; Ohtsuka, S.; Okada, S.; Matsunaga, S. Cytotoxic linear acetylenes from a marine sponge Pleroma sp. Tetrahedron 2015, 71, 9564–9570. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Z.; Wang, Z.; Liu, X.; Jiang, Y.; Jiao, X.; Xie, P. Total Synthesis of the Proposed Structure of Characellide B. Org. Lett. 2021, 23, 3680–3684. [Google Scholar] [CrossRef]
- Suo, R.; Takada, K.; Irie, R.; Watanabe, R.; Suzuki, T.; Ise, Y.; Ohtsuka, S.; Okada, S.; Matsunaga, S. Poecillastrin H, a Chondropsin-Type Macrolide with a Conjugated Pentaene Moiety, from a Characella sp. Marine Sponge. J. Nat. Prod. 2018, 81, 1295–1299. [Google Scholar] [CrossRef]
- Bowman, E.J.; Gustafson, K.R.; Bowman, B.J.; Boyd, M.R. Identification of a New Chondropsin Class of Antitumor Compound That Selectively Inhibits V-ATPases*. J. Biol. Chem. 2003, 278, 44147–44152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B12: A review and future perspectives. Microb. Cell. Fact. 2017, 16, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doxey, A.C.; Kurtz, D.A.; Lynch, M.D.J.; Sauder, L.A.; Neufeld, J.D. Aquatic metagenomes implicate Thaumarchaeota in global cobalamin production. ISME J. 2015, 9, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Pulte, A.; Wagner, S.; Kogler, H.; Spiteller, P. Pelianthinarubins A and B, Red Pyrroloquinoline Alkaloids from the Fruiting Bodies of the Mushroom Mycena pelianthina. J. Nat. Prod. 2016, 79, 873–878. [Google Scholar] [CrossRef]
- Liu, D.; Tan, H.-Q.; Chen, W.-L.; Li, Y.-G.; Wang, E.-B. Resolution of chiral polyoxoanion [P2Mo18O62]6− with histidine. CrystEngComm 2010, 12, 2044–2046. [Google Scholar] [CrossRef]
- Rienzo, M.; Lummis, S.C.R.; Dougherty, D.A. Structural Requirements in the Transmembrane Domain of GLIC Revealed by Incorporation of Noncanonical Histidine Analogs. Chem. Biol. 2014, 21, 1700–1706. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Cohen, L.A.; El-Kadi, N.A.; King, M.M. Regiospecific alkylation of histidine and histamine at C-2. Tetrahedron 1997, 53, 2365–2370. [Google Scholar] [CrossRef]
- Stampfli, A.R.; Blankenfeldt, W.; Seebeck, F.P. Structural basis of ergothioneine biosynthesis. Curr. Opin. Struct. Biol. 2020, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; van der Donk, W.A.; Liu, W. Radical-Mediated Enzymatic Methylation: A Tale of Two SAMS. Acc. Chem. Res. 2012, 45, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Alkhalaf, L.M.; de los Santos, E.L.C.; Challis, G.L. Mechanistic insights into class B radical-S-adenosylmethionine methylases: Ubiquitous tailoring enzymes in natural product biosynthesis. Curr. Opin. Chem. Biol. 2016, 35, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, P.H.; Jansma, M.J.; Hoye, T.R. A guide to small-molecule structure assignment through computation of (1H and 13C) NMR chemical shifts. Nat. Protoc. 2014, 9, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16 Rev. C.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Tomasi, J.; Mennucci, B.; Cammi, R. Quantum Mechanical Continuum Solvation Models. Chem. Rev. 2005, 105, 2999–3094. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, T.; Schaumlöffel, A.; Hemberger, Y.; Bringmann, G. SpecDis: Quantifying the Comparison of Calculated and Experimental Electronic Circular Dichroism Spectra. Chirality 2013, 25, 243–249. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 6-Methylhercynine (8) | |
|---|---|---|
| δH, Mult. (J in Hz) | δC | |
| 1 | - | 169.5 |
| 2 | 3.98, dd (12.0, 3.5) | 76.5 |
| 3 | 3.46, dd (14.0, 3.5) | 22.6 |
| 3.36, dd (14.0, 12.0) | ||
| 4 | - | 127.7 |
| 6 | - | 144.0 |
| 8 | 7.29, s | 118.8 |
| 9 | 3.32, s | 52.2 |
| 10 | 2.73, s | 15.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afoullouss, S.; Sanchez, A.R.; Jennings, L.K.; Kee, Y.; Allcock, A.L.; Thomas, O.P. Unveiling the Chemical Diversity of the Deep-Sea Sponge Characella pachastrelloides. Mar. Drugs 2022, 20, 52. https://doi.org/10.3390/md20010052
Afoullouss S, Sanchez AR, Jennings LK, Kee Y, Allcock AL, Thomas OP. Unveiling the Chemical Diversity of the Deep-Sea Sponge Characella pachastrelloides. Marine Drugs. 2022; 20(1):52. https://doi.org/10.3390/md20010052
Chicago/Turabian StyleAfoullouss, Sam, Anthony R. Sanchez, Laurence K. Jennings, Younghoon Kee, A. Louise Allcock, and Olivier P. Thomas. 2022. "Unveiling the Chemical Diversity of the Deep-Sea Sponge Characella pachastrelloides" Marine Drugs 20, no. 1: 52. https://doi.org/10.3390/md20010052
APA StyleAfoullouss, S., Sanchez, A. R., Jennings, L. K., Kee, Y., Allcock, A. L., & Thomas, O. P. (2022). Unveiling the Chemical Diversity of the Deep-Sea Sponge Characella pachastrelloides. Marine Drugs, 20(1), 52. https://doi.org/10.3390/md20010052