
Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates

 , and
, and 
Abstract
1. Introduction
2. Results
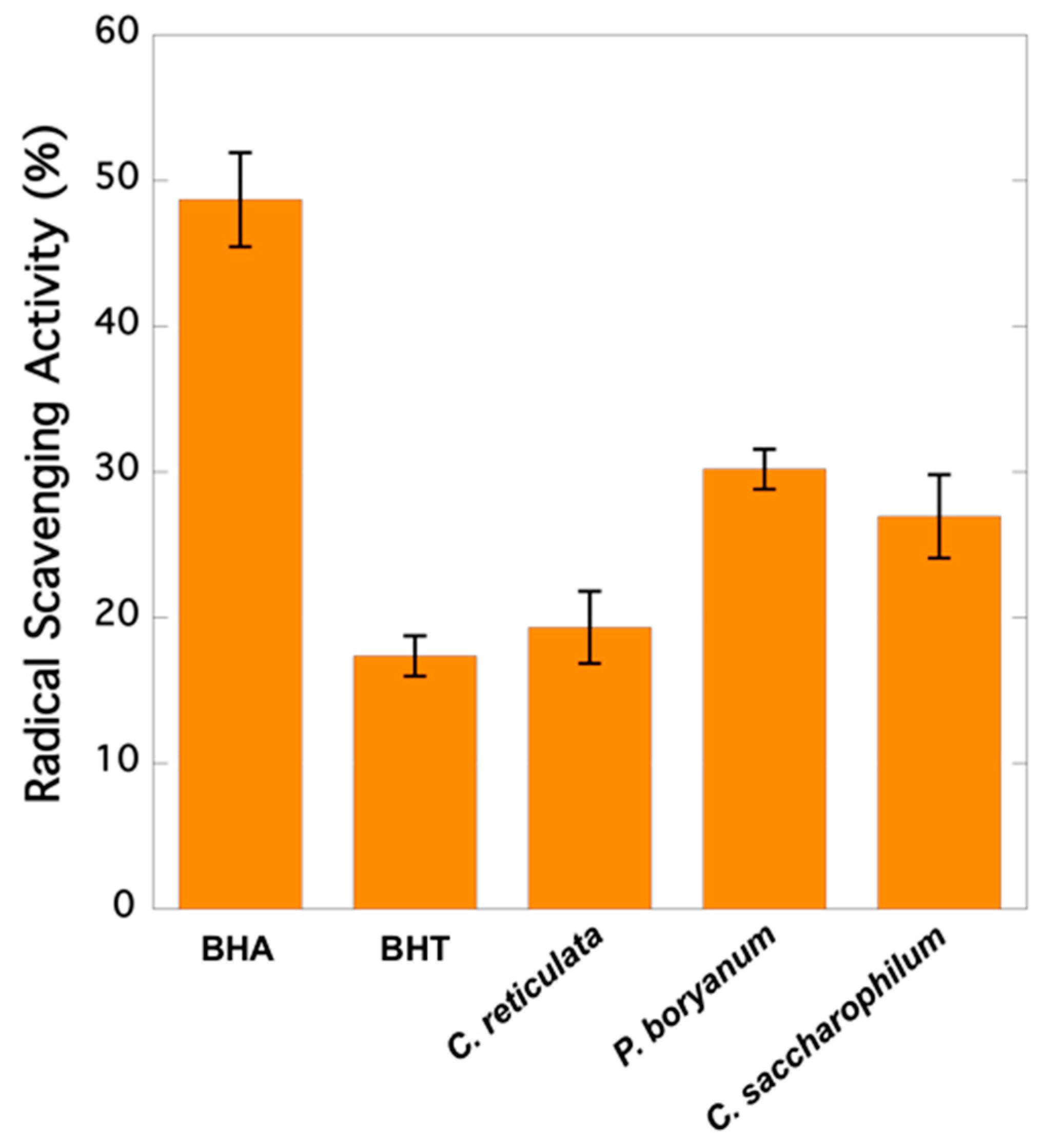
2.1. Radical Scavenging Activity
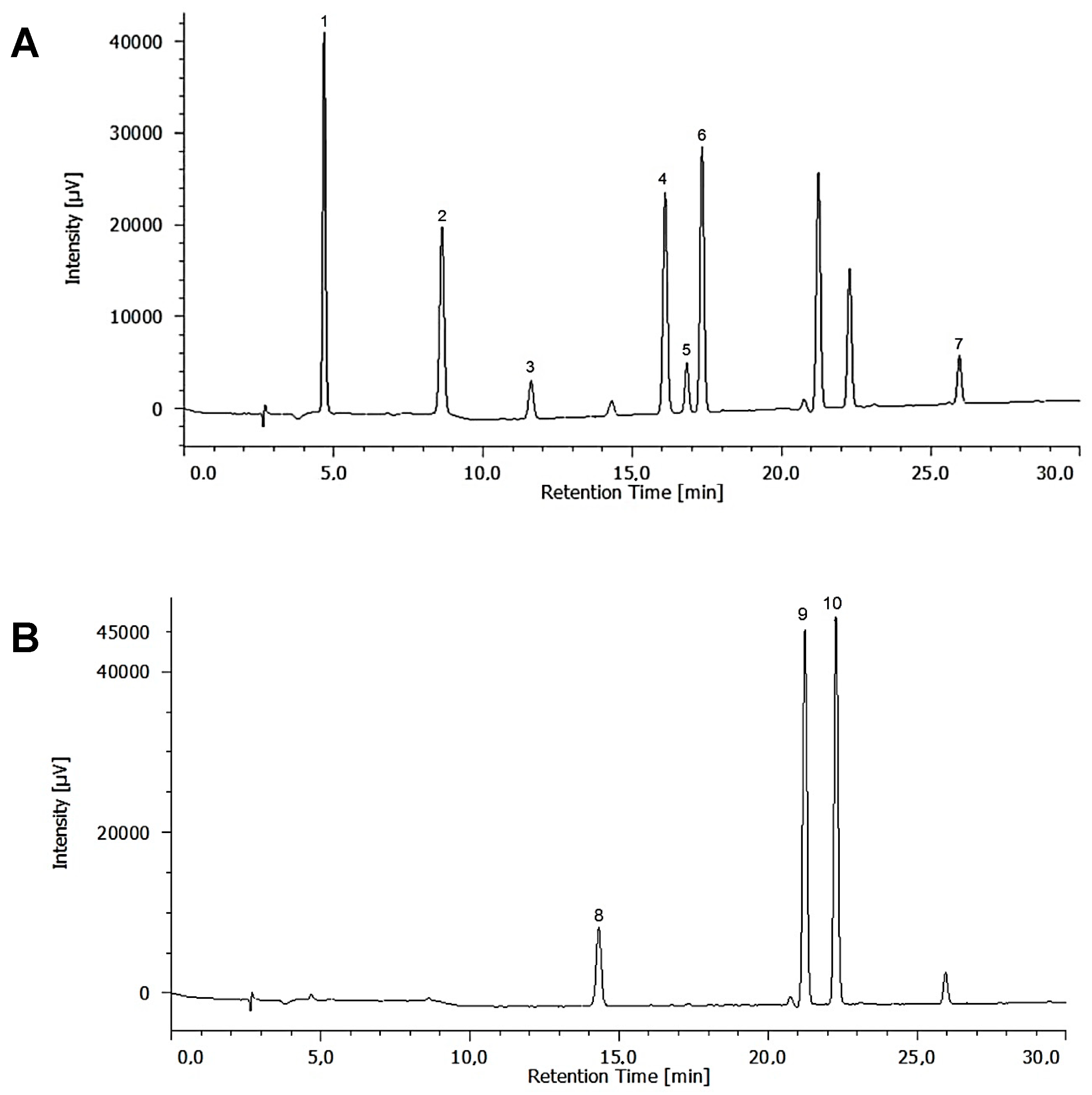
2.2. Identification and Quantification of Phenolic Compounds
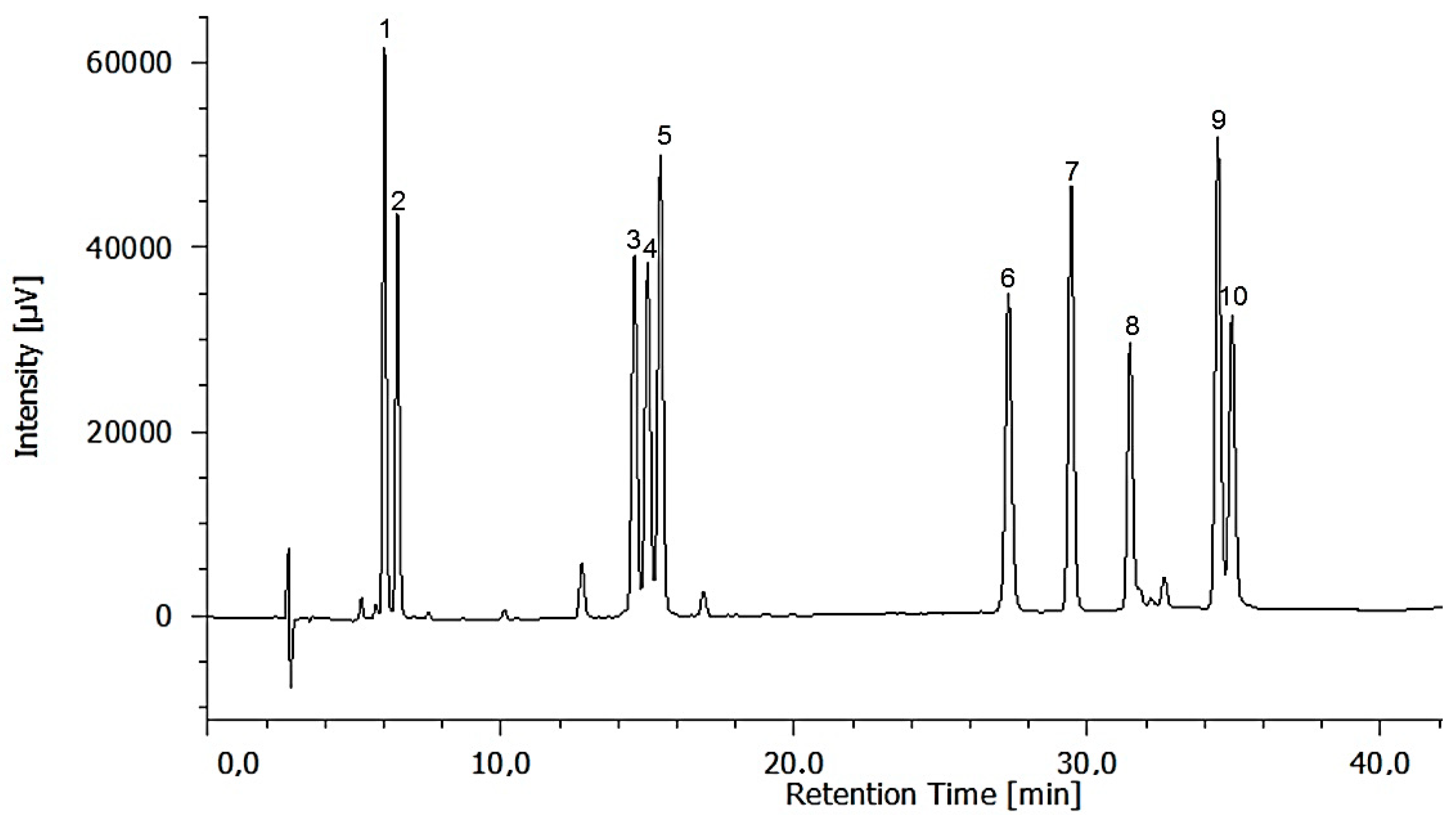
2.3. Identification and Quantification of Free and Total Analysed Amino Acids
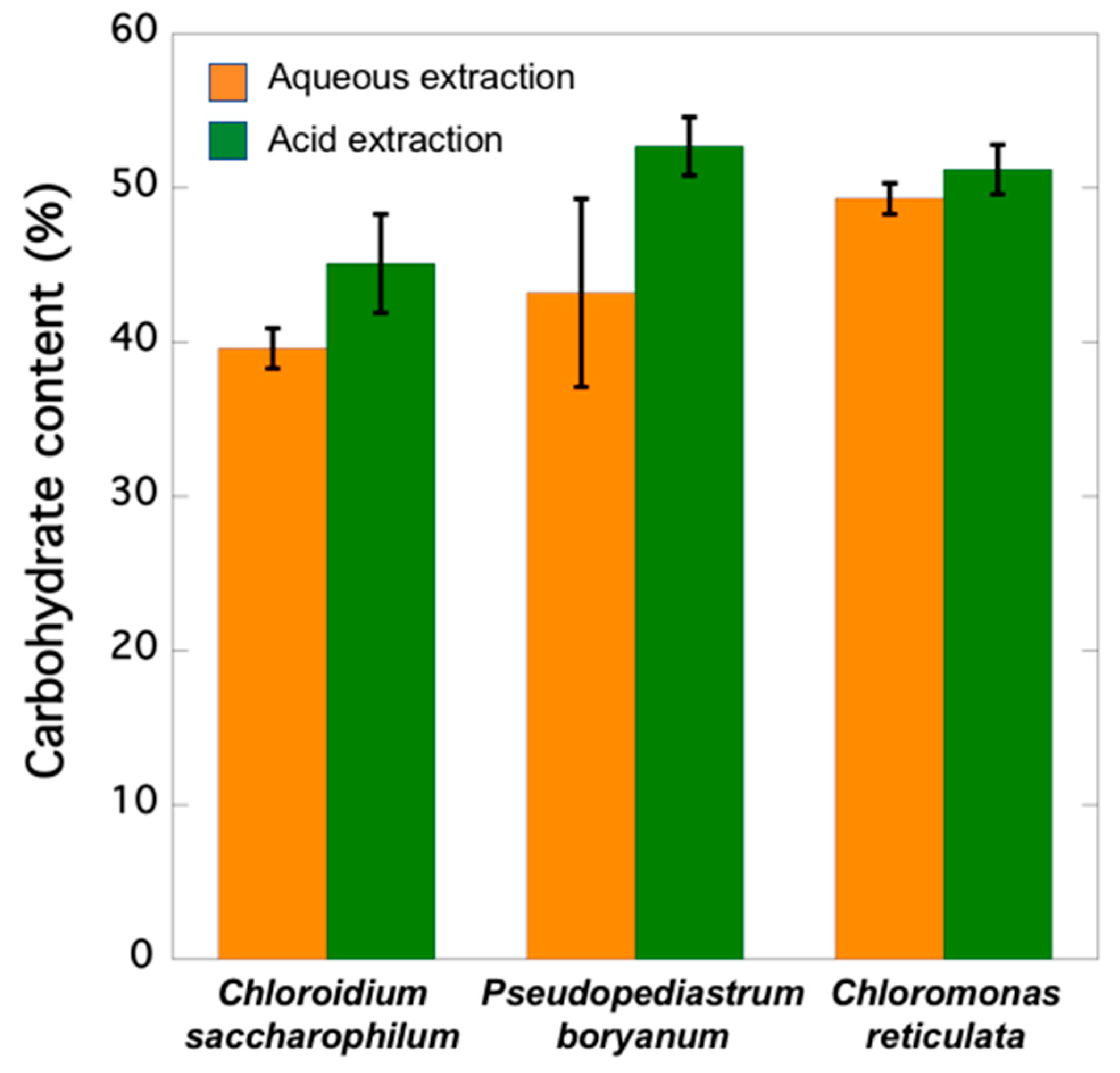
2.4. Carbohydrate Contents
3. Discussion
3.1. Algal Material and Extraction Procedures
3.2. Antioxidant (RSA) Activity and Phenolic Contents
3.3. Free Amino Acid Contents
3.4. Total Contents of Analysed Amino Acids
3.5. Carbohydrate Contents
4. Materials and Methods
4.1. Chemicals
4.2. Algal Material
4.3. Radical Scavenging Activity (RSA) Measurements
4.4. Phenolic Compounds Determination
4.5. Amino Acid Composition Determination
4.6. Carbohydrates Quantification
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UN General Assembly. Transforming Our World: The 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- FAO. The Future of Food and Agriculture-Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Ördög, V.; Stirk, W.A.; Lenobel, R.; Bancířová, M.; Strnad, M.; Van Staden, J.; Szigeti, J.; Németh, L. Screening Microalgae for Some Potentially Useful Agricultural and Pharmaceutical Secondary Metabolites. J. Appl. Phycol. 2004, 16, 309–314. [Google Scholar] [CrossRef]
- Nethravathy, M.U.; Mehar, J.G.; Mudliar, S.N.; Shekh, A.Y. Recent Advances in Microalgal Bioactives for Food, Feed, and Healthcare Products: Commercial Potential, Market Space, and Sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar] [CrossRef]
- Martínez Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Jin, W.; Zhang, F.; Linhardt, R.J. The Application of Seaweed Polysaccharides and Their Derived Products with Potential for the Treatment of Alzheimer’s Disease. Mar. Drugs 2021, 19, 89. [Google Scholar] [CrossRef]
- de Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Emergent Sources of Prebiotics: Seaweeds and Microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Amna Kashif, S.; Hwang, Y.J.; Park, J.K. Potent Biomedical Applications of Isolated Polysaccharides from Marine Microalgae Tetraselmis Species. Bioprocess. Biosyst. Eng. 2018, 41, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids: Metabolism: Functions, and Nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A Potential Alternative to Health Supplementation for Humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Ursu, A.V.; Laroche, C.; Zebib, B.; Pontalier, P.Y.; Vaca-Garcia, C. Release of Hydro-Soluble Microalgal Proteins Using Mechanical and Chemical Treatments. Algal Res. 2014, 3, 55–60. [Google Scholar] [CrossRef]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of Proteins from Two Marine Macroalgae, Ulva sp. and Gracilaria sp., for Food Application, and Evaluating Digestibility, Amino Acid Composition and Antioxidant Properties of the Protein Concentrates. Food Hydrocoll. 2019, 87, 194–203. [Google Scholar] [CrossRef]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-Teles, M.T.; Paula Carvalho, A.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese Coast as a Source of Proteinaceous Material: Total and Free Amino Acid Composition Profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Vitali, A. Proline-Rich Peptides: Multifunctional Bioactive Molecules as New Potential Therapeutic Drugs. Curr. Protein Pept. Sci. 2015, 16, 147–162. [Google Scholar] [CrossRef]
- Zou, C.; Brewer, M.; Cao, X.; Zang, R.; Lin, J.; Deng, Y.; Li, C. Antitumor Activity of 4-(N-Hydroxyphenyl)Retinamide Conjugated with Poly(l-Glutamic Acid) against Ovarian Cancer Xenografts. Gynecol. Oncol. 2007, 107, 441–449. [Google Scholar] [CrossRef] [PubMed]
- You, X.-Y.; Xu, Y.; Huang, Z.-B.; Li, Y.-P. Nonvolatile Taste Compounds of Jiangluobo (A Traditional Chinese Fermented Food). J. Food Qual. 2010, 33, 477–489. [Google Scholar] [CrossRef]
- Trevino, S.R.; Scholtz, J.M.; Pace, C.N. Amino Acid Contribution to Protein Solubility: Asp, Glu, and Ser Contribute More Favorably than the Other Hydrophilic Amino Acids in RNase Sa. J. Mol. Biol. 2007, 366, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and Hydroxyproline Metabolism: Implications for Animal and Human Nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef]
- Sansone, C.; Brunet, C. Promises and Challenges of Microalgal Antioxidant Production. Antioxidants 2019, 8, 199. [Google Scholar] [CrossRef]
- Rico, M.; López, A.; Santana-Casiano, J.M.; González, A.G.; González-Dávila, M. Variability of the Phenolic Profile in the Diatom Phaeodactylum tricornutum Growing under Copper and Iron Stress. Limnol. Oceanogr. 2013, 58, 144–152. [Google Scholar] [CrossRef]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef]
- Nova, P.; Martins, A.P.; Teixeira, C.; Abreu, H.; Silva, J.G.; Silva, A.M.; Freitas, A.C.; Gomes, A.M. Foods with Microalgae and Seaweeds Fostering Consumers Health: A Review on Scientific and Market Innovations. J. Appl. Phycol. 2020, 32, 1789–1802. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernández, F.G.; Castellari, M.; Villaró, S.; Bobo, G.; Aguiló-Aguayo, I. Effect of Microalgae Incorporation on the Physicochemical, Nutritional, and Sensorial Properties of an Innovative Broccoli Soup. LWT 2019, 111, 167–174. [Google Scholar] [CrossRef]
- Žugčić, T.; Abdelkebir, R.; Barba, F.J.; Rezek-Jambrak, A.; Gálvez, F.; Zamuz, S.; Granato, D.; Lorenzo, J.M. Effects of Pulses and Microalgal Proteins on Quality Traits of Beef Patties. J. Food Sci. Technol. 2018, 55, 4544–4553. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chen, T.; Chen, S.H.Y.; Liu, B.; Sun, P.; Sun, H.; Chen, F. The Potentials and Challenges of Using Microalgae as an Ingredient to Produce Meat Analogues. Trends Food Sci. Technol. 2021, 112, 188–200. [Google Scholar] [CrossRef]
- Fernandes, T.; Cordeiro, N. Microalgae as Sustainable Biofactories to Produce High-Value Lipids: Biodiversity, Exploitation, and Biotechnological Applications. Mar. Drugs 2021, 19, 573. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, A.; Bartosz, G.; Čipak, A.; Duburs, G.; Horáková, L.; Łuczaj, W.; Majekova, M.; Odysseos, A.D.; Rackova, L.; Skrzydlewska, E.; et al. Natural and Synthetic Antioxidants: An Updated Overview. Free Radic. Res. 2010, 44, 1216–1262. [Google Scholar] [CrossRef]
- Malavasi, V.; Soru, S.; Cao, G. Extremophile Microalgae: The Potential for Biotechnological Application. J. Phycol. 2020, 56, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Kröger, M.; Klemm, M.; Nelles, M. Extraction Behavior of Different Conditioned, S. Rubescens. Energies 2019, 12, 1336. [Google Scholar] [CrossRef]
- De Farias Neves, F.; Demarco, M.; Tribuzi, G. Drying and Quality of Microalgal Powders for Human Alimentation. In Microalgae: From Physiology to Application; InTech Open: London, UK, 2019; ISBN 978-1-83880-035-2. [Google Scholar]
- Pop, A.; Kiss, B.; Loghin, F. Endocrine Disrupting Effects of Butylated Hydroxyanisole (BHA–E320). Clujul Med. 2013, 86, 16–20. [Google Scholar] [PubMed]
- Saito, M.; Sakagami, H.; Fujisawa, S. Cytotoxicity and Apoptosis Induction by Butylated Hydroxyanisole (BHA) and Butylated Hydroxytoluene (BHT). Anticancer Res. 2003, 23, 4693–4701. [Google Scholar] [PubMed]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Corrêa da Silva, M.G.; Pires Ferreira, S.; Dora, C.L.; Hort, M.A.; Giroldo, D.; Prates, D.F.; Radmann, E.M.; Bemvenuti, R.H.; Costa, J.A.V.; Badiale-Furlong, E.; et al. Phenolic Compounds and Antioxidant Capacity of Pediastrum boryanum (Chlorococcales) Biomass. Int. J. Environ. Health Res. 2020, 1–13. [Google Scholar] [CrossRef]
- Blagojević, D.; Babić, O.; Rašeta, M.; Šibul, F.; Janjušević, L.; Simeunović, J. Antioxidant Activity and Phenolic Profile in Filamentous Cyanobacteria: The Impact of Nitrogen. J. Appl. Phycol. 2018, 30, 2337–2346. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Correlations of Antioxidant Activity against Phenolic Content Revisited: A New Approach in Data Analysis for Food and Medicinal Plants. J. Food Sci. 2009, 74, R107–R113. [Google Scholar] [CrossRef] [PubMed]
- Klejdus, B.; Kopecký, J.; Benešová, L.; Vacek, J. Solid-Phase/Supercritical-Fluid Extraction for Liquid Chromatography of Phenolic Compounds in Freshwater Microalgae and Selected Cyanobacterial Species. J. Chromatogr. A 2009, 1216, 763–771. [Google Scholar] [CrossRef]
- Onofrejová, L.; Vašíčková, J.; Klejdus, B.; Stratil, P.; Mišurcová, L.; Kráčmar, S.; Kopecký, J.; Vacek, J. Bioactive Phenols in Algae: The Application of Pressurized-Liquid and Solid-Phase Extraction Techniques. J. Pharm. Biomed. Anal. 2010, 51, 464–470. [Google Scholar] [CrossRef]
- Hwang, H.-S.; Winkler-Moser, J.K.; Liu, S.X. Study on Antioxidant Activity of Amino Acids at Frying Temperatures and Their Interaction with Rosemary Extract, Green Tea Extract, and Ascorbic Acid. J. Food Sci. 2019, 84, 3614–3623. [Google Scholar] [CrossRef]
- Marquis, V.; Smirnova, E.; Poirier, L.; Zumsteg, J.; Schweizer, F.; Reymond, P.; Heitz, T. Stress- and Pathway-Specific Impacts of Impaired Jasmonoyl-Isoleucine (JA-Ile) Catabolism on Defense Signalling and Biotic Stress Resistance. Plant Cell Environ. 2020, 43, 1558–1570. [Google Scholar] [CrossRef]
- Rubino, J.T.; Franz, K.J. Coordination Chemistry of Copper Proteins: How Nature Handles a Toxic Cargo for Essential Function. J. Inorg. Biochem. 2012, 107, 129–143. [Google Scholar] [CrossRef]
- Dietzen, D. Amino Acids, Peptides, and Proteins. In Principles and Applications of Molecular Diagnostics; Elsevier: Amsterdam, The Netherlands, 2018; pp. 345–380. ISBN 9780128160619. [Google Scholar]
- Vendruscolo, R.G.; Facchi, M.M.X.; Maroneze, M.M.; Fagundes, M.B.; Cichoski, A.J.; Zepka, L.Q.; Barin, J.S.; Jacob-Lopes, E.; Wagner, R. Polar and Non-Polar Intracellular Compounds from Microalgae: Methods of Simultaneous Extraction, Gas Chromatography Determination and Comparative Analysis. Food Res. Int. 2018, 109, 204–212. [Google Scholar] [CrossRef]
- Kolmakova, A.A.; Kolmakov, V.I. Amino Acid Composition of Green Microalgae and Diatoms, Cyanobacteria, and Zooplankton (Review). Ecol. Physiol. Biochem. Hydrobionts 2019, 12, 452–461. [Google Scholar] [CrossRef]
- Peinado, I.; Girón, J.; Koutsidis, G.; Ames, J.M. Chemical Composition, Antioxidant Activity and Sensory Evaluation of Five Different Species of Brown Edible Seaweeds. Food Res. Int. 2014, 66, 36–44. [Google Scholar] [CrossRef]
- Machado, M.; Machado, S.; Pimentel, F.B.; Freitas, V.; Alves, R.C.; Oliveira, M.B.P.P. Amino Acid Profile and Protein Quality Assessment of Macroalgae Produced in an Integrated Multi-Trophic Aquaculture System. Foods 2020, 9, 1382. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic Stresses as Tools for Metabolites in Microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Granum, E.; Kirkvold, S.; Myklestad, S.M. Cellular and Extracellular Production of Carbohydrates and Amino Acids by the Marine Diatom Skeletonema costatum: Diel Variations and Effects of N Depletion. Mar. Ecol. Prog. Ser. 2002, 242, 83–94. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Sournia, A.; Birrien, J.-L. A Daily Study of the Diatom Spring Bloom at Roscoff (France) in 1985. III. Free Amino Acids Composition Studied by HPLC Analysis. J. Plankton Res. 1992, 14, 409–421. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweed Proteins: Biochemical, Nutritional Aspects and Potential Uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Cobos, M.; Pérez, S.; Braga, J.; Vargas-Arana, G.; Flores, L.; Paredes, J.D.; Maddox, J.D.; Marapara, J.L.; Castro, J.C. Nutritional Evaluation and Human Health-Promoting Potential of Compounds Biosynthesized by Native Microalgae from the Peruvian Amazon. World J. Microbiol. Biotechnol. 2020, 36, 121. [Google Scholar] [CrossRef] [PubMed]
- Biancarosa, I.; Espe, M.; Bruckner, C.G.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E.J. Amino Acid Composition, Protein Content, and Nitrogen-to-Protein Conversion Factors of 21 Seaweed Species from Norwegian Waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of Intracellular Nitrogen in Marine Microalgae: Calculation of New Nitrogen-to-Protein Conversion Factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Sui, Y.; Muys, M.; Vermeir, P.; D’Adamo, S.; Vlaeminck, S.E. Light Regime and Growth Phase Affect the Microalgal Production of Protein Quantity and Quality with Dunaliella salina. Bioresour. Technol. 2019, 275, 145–152. [Google Scholar] [CrossRef]
- Araya, M.; García, S.; Rengel, J.; Pizarro, S.; Álvarez, G. Determination of Free and Protein Amino Acid Content in Microalgae by HPLC-DAD with Pre-Column Derivatization and Pressure Hydrolysis. Mar. Chem. 2021, 234, 103999. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced Growth and Lipid Production of Microalgae under Mixotrophic Culture Condition: Effect of Light Intensity, Glucose Concentration and Fed-Batch Cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef]
- Wong, Y. Effects of Light Intensity, Illumination Cycles on Microalgae Haematococcus pluvialis for Production of Astaxanthin. J. Mar. Biol. Aquac. 2016, 2, 1–6. [Google Scholar] [CrossRef][Green Version]
- Seyfabadi, J.; Ramezanpour, Z.; Khoeyi, Z.A. Protein, Fatty Acid, and Pigment Content of Chlorella Vulgaris under Different Light Regimes. J. Appl. Phycol. 2011, 23, 721–726. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein Content and Amino Acid Composition of Commercially Available Plant-Based Protein Isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef]
- Schulze, C.; Strehle, A.; Merdivan, S.; Mundt, S. Carbohydrates in Microalgae: Comparative Determination by TLC, LC-MS without Derivatization, and the Photometric Thymol-Sulfuric Acid Method. Algal Res. 2017, 25, 372–380. [Google Scholar] [CrossRef]
- Templeton, D.W.; Quinn, M.; Van Wychen, S.; Hyman, D.; Laurens, L.M.L. Separation and Quantification of Microalgal Carbohydrates. J. Chromatogr. A 2012, 1270, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Visca, A.; Di Caprio, F.; Spinelli, R.; Altimari, P.; Cicci, A.; Iaquaniello, G.; Toro, L.; Pagnanelli, F. Microalgae Cultivation for Lipids and Carbohydrates Production. Chem. Eng. Trans. 2017, 57, 127–132. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Vambe, M.; Lovász, C.; Molnár, Z.; van Staden, J.; Ördög, V. Effect of Cell Disruption Methods on the Extraction of Bioactive Metabolites from Microalgal Biomass. J. Biotechnol. 2020, 307, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, K.; Thirumaran, G.; Devi, G.K.; Hemalatha, A.; Anantharaman, P. Biochemical Composition of Seaweeds from Mandapam Coastal Regions along Southeast Coast of India. Am. Eurasian J. Bot. 2008, 1, 32–37. [Google Scholar]
- Suarez Garcia, E.; Suarez Ruiz, C.A.; Tilaye, T.; Eppink, M.H.M.; Wijffels, R.H.; van den Berg, C. Fractionation of Proteins and Carbohydrates from Crude Microalgae Extracts Using an Ionic Liquid Based-Aqueous Two Phase System. Sep. Purif. Technol. 2018, 204, 56–65. [Google Scholar] [CrossRef]
- Jutur, P.P.; Nesamma, A.A.; Shaikh, K.M. Algae-Derived Marine Oligosaccharides and Their Biological Applications. Front. Mar. Sci. 2016, 3, 83. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Lucas, B.F.; Alvarenga, A.G.P.; Moreira, J.B.; de Morais, M.G. Microalgae Polysaccharides: An Overview of Production, Characterization, and Potential Applications. Polysaccharides 2021, 2, 759–772. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and Mechanisms of Antioxidant Activity Using the DPPH·Free Radical Method. LWT Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Dvořáková, M.; Hulín, P.; Karabín, M.; Dostálek, P. Determination of Polyphenols in Beer by an Effective Method Based on Solid-Phase Extraction and High Performance Liquid Chromatography with Diode-Array Detection. Czech J. Food Sci. 2007, 25, 182–188. [Google Scholar] [CrossRef]
- Shi, Z.; Li, H.; Li, Z.; Hu, J.; Zhang, H. Pre-Column Derivatization RP-HPLC Determination of Amino Acids in Asparagi Radix before and after Heating Process. IERI Procedia 2013, 5, 351–356. [Google Scholar] [CrossRef]
- Jansen, E. Simple Determination of Sugars in Cigarettes. J. Anal. Bioanal. Tech. 2014, 5, 5–7. [Google Scholar] [CrossRef]
- Brooks, J.R.; Griffin, V.K.; Kattan, M.W. A Modified Method for Total Carbohydrate Analysis of Glucose Syrups, Maltodextrins, and Other Starch Hydrolysis Products. Cereal Chem. 1986, 63, 465–466. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenol | Chloromonas cf. reticulata | Pseudopediastrum boryanum | Chloroidium saccharophilum |
|---|---|---|---|
| Gallic acid | 5.08 ± 0.25 | 1.44 ± 0.99 | 2.88 ± 1.22 |
| Protocatechuic acid | n.d. * | 1.23 ± 0.11 | 3.08 ± 1.34 |
| Catechin | 5.68 ± 0.74 | 4.14 ± 0.13 | 12.34 ± 1.13 |
| Vanillic acid | 1.61 ± 0.21 | 2.63 ± 0.23 | 14.15 ± 2.84 |
| Epicatechin | 10.43 ± 0.21 | 5.27 ± 0.84 | 1.90 ± 0.33 |
| Syringic acid | 3.12 ± 0.32 | 1.12 ± 0.10 | 5.84 ± 0.66 |
| Rutin | n.d. * | 6.40 ± 1.03 | 6.87 ± 3.42 |
| Gentisic acid | n.d. * | n.d. * | 2.42 ± 2.21 |
| Coumaric acid | n.d. * | n.d. * | 3.95 ± 2.09 |
| Ferulic acid | n.d. * | 2.44 ± 0.34 | 2.40 ± 1.67 |
| Total | 27.10 ± 2.03 | 26.40 ± 4.02 | 55.83 ± 16.90 |
| Amino Acid | Chloromonas cf. reticulata | Pseudopediastrum boryanum | Chloroidium saccharophilum |
|---|---|---|---|
| Arginine | 107.88 ± 1.67 | 448.00 ± 90.00 | 668.34 ± 48.16 |
| Glutamic acid | 461.82 ± 1.70 | 1937.00 ± 28.00 | 5630.37 ± 135.89 |
| Aspartic acid | 89.31 ± 0.55 | 157.57 ± 16.49 | 805.95 ± 18.82 |
| Proline | 112.99 ± 0.51 | 1282.00 ± 38.00 | 5546.42 ± 141.14 |
| Methionine | 146.00 ± 6.30 | 201.00 ± 77.00 | 805.95 ± 90.52 |
| Valine | 130.00 ± 0.38 | 285.00 ± 6.14 | 1379.65 ± 159.98 |
| Lysine | 196.84 ± 0.87 | 697.65 ± 52.14 | 2847.93 ± 157.67 |
| Isoleucine | 65.34 ± 0.08 | 86.03 ± 4.00 | 866.81 ± 15.26 |
| Phenylalanine | 107.42 ± 0.19 | 108.00 ± 2.00 | 1379.65 ± 30.57 |
| Histidine | 116.67 ± 1.22 | 154.00 ± 21.00 | 530.92 ± 38.35 |
| Sum of amino acids in µg g−1 of dry weight | |||
| ∑NEEA | 772.0 ± 4.43 | 3824.6 ± 172.5 | 12,651.1 ± 344.0 |
| ∑EEA | 762.3 ± 9.04 | 1531.7 ± 162.3 | 7810.9 ± 492.5 |
| ∑FAA | 1534.33 ± 13.5 | 5356.3 ± 334.8 | 20,462.0 ± 836.5 |
| Amino Acid | Chloromonas cf. reticulata | Pseudopediastrum boryanum | Chloroidium saccharophilum |
|---|---|---|---|
| Arginine | 21.29 ± 3.47 | 7.12 ± 0.75 | 5.98 ± 0.66 |
| Glutamic acid | 23.84 ± 6.25 | 19.48 ± 1.41 | 30.50 ± 1.28 |
| Aspartic acid | 14.67 ± 3.59 | 11.38 ± 0.84 | 3.44 ± 0.21 |
| Proline | 51.69 ± 1.75 | 49.90 ± 4.25 | 31.68 ± 1.42 |
| Methionine | 40.81 ± 9.78 | 137.20 ± 24.41 | 29.89 ± 8.96 |
| Valine | 19.72 ± 1.79 | 20.21 ± 1.59 | 18.48 ± 0.45 |
| Lysine | 31.56 ± 1.66 | 36.20 ± 2.32 | 36.49 ± 1.99 |
| Isoleucine | 13.28 ± 1.35 | 13.43 ± 1.32 | 11.36 ± 0.71 |
| Phenylalanine | 19.76 ± 1.37 | 20.01 ± 1.20 | 14.92 ± 0.21 |
| Histidine | 6.07 ± 0.13 | 5.63 ± 0.69 | 1.81 ± 0.01 |
| Sum of amino acids in mg g−1 of dry weight (%) | |||
| ∑NEEA | 111.5 ± 15.1 (45.9) | 87.9 ± 7.25 (27.4) | 71.6 ± 3.57 (38.8) |
| ∑EEA | 131.2 ± 16.1 (54.1) | 232.7 ± 30 (72.6) | 112.9 ± 11.3 (61.2) |
| ∑TAA | 242.7 ± 31.1 | 320. 6 ± 38.1 | 184.5 ± 14.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago-Díaz, P.; Rivero, A.; Rico, M.; Gómez-Pinchetti, J.L. Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates. Mar. Drugs 2022, 20, 40. https://doi.org/10.3390/md20010040
Santiago-Díaz P, Rivero A, Rico M, Gómez-Pinchetti JL. Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates. Marine Drugs. 2022; 20(1):40. https://doi.org/10.3390/md20010040
Chicago/Turabian StyleSantiago-Díaz, Paula, Argimiro Rivero, Milagros Rico, and Juan Luis Gómez-Pinchetti. 2022. "Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates" Marine Drugs 20, no. 1: 40. https://doi.org/10.3390/md20010040
APA StyleSantiago-Díaz, P., Rivero, A., Rico, M., & Gómez-Pinchetti, J. L. (2022). Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates. Marine Drugs, 20(1), 40. https://doi.org/10.3390/md20010040