
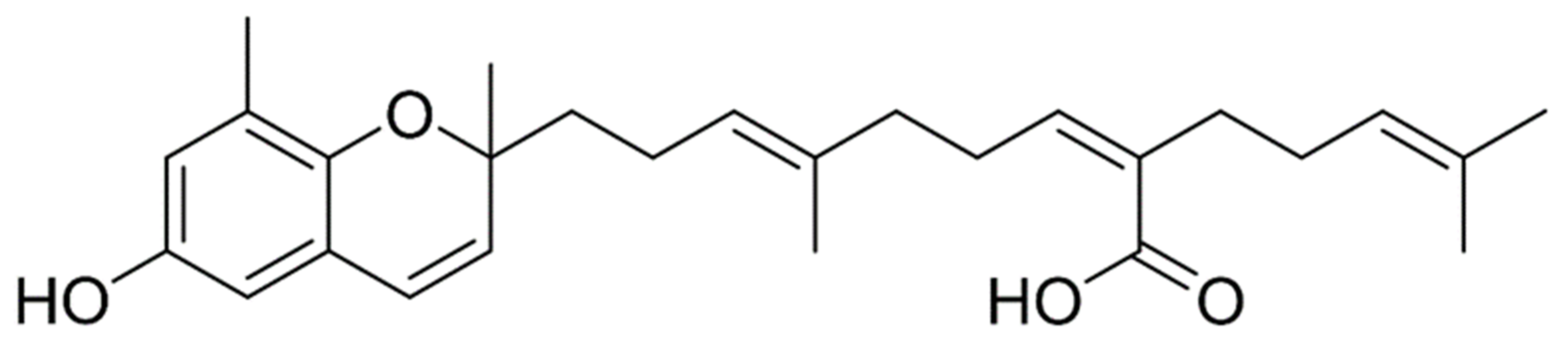
Sargachromenol Isolated from Sargassum horneri Inhibits Particulate Matter-Induced Inflammation in Macrophages through Toll-like Receptor-Mediated Cell Signaling Pathways


 , , ,
, , ,  and
and 
Abstract
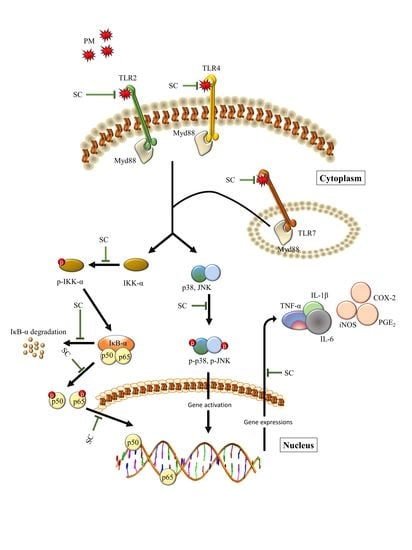
:
1. Introduction
2. Results
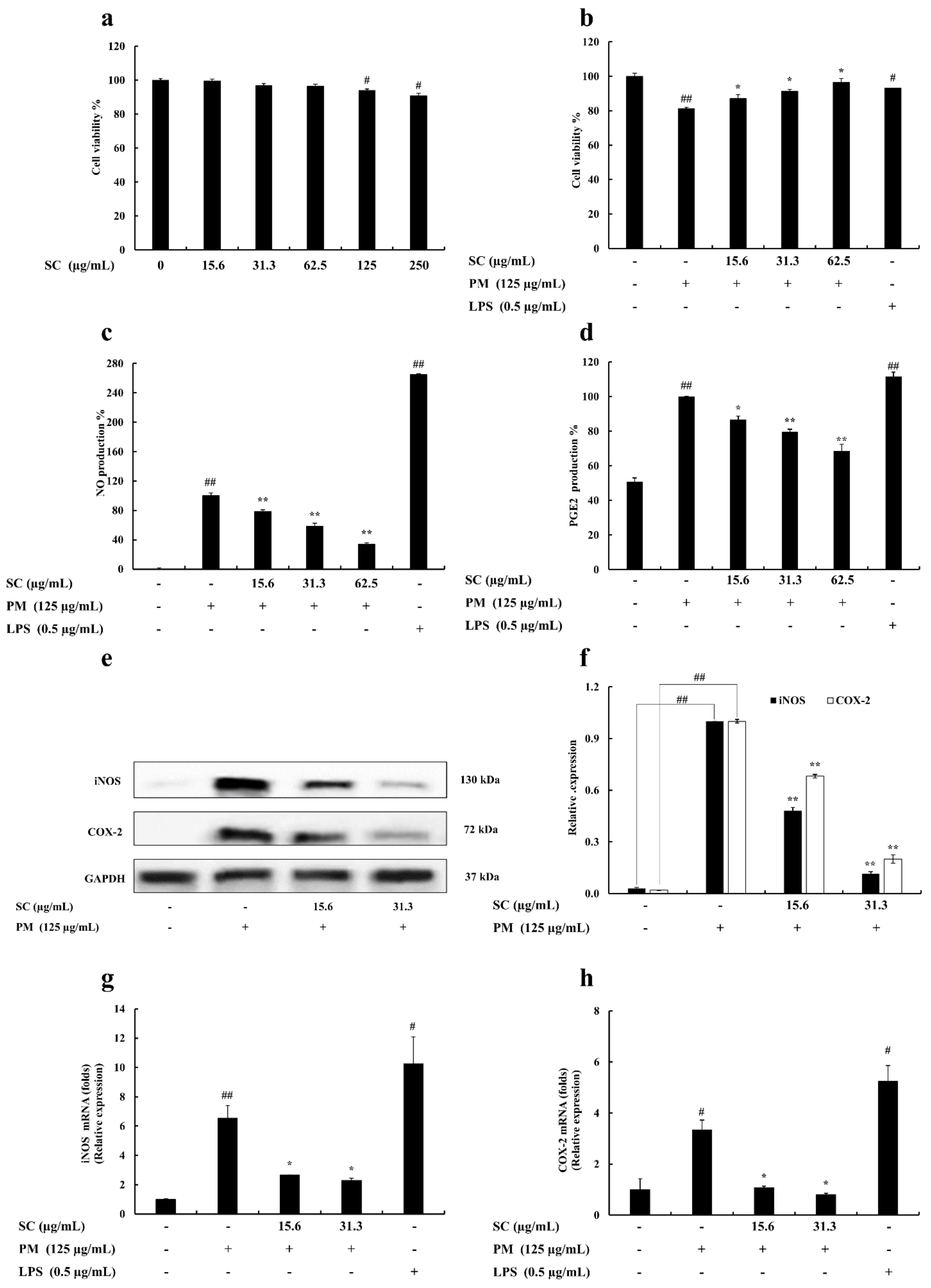
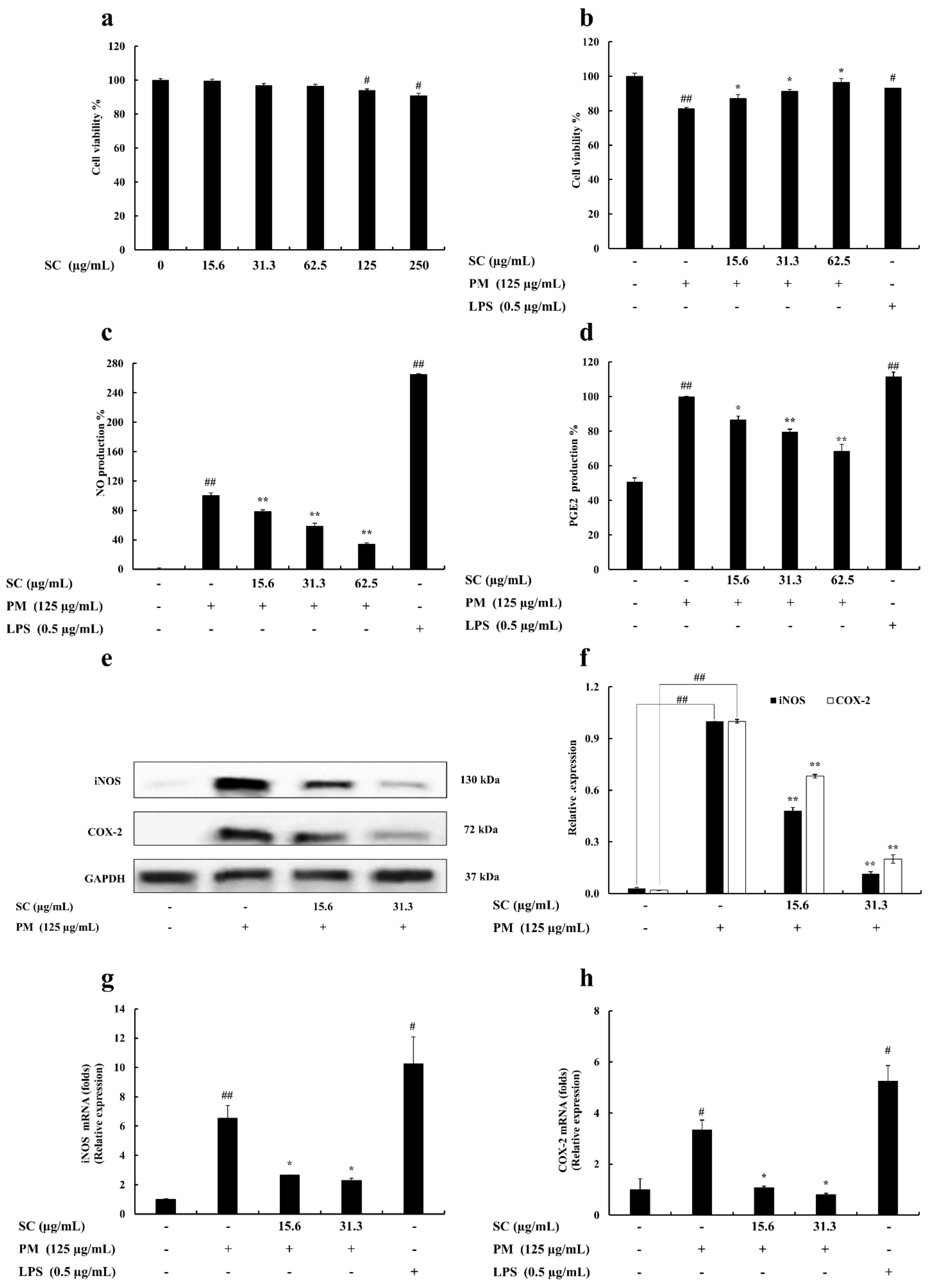
2.1. Cell Viability
2.2. Determination of Nitric Oxide (NO) and Prostaglandin E2 (PGE2) Production
2.3. Evaluation of Inducible Nitric Oxide Synthase (iNOS) and Cyclooxygenase-2 (COX-2) Levels in RAW 264.7 Macrophages
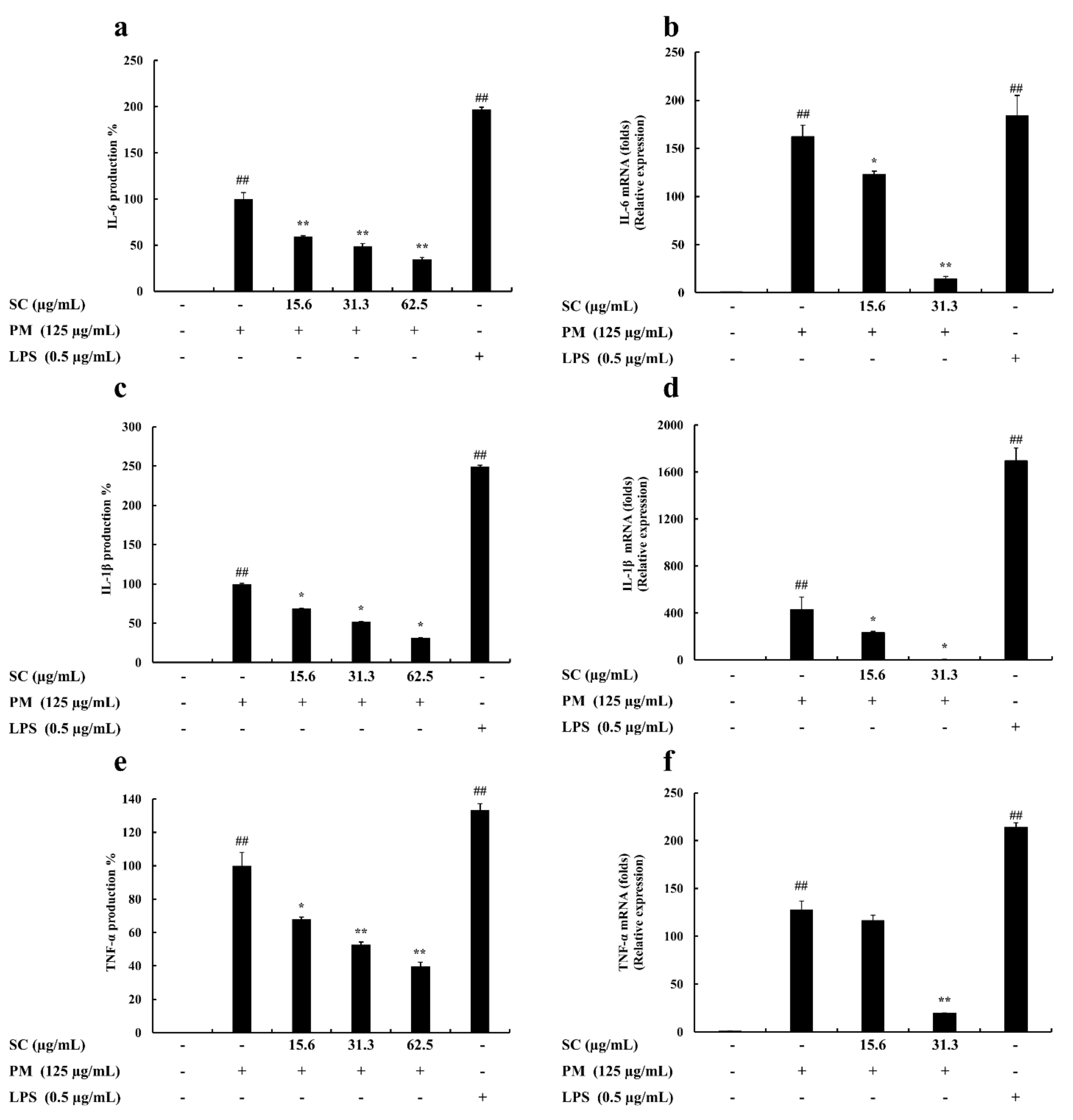
2.4. Evaluation of Cytokine Production
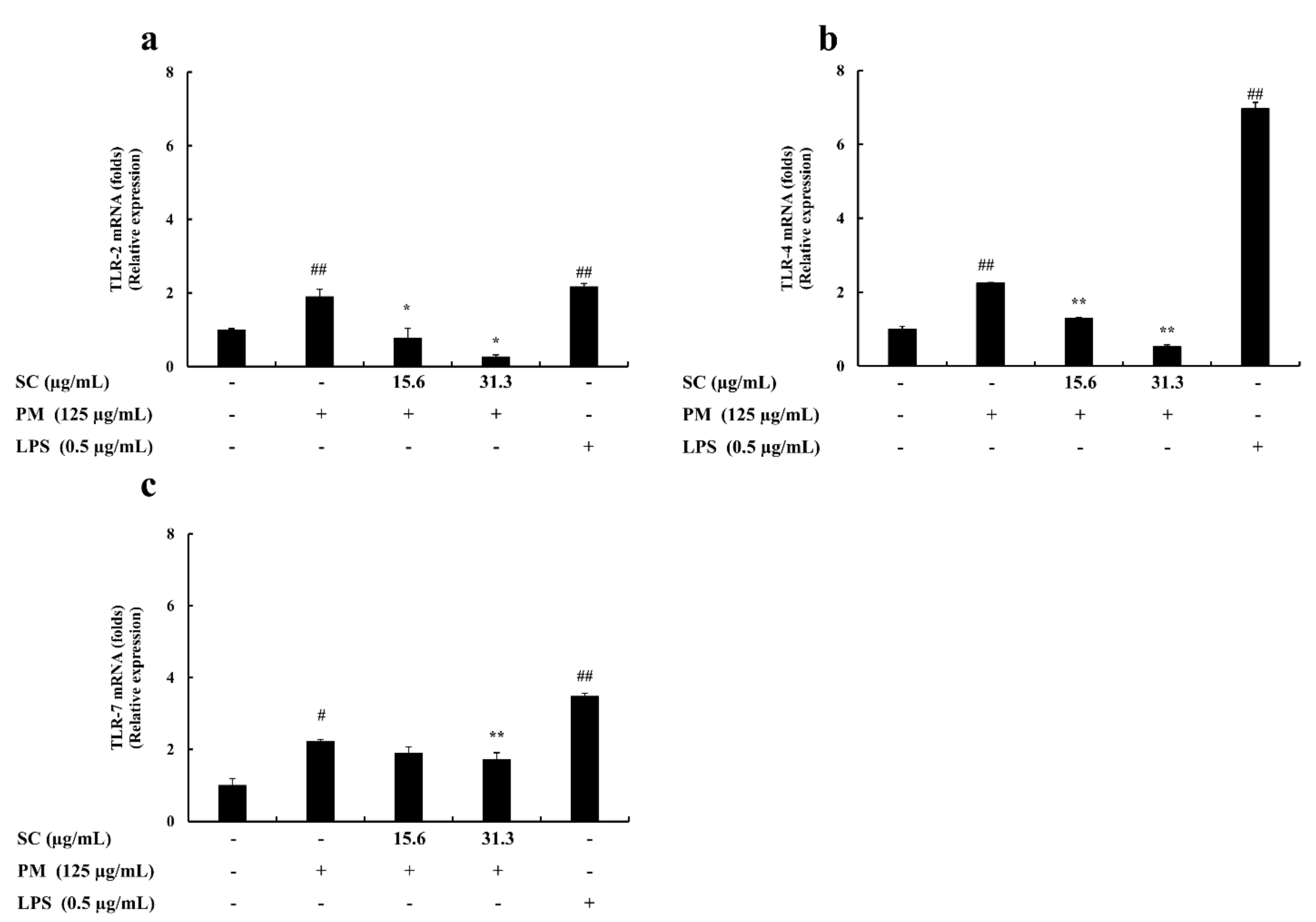
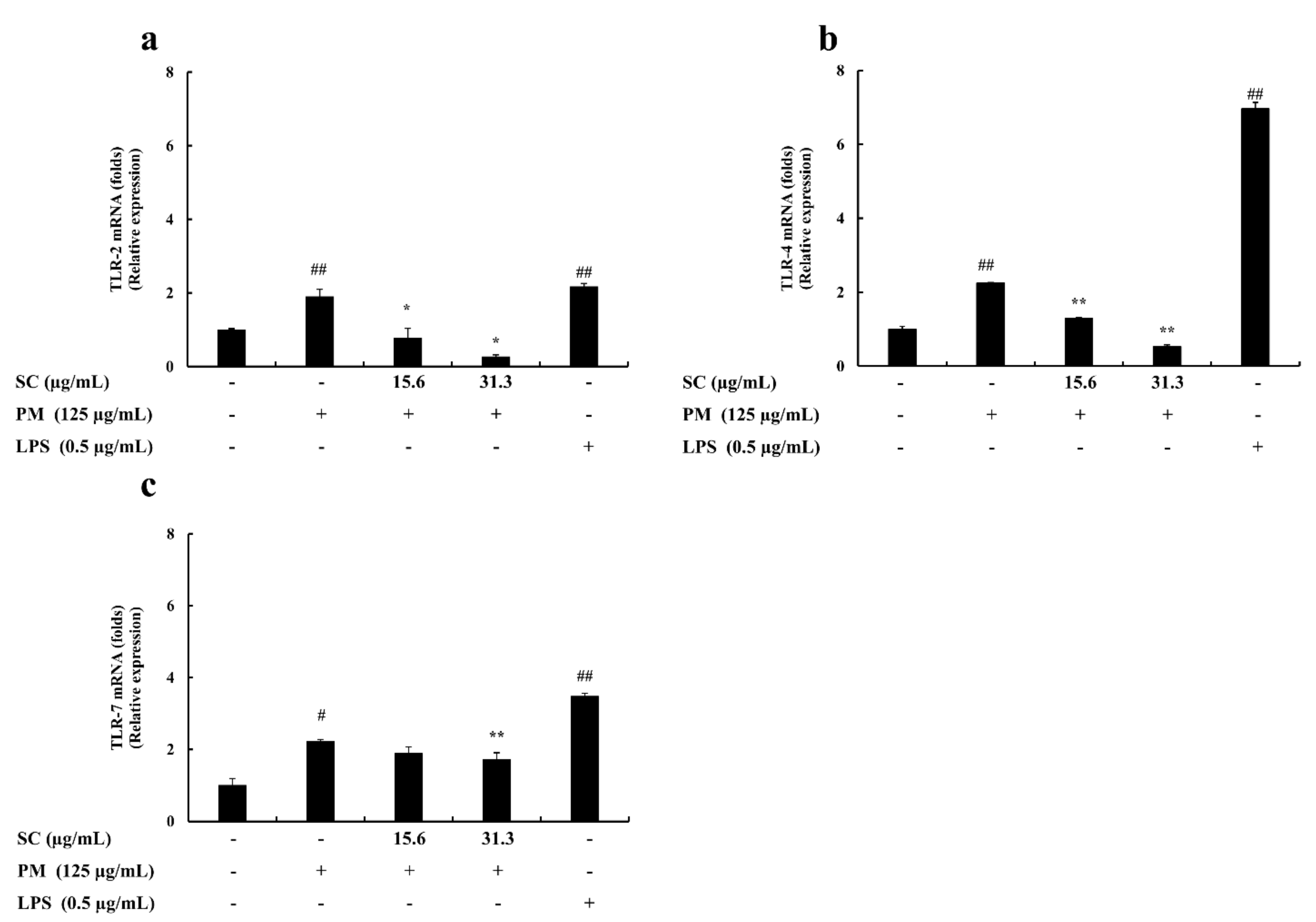
2.5. Gene Expression Levels Evaluation
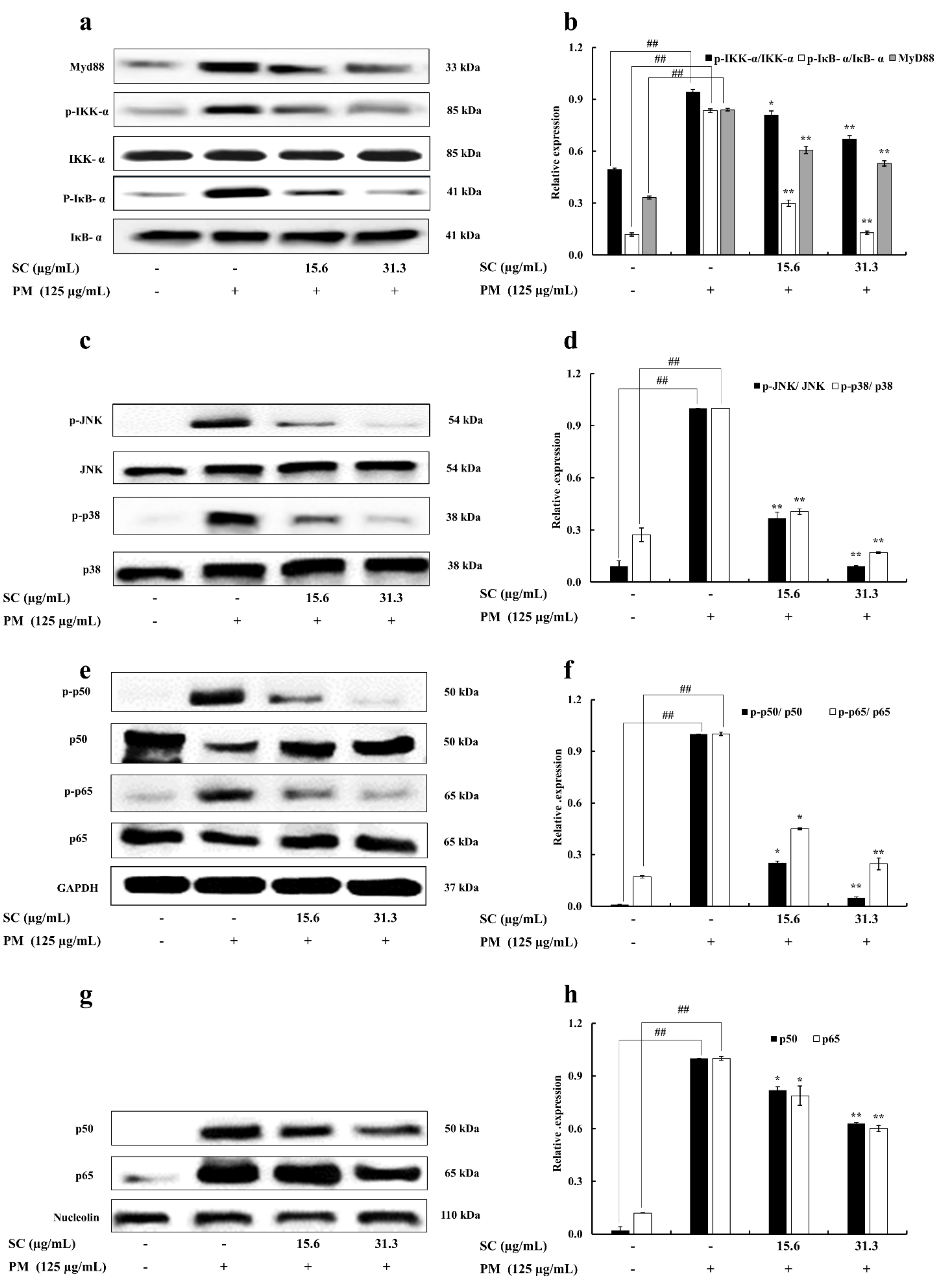
2.6. Western Blot Analysis
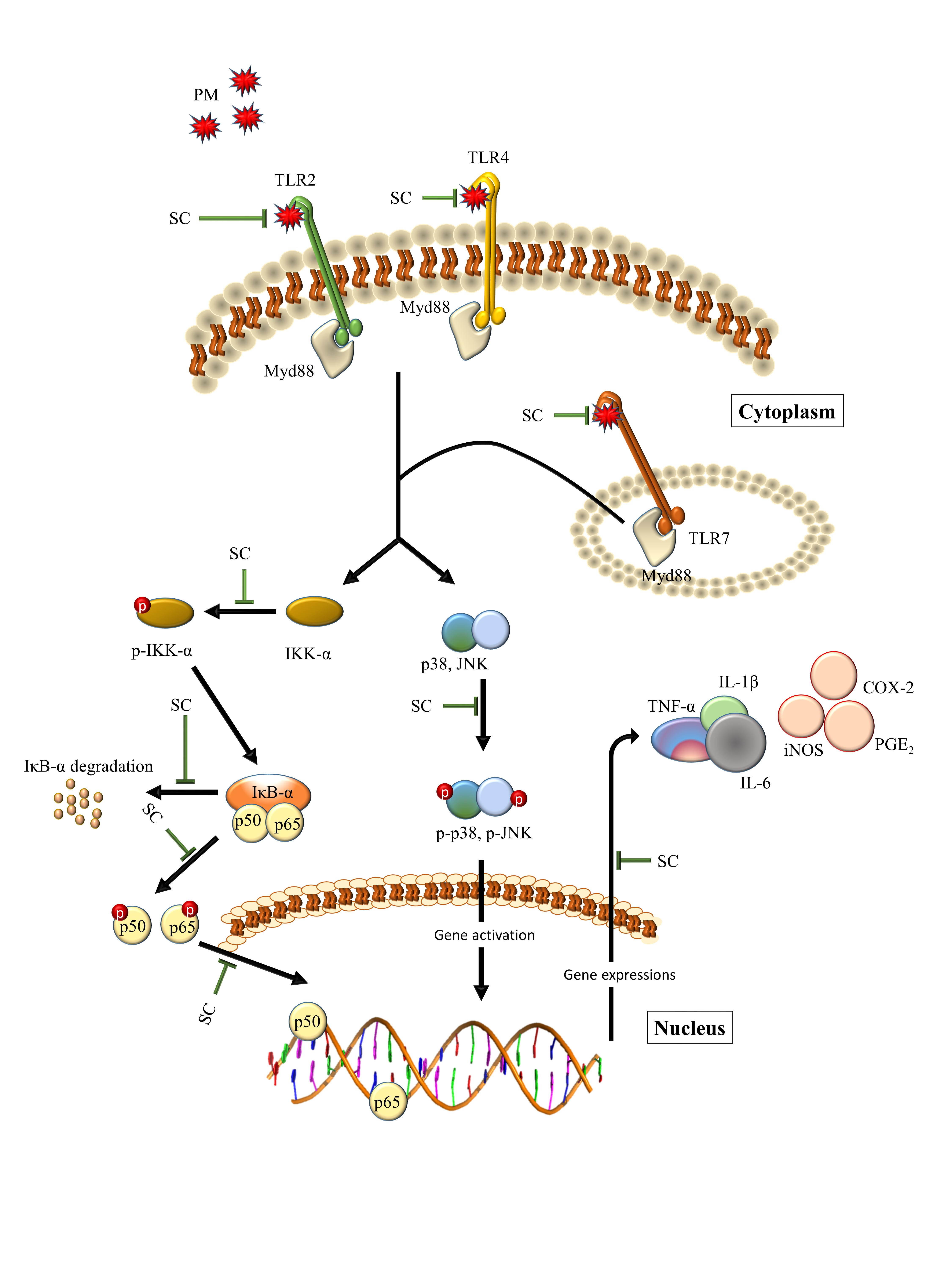
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Chemicals and Reagents
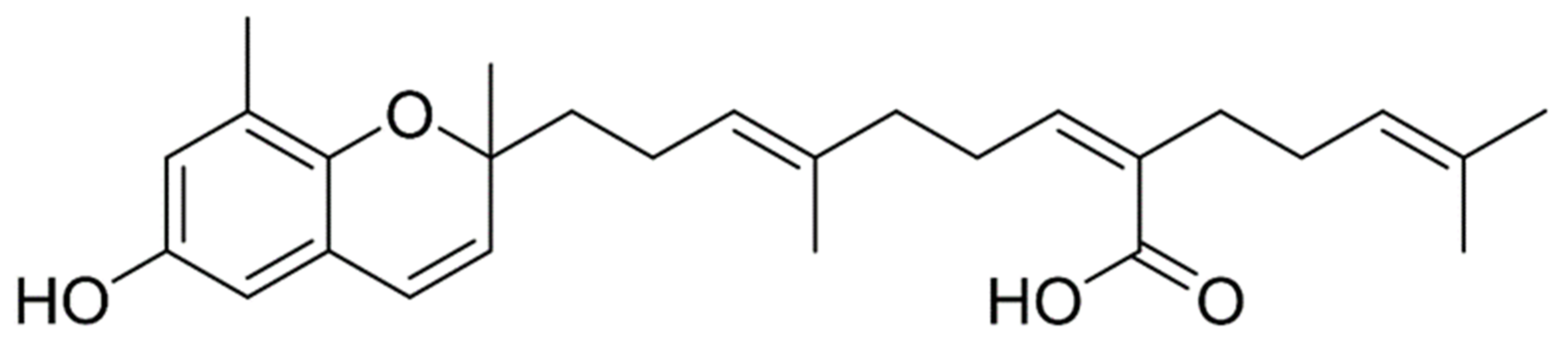
4.3. Preparation of 70% Ethanol Extract and Compound Isolation
4.4. Cell Culture and Sample Preparation
4.5. Dose Range Determination
4.6. Determination of Cell Viability
4.7. Determination of Nitric Oxide (NO) Production
4.8. Evaluation of the Production of Cytokines and PGE2
4.9. Western Blot Analysis
4.10. Total RNA Extraction and cDNA Production
4.11. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Dockery, D.W.; Pope, C.A., 3rd. Acute respiratory effects of particulate air pollution. Annu. Rev. Public Health 1994, 15, 107–132. [Google Scholar] [CrossRef]
- Pope, C.A., 3rd; Burnett, R.T.; Thun, M.J.; Calle, E.E.; Krewski, D.; Ito, K.; Thurston, G.D. Lung cancer, cardiopulmonary mortality, and long-term exposure to fine particulate air pollution. JAMA 2002, 287, 1132–1141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Tsunekawa, A.; Tsubo, M. Contributions of sandy lands and stony deserts to long-distance dust emission in China and Mongolia during 2000–2006. Glob. Planet. Change 2008, 60, 487–504. [Google Scholar] [CrossRef]
- Qian, W.H.; Quan, L.S.; Shi, S.Y. Variations of the dust storm in China and its climatic control. J. Clim. 2002, 15, 1216–1229. [Google Scholar] [CrossRef]
- Ichinose, T.; Nishikawa, M.; Takano, H.; Sera, N.; Sadakane, K.; Mori, I.; Yanagisawa, R.; Oda, T.; Tamura, H.; Hiyoshi, K.; et al. Pulmonary toxicity induced by intratracheal instillation of Asian yellow dust (Kosa) in mice. Environ. Toxicol. Pharmacol. 2005, 20, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Sheen, P.C.; Chen, E.R.; Liu, Y.K.; Wu, T.N.; Yang, C.Y. Effects of Asian dust storm events on daily mortality in Taipei, Taiwan. Environ. Res. 2004, 95, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Timblin, C.; BeruBe, K.; Gordon, T.; McKinney, W.; Driscoll, K.; Vacek, P.; Mossman, B.T. Inhaled particulate matter causes expression of nuclear factor (NF)-kappaB-related genes and oxidant-dependent NF-kappaB activation in vitro. Am. J Respir. Cell Mol. Biol. 2000, 23, 182–187. [Google Scholar] [CrossRef]
- Jeffrey, K.L.; Camps, M.; Rommel, C.; Mackay, C.R. Targeting dual-specificity phosphatases: Manipulating MAP kinase signalling and immune responses. Nat. Rev. Drug Discov. 2007, 6, 391–403. [Google Scholar] [CrossRef]
- Higashisaka, K.; Fujimura, M.; Taira, M.; Yoshida, T.; Tsunoda, S.; Baba, T.; Yamaguchi, N.; Nabeshi, H.; Yoshikawa, T.; Nasu, M.; et al. Asian dust particles induce macrophage inflammatory responses via mitogen-activated protein kinase activation and reactive oxygen species production. J. Immunol. Res. 2014, 2014, 856154. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-Like Receptor Signaling Pathways. Front. Immunol 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-J.; Kim, S.-M.; Kim, H.-G.; Oh, H.-R.; Lee, K.-B.; Lee, Y.-K.; Park, Y.-I. Purification and Anticoagulant Activity of a Fucoidan from Korean Undaria pinnatifida Sporophyll. Algae 2007, 22, 247–252. [Google Scholar] [CrossRef]
- Erbert, C.; Lopes, A.A.; Yokoya, N.S.; Furtado, N.A.J.C.; Conti, R.; Pupo, M.T.; Lopes, J.L.C.; Debonsi, H.M. Antibacterial compound from the endophytic fungus Phomopsis longicolla isolated from the tropical red seaweed Bostrychia radicans. Bot. Mar. 2012, 55, 435–440. [Google Scholar] [CrossRef]
- Ku, C.M.; Lin, J.Y. Anti-inflammatory effects of 27 selected terpenoid compounds tested through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food Chem. 2013, 141, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- NIZAMUDDIN, M. Classification and the distribution of the Fucales. J. Bot. Mar. 1962, 4, 191–203. [Google Scholar] [CrossRef]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar]
- Lee, S.H.; Kwak, C.H.; Lee, S.K.; Ha, S.H.; Park, J.; Chung, T.W.; Ha, K.T.; Suh, S.J.; Chang, Y.C.; Chang, H.W.; et al. Anti-Inflammatory Effect of Ascochlorin in LPS-Stimulated RAW 264.7 Macrophage Cells Is Accompanied With the Down-Regulation of iNOS, COX-2 and Proinflammatory Cytokines Through NF-kappaB, ERK1/2, and p38 Signaling Pathway. J. Cell Biochem. 2016, 117, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, Y.H.; Li, C.; Wang, X.L.; He, X.J. Triterpenes isolated from acorns of Quercus serrata var. brevipetiolata exert anti-inflammatory activity. Ind. Crop. Prod. 2016, 91, 302–309. [Google Scholar] [CrossRef]
- Hseu, Y.C.; Wu, F.Y.; Wu, J.J.; Chen, J.Y.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Yang, H.L. Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kappaB pathway. Int. Immunopharmacol. 2005, 5, 1914–1925. [Google Scholar] [CrossRef]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Attur, M.G.; Patel, R.; Thakker, G.; Vyas, P.; Levartovsky, D.; Patel, P.; Naqvi, S.; Raza, R.; Patel, K.; Abramson, D.; et al. Differential anti-inflammatory effects of immunosuppressive drugs: Cyclosporin, rapamycin and FK-506 on inducible nitric oxide synthase, nitric oxide, cyclooxygenase-2 and PGE2 production. Inflamm. Res. 2000, 49, 20–26. [Google Scholar] [CrossRef]
- Dinarello, C.A. The paradox of pro-inflammatory cytokines in cancer. Cancer Metastasis Rev. 2006, 25, 307–313. [Google Scholar] [CrossRef]
- Chen, C.C.; Wang, J.K.; Chen, W.C.; Lin, S.B. Protein kinase C eta mediates lipopolysaccharide-induced nitric-oxide synthase expression in primary astrocytes. J. Biol. Chem. 1998, 273, 19424–19430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma-Uszynski, S.; Stenger, S.; Takeuchi, O.; Ochoa, M.T.; Engele, M.; Sieling, P.A.; Barnes, P.F.; Rollinghoff, M.; Bolcskei, P.L.; Wagner, M.; et al. Induction of direct antimicrobial activity through mammalian toll-like receptors. Science 2001, 291, 1544–1547. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. TLR Signaling Pathways, Seminars in Immunology; Elsevier: Amsterdam, The Netherlands, 2004; pp. 3–9. [Google Scholar]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.M.; Rajapakse, N.; Kim, S.K. Anti-inflammatory effect of Ishige okamurae ethanolic extract via inhibition of NF-kappaB transcription factor in RAW 264.7 cells. Phytother. Res. 2009, 23, 628–634. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Boutros, T.; Chevet, E.; Metrakos, P. Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase regulation: Roles in cell growth, death, and cancer. Pharmacol. Rev. 2008, 60, 261–310. [Google Scholar] [CrossRef]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Kuan, C.Y.; Yang, D.D.; Samanta Roy, D.R.; Davis, R.J.; Rakic, P.; Flavell, R.A. The Jnk1 and Jnk2 protein kinases are required for regional specific apoptosis during early brain development. Neuron 1999, 22, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Hui, L.; Bakiri, L.; Stepniak, E.; Wagner, E.F. p38alpha: A suppressor of cell proliferation and tumorigenesis. Cell Cycle 2007, 6, 2429–2433. [Google Scholar] [CrossRef]
- Schieven, G.L. The biology of p38 kinase: A central role in inflammation. Curr. Top. Med. Chem. 2005, 5, 921–928. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, H.-S.; Kim, S.-Y.; Ahn, G.; Kim, H.-J.; Fu, X.; Jee, Y.; Jeon, Y.-J. Ethanol extract separated from Sargassum horneri (Turner) abate LPS-induced inflammation in RAW 264.7 macrophages. Fish. Aquat. Sci. 2019, 22, 6. [Google Scholar] [CrossRef] [Green Version]
- Han, E.-J.; Jayawardena, T.U.; Jang, J.-H.; Fernando, I.P.S.; Jee, Y.; Jeon, Y.-J.; Lee, D.-S.; Lee, J.-M.; Yim, M.-J.; Wang, L.; et al. Sargachromenol Purified from Sargassum horneri Inhibits Inflammatory Responses via Activation of Nrf2/HO-1 Signaling in LPS-Stimulated Macrophages. Mar. Drugs 2021, 19, 497. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Leiro, J.; Alvarez, E.; Garcia, D.; Orallo, F. Resveratrol modulates rat macrophage functions. Int. Immunopharmacol. 2002, 2, 767–774. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.; Fernando, I.P.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, T.U.; Kim, H.S.; Sanjeewa, K.K.A.; Kim, S.Y.; Rho, J.R.; Jee, Y.; Ahn, G.; Jeon, Y.J. Sargassum horneri and isolated 6-hydroxy-4,4,7a-trimethyl-5,6,7,7a-tetrahydrobenzofuran-2(4H)-one (HTT); LPS-induced inflammation attenuation via suppressing NF-kappa B, MAPK and oxidative stress through Nrf2/HO-1 pathways in RAW 264.7 macrophages. Algal Res. 2019, 40, 101513. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| GAPDH | Sense | 5′-AAGGGTCATCATCTCTGCCC-3′ |
| Antisense | 5′-GTGATGGCATGGACTGTGGT-3′ | |
| iNOS | Sense | 5′-ATGTCCGAAGCAAACATCAC-3′ |
| Antisense | 5′-TAATGTCCAGGAAGTAGGTG-3′ | |
| COX-2 | Sense | 5′-CAGCAAATCCTTGCTGTTCC-3′ |
| Antisense | 5′-TGGGCAAAGAATGCAAACATC-3′ | |
| IL-6 | Sense | 5′-GTACTCCAGAAGACCAGAGG-3′ |
| Antisense | 5′-TGCTGGTGACAACCACGGCC-3′ | |
| IL-1β | Sense | 5′-CAGGATGAGGACATGAGCACC-3′ |
| Antisense | 5′-CTCTGCAGACTCAAACTCCAC-3′ | |
| TNF-α | Sense | 5′-TTGACCTCAGCGCTGAGTTG-3′ |
| Antisense | 5′-CCTGTAGCCCACGTCGTAGC-3′ | |
| TLR-2 | Sense | 5′-CAGCTGGAGAACTCTGACCC-3′ |
| Antisense | 5′-CAAAGAGCCTGAAGTGGGAG-3′ | |
| TLR-4 | Sense | 5′-CAACATCATCCAGGAAGGC-3 |
| Antisense | 5′-GAAGGCGATACAATTCCACC-3′ | |
| TLR-7 | Sense | 5′-TTCCTTCCGTAGGCTGAACC-3′ |
| Antisense | 5′-GTAAGCTGGATGGCAGATCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagahawatta, D.P.; Kim, H.-S.; Jee, Y.-H.; Jayawardena, T.U.; Ahn, G.; Namgung, J.; Yeo, I.-K.; Sanjeewa, K.K.A.; Jeon, Y.-J. Sargachromenol Isolated from Sargassum horneri Inhibits Particulate Matter-Induced Inflammation in Macrophages through Toll-like Receptor-Mediated Cell Signaling Pathways. Mar. Drugs 2022, 20, 28. https://doi.org/10.3390/md20010028
Nagahawatta DP, Kim H-S, Jee Y-H, Jayawardena TU, Ahn G, Namgung J, Yeo I-K, Sanjeewa KKA, Jeon Y-J. Sargachromenol Isolated from Sargassum horneri Inhibits Particulate Matter-Induced Inflammation in Macrophages through Toll-like Receptor-Mediated Cell Signaling Pathways. Marine Drugs. 2022; 20(1):28. https://doi.org/10.3390/md20010028
Chicago/Turabian StyleNagahawatta, D. P., Hyun-Soo Kim, Young-Heun Jee, Thilina U. Jayawardena, Ginnae Ahn, Jin Namgung, In-Kyu Yeo, K. K. Asanka Sanjeewa, and You-Jin Jeon. 2022. "Sargachromenol Isolated from Sargassum horneri Inhibits Particulate Matter-Induced Inflammation in Macrophages through Toll-like Receptor-Mediated Cell Signaling Pathways" Marine Drugs 20, no. 1: 28. https://doi.org/10.3390/md20010028
APA StyleNagahawatta, D. P., Kim, H.-S., Jee, Y.-H., Jayawardena, T. U., Ahn, G., Namgung, J., Yeo, I.-K., Sanjeewa, K. K. A., & Jeon, Y.-J. (2022). Sargachromenol Isolated from Sargassum horneri Inhibits Particulate Matter-Induced Inflammation in Macrophages through Toll-like Receptor-Mediated Cell Signaling Pathways. Marine Drugs, 20(1), 28. https://doi.org/10.3390/md20010028