
Secondary Metabolite Production Potential of Mangrove-Derived Streptomyces olivaceus



{kind=link}
Abstract
1. Introduction
2. Results and Discussion
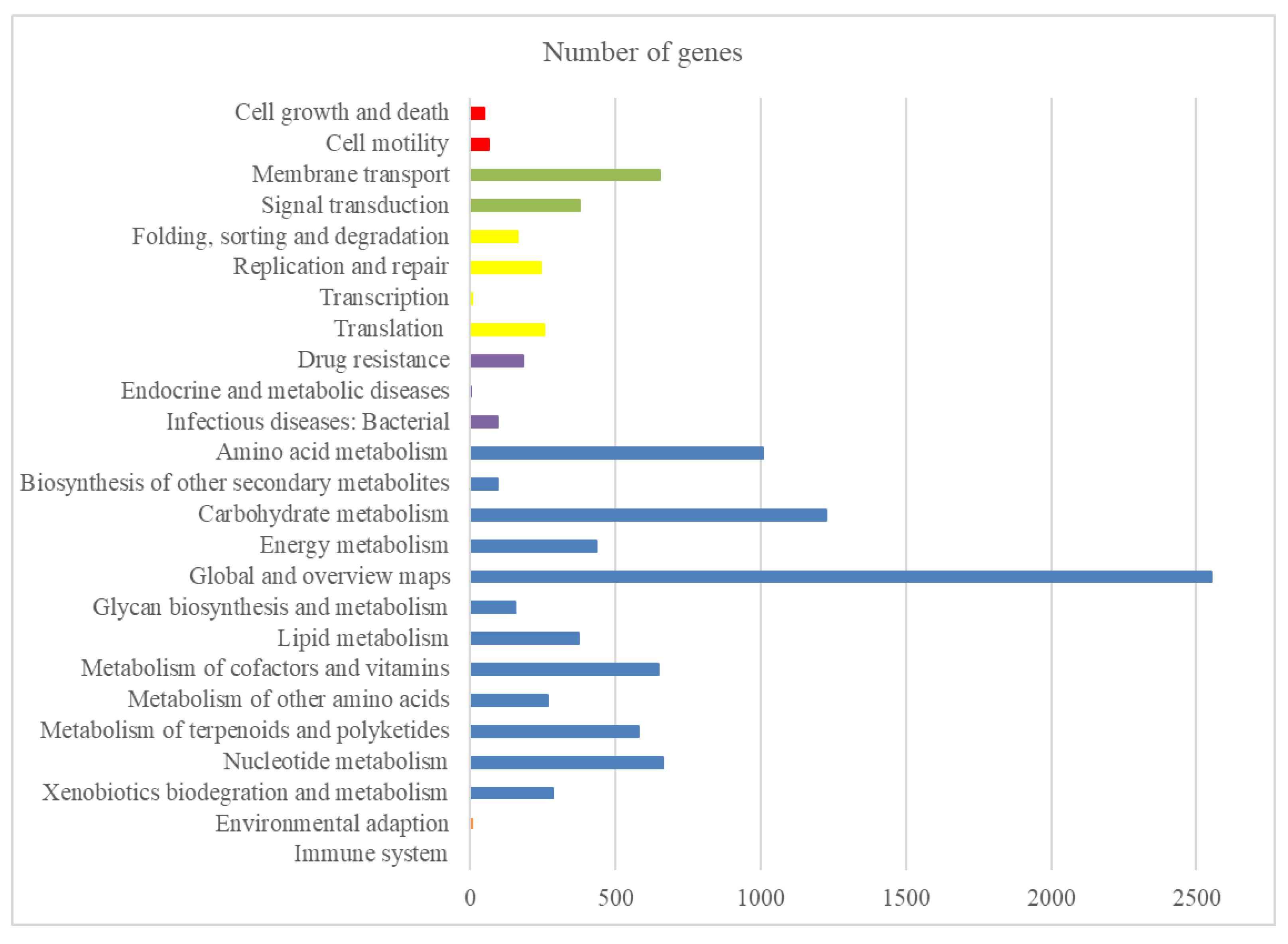
2.1. General Genome Features
2.2. Biosynthetic Gene Cluster Prediction
2.3. Secondary Metabolites Produced by the S. olivaceus Strain
3. Materials and Methods
3.1. Isolation of the S. olivaceus Strain
3.2. DNA Extraction and Whole Genome Sequencing
3.3. Fermentation Culture and Mass Spectrometric Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mohr, K.I. History of antibiotics research, in how to overcome the antibiotic crisis. Curr. Top. Microbiol. 2016, 398, 237–272. [Google Scholar]
- Ligon, B.L. Penicillin: Its discovery and early development. Semin. Pediatric Infect. Dis. 2004, 15, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Abed, N.; Couvreur, P. Nanocarriers for antibiotics: A promising solution to treat intracellular bacterial infections. Int. J. Antimicrob. A 2014, 43, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Raja, A.; Prabakarana, P. Actinomycetes and drug-an overview. Am. J. Drug Discov. Dev. 2011, 1, 75–84. [Google Scholar] [CrossRef]
- Guan, S.H.; Sattler, I.; Lin, W.H.; Guo, D.A.; Grabley, S. p-Aminoacetophenonic acids produced by a mangrove endophyte: Streptomyces griseus subsp. J. Nat. Prod. 2005, 68, 1198–1200. [Google Scholar] [CrossRef] [PubMed]
- de Lima Procópio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Braz. Raz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Maier, A.; Fiebig, H.H.; Lin, W.H.; Peschel, G.; Hertweck, C. Kandenols A–E, eudesmenes from an endophytic Streptomyces sp. of the mangrove tree Kandelia candel. J. Nat. Prod. 2012, 75, 2223–2227. [Google Scholar] [CrossRef] [PubMed]
- Ser, H.L.; Zainal, N.; Palanisamy, U.D.; Goh, B.H.; Yin, W.F.; Chan, K.G.; Lee, L.H. Streptomyces gilvigriseus sp. nov., a novel actinobacterium isolated from mangrove forest soil. Antonie Leeuwenhoek 2015, 107, 1369–1378. [Google Scholar] [CrossRef]
- Ser, H.L.; Tan, W.S.; Mutalib, N.S.A.; Yin, W.F.; Chan, K.G.; Goh, B.H.; Lee, L.H. Genome sequence of Streptomyces gilvigriseus MUSC 26T isolated from mangrove forest. Braz. J. Microbiol. 2018, 49, 207–209. [Google Scholar] [CrossRef]
- Hu, D.; Chen, Y.; Sun, C.; Jin, T.; Fan, G.; Liao, Q.; Mok, K.M.; Lee, S.M.-Y. Genome guided investigation of antibiotics producing actinomycetales strain isolated from a Macau mangrove ecosystem. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Hu, D.; Gao, C.; Sun, C.; Jin, T.; Fan, G.; Mok, K.M.; Lee, S.M.-Y. Genome-guided and mass spectrometry investigation of natural products produced by a potential new actinobacterial strain isolated from a mangrove ecosystem in Futian, Shenzhen, China. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Hu, D.; Sun, C.; Jin, T.; Fan, G.; Mok, K.M.; Li, K.; Lee, S.M.-Y. Exploring the potential of antibiotic production from rare actinobacteria by whole-genome sequencing and guided MS/MS analysis. Front. Microbiol. 2020, 11, 1540. [Google Scholar] [CrossRef]
- Asolkar, R.N.; Maskey, R.P.; Helmke, E.; Laatsch, H.; Chalcomycin, B. A new macrolide antibiotic from the marine isolate Streptomyces sp. B7064. J. Antibiot. 2002, 55, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Xu, M.J.; Zhao, Y.L.; Xu, J. A novel benzo [f][1, 7] naphthyridine produced by Streptomyces albogriseolus from mangrove sediments. Molecules 2010, 15, 9298–9307. [Google Scholar] [CrossRef] [PubMed]
- Thatoi, H.; Behera, B.C.; Mishra, R.R.; Dutta, S.K. Biodiversity and biotechnological potential of microorganisms from mangrove ecosystems: A review. Ann. Microbiol. 2013, 63, 1–19. [Google Scholar] [CrossRef]
- Kiranmayi, M.U.; Poda, S.; Vijayalakshmi, M. Production and optimization of L-asparaginase by an actinobacterium isolated from Nizampatnam mangrove ecosystem. J. Environ. Biol. 2014, 35, 799. [Google Scholar] [PubMed]
- Xu, D.B.; Ye, W.W.; Han, Y.; Deng, Z.X.; Hong, K. Natural products from mangrove actinomycetes. Mar. Drugs. 2014, 12, 2590–2613. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3 (2). Nature 2002, 417, 141. [Google Scholar] [CrossRef]
- Komatsu, M.; Komatsu, K.; Koiwai, H.; Yamada, Y.; Kozone, I.; Izumikawa, M.; Hashimoto, J.; Takagi, M.; Omura, S.; Shin-ya, K. Engineered Streptomyces avermitilis host for heterologous expression of biosynthetic gene cluster for secondary metabolites. ACS Synth. Biol. 2013, 2, 384–396. [Google Scholar] [CrossRef]
- Liu, C.X.; Zhang, J.; Wang, X.J.; Qian, P.T.; Wang, J.D.; Gao, Y.M.; Yan, Y.J.; Zhang, S.Z.; Xu, P.F.; Li, W.B.; et al. Antifungal activity of borrelidin produced by a Streptomyces strain isolated from soybean. J. Agric. Food. Chem. 2012, 60, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from bacterial and algal origin is a compatible solute in microalgae. Mar. Drugs. 2020, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Sauer, T.; Galinski, E.A. Bacterial milking: A novel bioprocess for production of compatible solutes. Biotechnol. Bioeng. 1998, 57, 306–313. [Google Scholar] [CrossRef]
- Dong, C.; Yao, Y. Isolation, characterization of melanin derived from Ophiocordyceps sinensis, an entomogenous fungus endemic to the Tibetan Plateau. J. Biosci. Bioeng. 2012, 113, 474–479. [Google Scholar] [CrossRef]
- Iwasa, T.; Yamamoto, H.; Shibata, M. Studies on validamycins, new antibiotics. I. Streptomyces hygroscopicus var. limoneus nov. var., validamycin-producing organism. Jpn. J. Antibiot. 1970, 23, 595–602. [Google Scholar] [CrossRef]
- Zhou, M.; Duan, Y.; Wang, J. Metconazole-Containing Pesticide Composition for Preventing and Controlling Fusarium Head Blight and Application Thereof. U.S. Patent 9,949,479 B2, 24 April 2018. [Google Scholar]
- Li, J.; Duan, Y.; Bian, C.; Pan, X.; Yao, C.; Wang, J.; Zhou, M. Effects of validamycin in controlling Fusarium head blight caused by Fusarium graminearum: Inhibition of DON biosynthesis and induction of host resistance. Pestic. Biochem. Phys. 2019, 153, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Suzuki-Nishimito, M.; Fukuchi, A.; Matsuura, K. Effective control of cabbage black rot by validamycin A and its effect on extracellular polysaccharide production of Xanthomonas campestris pv. campestris. J. Pestic. Sci. 2004, 29, 209–213. [Google Scholar] [CrossRef][Green Version]
- Ishikawa, R.; Shirouzu, K.; Nakashita, H.; Lee, H.Y.; Motoyama, T.; Yamaguchi, I.; Teraoka, T.; Arie, T. Foliar spray of validamycin A or validoxylamine A controls tomato fusarium wilt. Phytopathology 2005, 95, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Duan, Y.; Wang, J.; Xiu, Q.; Wang, J.; Hou, Y.; Song, X.; Zhou, M. Validamycin A induces broad-spectrum resistance involving in salicylic acid, and jasmonic acid/ethylene signaling pathways. Mol. Plant. Microbe Interact. 2020, 33, 1424–1437. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Lee, S.M.-Y.; Li, K.; Mok, K.M. Secondary Metabolite Production Potential of Mangrove-Derived Streptomyces olivaceus. Mar. Drugs 2021, 19, 332. https://doi.org/10.3390/md19060332
Hu D, Lee SM-Y, Li K, Mok KM. Secondary Metabolite Production Potential of Mangrove-Derived Streptomyces olivaceus. Marine Drugs. 2021; 19(6):332. https://doi.org/10.3390/md19060332
Chicago/Turabian StyleHu, Dini, Simon Ming-Yuen Lee, Kai Li, and Kai Meng Mok. 2021. "Secondary Metabolite Production Potential of Mangrove-Derived Streptomyces olivaceus" Marine Drugs 19, no. 6: 332. https://doi.org/10.3390/md19060332
APA StyleHu, D., Lee, S. M.-Y., Li, K., & Mok, K. M. (2021). Secondary Metabolite Production Potential of Mangrove-Derived Streptomyces olivaceus. Marine Drugs, 19(6), 332. https://doi.org/10.3390/md19060332