
Salmon (Salmo salar) Side Streams as a Bioresource to Obtain Potential Antioxidant Peptides after Applying Pressurized Liquid Extraction (PLE)



Abstract
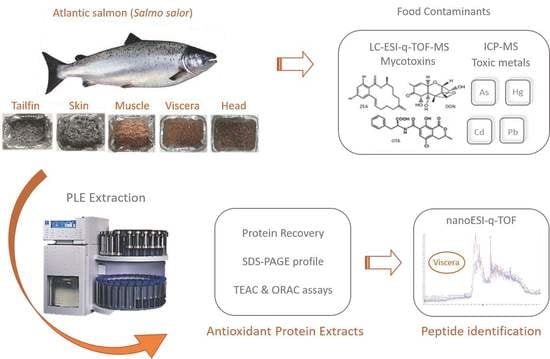
:
1. Introduction
2. Results and Discussion
2.1. Total Antioxidant Capacity
2.2. Protein Recovery Percentage
2.3. Protein Molecular Weight Distribution
2.4. Identification of Peptides in Viscera Extracts
2.5. Determination of Heavy Metals and Mycotoxins in Salmon Side Streams
3. Materials and Methods
3.1. Reagents
3.2. Raw Material and Sample Preparation
3.3. Pressurized Liquid Extraction (PLE) Process
3.4. Evaluation of Total Antioxidant Capacity
3.4.1. Trolox Equivalent Antioxidant Capacity Assay (TEAC)
3.4.2. Oxygen Radical Absorbance Capacity Assay (ORAC)
3.5. Determination of Protein Recovery
3.6. Molecular Weight Distribution of Protein Fragments
3.7. Identification of Peptides in Viscera Extracts
3.7.1. Sample Preparation
3.7.2. Mass Spectrometry Analysis
3.7.3. Data Analysis
3.8. Analysis of Heavy Metals in Salmon Side Stream Materials
3.9. Analysis of Mycotoxins in Salmon Side Stream Materials
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badiola, M.; Gartzia, I.; Basurko, O.C.; Mendiola, D. Land-based growth of Atlantic salmon (Salmo salar) and consumers’ acceptance. Aquac. Res. 2017, 48, 4666–4683. [Google Scholar] [CrossRef]
- Haq, M.; Ahmed, R.; Cho, Y.J.; Chun, B.S. Quality properties and bio-potentiality of edible oils from Atlantic salmon by-products extracted by supercritial carbon dioxide and conventional methods. Waste Biomass Valorization 2017, 8, 1953–1967. [Google Scholar] [CrossRef]
- Anonymous. The EU Fish Market 2020 Edition Is Now Online|Fisheries. Available online: https://ec.europa.eu/fisheries/press/eu-fish-market-2020-edition-now-online_en (accessed on 24 March 2021).
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef]
- He, S.; Franco, C.; Zhang, W. Characterisation of processing wastes of Atlantic salmon (Salmo salar) and Yellowtail kingfish (Seriola lalandi) harvested in Australia. Int. J. Food Sci. Technol. 2011, 46, 1898–1904. [Google Scholar] [CrossRef]
- Aspevik, T.; Thoresen, L.; Steinsholm, S.; Carlehög, M.; Kousoulaki, K. Sensory and chemical properties of protein hydrolysates based on mackerel (Scomber scombrus) and salmon (Salmo salar) side stream materials. J. Aquat. Food Prod. Technol. 2021, 30, 1–12. [Google Scholar] [CrossRef]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and bioactive properties of peptides derived from marine side streams. Mar. Drugs. 2021, 19, 71. [Google Scholar] [CrossRef]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from fish by-product protein hydrolysates and its functional properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. Rev. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Kim, J.-G.; Je, J.-Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef]
- Preventing Food Waste, Promoting Circular Economy. Available online: https://ec.europa.eu/commission/presscorner/detail/en/IP_19_2391 (accessed on 1 February 2021).
- Al Khawli, F.; Pateiro, M.; Domínguez, R.; Lorenzo, J.M.; Gullón, P.; Kousoulaki, K.; Ferrer, E.; Berrada, H.; Barba, F.J. Innovative green technologies of intensification for valorization of seafood and their by-products. Mar. Drugs 2019, 17, 689. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.F.; Ekorong, F.J.A.A.; Karkal, S.S.; Cathrine, M.S.B.; Kudre, T.G. Green and innovative techniques for recovery of valuable compounds from seafood by-products and discards: A review. Trends Food Sci. Technol. 2019, 85, 10–22. [Google Scholar] [CrossRef]
- Zia, S.; Khan, M.R.; Shabbir, M.A.; Aslam Maan, A.; Khan, M.K.I.; Nadeem, M.; Khalil, A.A.; Din, A.; Aadil, R.M. An inclusive overview of advanced thermal and nonthermal extraction techniques for bioactive compounds in food and food-related matrices. Food Rev. Int. 2020. [Google Scholar] [CrossRef]
- Alvarez-Rivera, G.; Bueno, M.; Ballesteros-Vivas, D.; Mendiola, J.A.; Ibañez, E. Pressurized liquid extraction. In Liquid-Phase Extraction; Elsevier: Amsterdam, The Netherlands, 2019; pp. 375–398. ISBN 9780128169117. [Google Scholar]
- Andreu, V.; Picó, Y. Pressurized liquid extraction of organic contaminants in environmental and food samples. TrAC-Trends Anal. Chem. 2019, 118, 709–721. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, J.; Collado, M.C.; Barba, F.J. accelerated solvent extraction and pulsed electric fields for valorization of rainbow trout (Oncorhynchus mykiss) and sole (Dover sole) by-products: Protein content, molecular weight distribution and antioxidant potential of the extracts. Mar. Drugs 2021, 19, 207. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, B.; Pallarés, N.; Barba, F.J.; Berrada, H. An integrated approach for the valorization of sea bass (Dicentrarchus labrax) side streams: Evaluation of contaminants and development of antioxidant protein extracts by pressurized liquid extraction. Foods 2021, 10, 546. [Google Scholar] [CrossRef]
- de la Fuente, B.; Pallarés, N.; Berrada, H.; Barba, F.J. Development of antioxidant protein extracts from gilthead sea bream (Sparus aurata) side streams assisted by Pressurized Liquid Extraction (PLE). Mar. Drugs 2021, 19, 199. [Google Scholar] [CrossRef] [PubMed]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kützing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef] [Green Version]
- Tolosa, J.; Barba, F.J.; Pallarés, N.; Ferrer, E. Mycotoxin identification and in silico toxicity assessment prediction in Atlantic salmon. Mar. Drugs 2020, 18, 629. [Google Scholar] [CrossRef]
- Gonçalves, R.A.; Schatzmayr, D.; Albalat, A.; Mackenzie, S. Mycotoxins in aquaculture: Feed and food. Rev. Aquac. 2020, 12, 145–175. [Google Scholar] [CrossRef]
- Kalantzi, I.; Pergantis, S.A.; Black, K.D.; Shimmield, T.M.; Papageorgiou, N.; Tsapakis, M.; Karakassis, I. Metals in tissues of seabass and seabream reared in sites with oxic and anoxic substrata and risk assessment for consumers. Food Chem. 2016, 194, 659–670. [Google Scholar] [CrossRef]
- Anonymous. Food and Agriculture Organization. Feed Production. Available online: http://www.fao.org/fishery/affris/species-profiles/atlantic-salmon/feed-production/en/ (accessed on 24 March 2021).
- Kandyliari, A.; Karavoltsos, S.; Sakellari, A.; Anastasiadis, P.; Asderis, M.; Papandroulakis, N.; Kapsofefalou, M. Trace metals in six fish by-products of two farmed fishes, the gilthead sea bream (Sparus aurata) and the meager (Argyrosomus regius): Interactions with the environment and feed. Hum. Ecol. Risk Assess. Int. J. 2020, 27, 1–21. [Google Scholar] [CrossRef]
- Sae-Leaw, T.; Karnjanapratum, S.; O’Callaghan, Y.C.; O’Keeffe, M.B.; FitzGerald, R.J.; O’Brien, N.M.; Benjakul, S. Purification and identification of antioxidant peptides from gelatin hydrolysate of seabass skin. J. Food Biochem. 2017, 41, e12350. [Google Scholar] [CrossRef]
- Firatligil-Durmus, E.; Evranuz, O. Response surface methodology for protein extraction optimization of red pepper seed (Capsicum frutescens). LWT-Food Sci. Technol. 2010, 43, 226–231. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM database of bioactive peptides: Current opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide identification in a salmon gelatin hydrolysate with antihypertensive, dipeptidyl peptidase IV inhibitory and antioxidant activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef]
- Khristoforova, N.K.; Tsygankov, V.Y.; Lukyanova, O.N.; Boyarova, M.D. High mercury bioaccumulation in Pacific salmons from the Sea of Okhotsk and the Bering Sea. Environ. Chem. Lett. 2018, 16, 575–579. [Google Scholar] [CrossRef]
- Winiarska-Mieczan, A.; Florek, M.; Kwiecień, M.; Kwiatkowska, K.; Krusiński, R. Cadmium and lead content in chosen commercial fishery products consumed in Poland and risk estimations on fish consumption. Biol. Trace Elem. Res. 2018, 182, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Nøstbakken, O.J.; Hove, H.T.; Duinker, A.; Lundebye, A.K.; Berntssen, M.H.G.; Hannisdal, R.; Lunestad, B.T.; Maage, A.; Madsen, L.; Torstensen, B.E.; et al. Contaminant levels in Norwegian farmed Atlantic salmon (Salmo salar) in the 13-year period from 1999 to 2011. Environ. Int. 2015, 74, 274–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmedo, P.; Pla, A.; Hernández, A.F.; Barbier, F.; Ayouni, L.; Gil, F. Determination of toxic elements (mercury, cadmium, lead, tin and arsenic) in fish and shellfish samples. Risk assessment for the consumers. Environ. Int. 2013, 59, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Setting Maximum Levels for Certain Contaminants in Food Stuffs (Text with EEA Relevance). 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32006R1881 (accessed on 24 March 2021).
- Bernhoft, A.; Høgåsen, H.R.; Rosenlund, G.; Ivanova, L.; Berntssen, M.H.G.; Alexander, J.; Eriksen, G.S.; Fæste, C.K. Tissue distribution and elimination of deoxynivalenol and ochratoxin A in dietary-exposed Atlantic salmon (Salmo salar). Food Addit. Contam. Part A 2017, 34, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Nácher-Mestre, J.; Ballester-Lozano, G.F.; Garlito, B.; Portolés, T.; Calduch-Giner, J.; Serrano, R.; Hernández, F.; Berntssen, M.H.G.; Pérez-Sánchez, J. Comprehensive overview of feed-to-fillet transfer of new and traditional contaminants in Atlantic salmon and gilthead sea bream fed plant-based diets. Aquac. Nutr. 2018, 24, 1782–1795. [Google Scholar] [CrossRef]
- Nácher-Mestre, J.; Serrano, R.; Beltrán, E.; Pérez-Sánchez, J.; Silva, J.; Karalazos, V.; Hernández, F.; Berntssen, M.H.G. Occurrence and potential transfer of mycotoxins in gilthead sea bream and Atlantic salmon by use of novel alternative feed ingredients. Chemosphere 2015, 128, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosa, J.; Barba, F.J.; Font, G.; Ferrer, E. Mycotoxin incidence in some fish products: QuEChERS methodology and liquid chromatography linear ion trap tandem mass spectrometry approach. Molecules 2019, 24, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, W. Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Shilov, I.V.; Seymourt, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The paragon algorithm, a next generation search engine that uses sequence temperature values sequence temperature values and feature probabilities to identify peptides from tandem mass spectra. Mol. Cell. Proteomics 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein of Origin of the Identified Peptide | Sequence | Obs MW | Obs m/z | Theor z |
|---|---|---|---|---|
| Collagen alpha-2(I) chain | GESGPTGNGGPVGA | 1155.52 | 578.77 | 2 |
| Collagen alpha-2(I) chain | GPAGPHGPPG | 842.4 | 422.21 | 2 |
| Collagen alpha-2(I) chain | SGETGSAGITGPAGPR | 1413.68 | 707.85 | 2 |
| Uncharacterized PE-PGRS family protein | GGNGGAGGAGGNGGAGGLGG | 1370.62 | 686.32 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPL | 819.45 | 410.73 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPLGP | 973.52 | 487.77 | 2 |
| Collagen alpha-3(V) chain | GPAGHPGPPG | 842.4 | 422.21 | 2 |
| Collagen alpha-1(I) chain | GETGPAGPAG | 812.4 | 407.21 | 2 |
| Collagen alpha-1(I) chain | GLPGSPGPAGEAGK | 1193.6 | 597.81 | 2 |
| Glycine-rich protein DOT1 | GGGGGHGGGAGGGGGGGPGG | 1292.58 | 647.3 | 2 |
| Collagen alpha-4(IV) chain | GPIGPLGPLGP | 973.52 | 487.77 | 2 |
| Probable heat shock protein ssa1 | PGGAPGGMPGGAP | 1021.47 | 511.74 | 2 |
| WAG22 antigen | PAGTAAGGAGGAGGAPGL | 1308.6 | 655.31 | 2 |
| Collagen alpha-1(I) chain | GETGPAGPAG | 812.4 | 407.21 | 2 |
| Histone H2A | AQGGVLPNIQ | 995.54 | 498.78 | 2 |
| 60 kDa heat shock protein, mitochondrial | VGGTSDVEVNEK | 1232.58 | 617.3 | 2 |
| Collagen alpha-1(XXII) chain | GYAKDGLPGIPGPQGET | 1655.76 | 828.89 | 2 |
| Filamin-A | VITPEEIVDPNVDEH | 1704.81 | 569.28 | 3 |
| Glycine-rich cell wall structural protein | GGGEGYGGGGANGGGY | 1285.6 | 643.81 | 2 |
| Fumarylacetoacetase | IGVAIGDQILDLSVIK | 1652.97 | 827.49 | 2 |
| Pulmonary surfactant-associated protein A | GPLGPPGGMPGH | 1072.53 | 537.27 | 2 |
| Collagen alpha-1(I) chain | GETGPAGPAG | 812.4 | 407.21 | 2 |
| Collagen alpha-4(IV) chain | GPPGLPGPPGPPGHKGF | 1607.77 | 804.89 | 2 |
| WAS/WASL-interacting protein family member | GGGGGGGGGGGGSGGNFGGGGPP | 1586.64 | 794.33 | 2 |
| Adenylate cyclase type 10 | GRVNIQDLQKNKFLMRANT | 2245.16 | 749.4 | 3 |
| Forkhead box protein K1 | QPPPGPPPPPP | 1076.55 | 539.28 | 2 |
| Exocyst complex component SEC5 | ALMILIVVHSECFR | 1629.91 | 815.96 | 2 |
| tRNA dimethylallyltransferase | EAARDGWPAL | 1084.53 | 543.27 | 2 |
| Orotidine 5’-phosphate decarboxylase | RPAGAEAGDQK | 1098.54 | 550.28 | 2 |
| Homogentisate 1,2-dioxygenase | GPIGSNGLANPR | 1152.52 | 577.27 | 2 |
| Wolframin | NTAPLGPSCPQPPPAP | 1508.68 | 755.35 | 2 |
| Keratin, type II cytoskeletal I | TALGGAAGGMGGGGGMGGGM | 1538.63 | 770.32 | 2 |
| Integrin-linked-kinase-associated- serine/ | ||||
| threonine phosphatase 2C | GLPPAGSGNSGSLATSGS | 1515.64 | 758.83 | 2 |
| Collagen alpha-4(IV) chain | ACAGMIGPPGPQGFP | 1399.63 | 467.55 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPLGP | 973.52 | 487.77 | 2 |
| Fatty acid-binding protein, liver | AIGLPDDLIQK | 1181.67 | 591.84 | 2 |
| Actin-related protein 3 | VIDSGDGVTH | 998.47 | 500.24 | 2 |
| Collagen alpha-1(I) chain | GAPGPVGPAGKGETGPAGPAGPAG | 1925.86 | 963.94 | 2 |
| Chaperone protein DnaK | QAGEGGAGAGAGAAG | 1100.52 | 551.27 | 2 |
| Collagen alpha-2(V) chain | GNPGPLGPIGP | 974.52 | 488.27 | 2 |
| Collagen alpha-1(XVIII) chain | LPGPPGPPGPPGPRGYPG | 1665.79 | 833.9 | 2 |
| POTE ankyrin domain family member E | VMDSGDGVTH | 1016.43 | 509.22 | 2 |
| Uncharacterized protein SE_1560 | GPLVLVDTDDL | 1155.61 | 578.81 | 2 |
| Serum albumin 1 | AIQPDTEFTPPELDASS | 1816.84 | 909.43 | 2 |
| Phosphoenolpyruvate guanylyltransferase | SLAMLNDVLVAL | 1257.73 | 629.87 | 2 |
| Collagen alpha-1(I) chain | AGPPGADGQPGAK | 1164.55 | 583.28 | 2 |
| DNA (cytosine-5)-methyltransferase 3A | DPASPNVATTP | 1068.5 | 535.26 | 2 |
| Protein Shroom4 | SQAPESHESRTGL | 1397.61 | 699.81 | 2 |
| Ataxin-2 homolog | PAGGGPQPAFTPP | 1192.56 | 597.29 | 2 |
| Magnesium-chelatase 38 kDa subunit | QSGENVVERDGL | 1301.6 | 651.81 | 2 |
| Uroporphyrinogen decarboxylase | DVAVQGNLDPL | 1139.61 | 570.81 | 2 |
| Collagen alpha-1(I) chain | AGAQGAPGPAGPA | 1021.47 | 511.74 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPLGP | 973.53 | 487.77 | 2 |
| Actin-related protein 3 | DSGDGVTH | 786.32 | 394.17 | 2 |
| tRNA-N6-adenosine-threonylcarbamoy | ||||
| ltransferase | LSLVVSGGHTELVL | 1422.69 | 712.35 | 2 |
| Calpain-12 | AGTGAGGPQ | 714.2 | 358.11 | 2 |
| Collagen alpha-1(XVII) chain | QNLVGPPGPPGPPGVSGD | 1623.77 | 812.89 | 2 |
| Fumarylacetoacetase | IGVAIGDQILDLSVIK | 1652.97 | 552 | 3 |
| Probable aquaporin PIP2-6 | DINAGGGACASVGLL | 1316.67 | 659.34 | 2 |
| 60 kDa chaperonin | AAVEEGIVAGGGTAF | 1347.58 | 674.8 | 2 |
| Arginine kinase | KGDRFLEAAGVNKLWPE | 1928.92 | 965.47 | 2 |
| Collagen alpha-2(I) chain | GETGSAGITGPAGPR | 1326.65 | 664.33 | 2 |
| Cytoplasmic dynein 1 light intermediate chain 1 | TGSPGGPGVSGGSPAGGAG | 1425.64 | 713.83 | 2 |
| Collagen alpha-2(I) chain | RGDGGPPGVTGFPGAA | 1411.63 | 706.82 | 2 |
| Collagen alpha-1(I) chain | AKGDTGAPGAPGSQGAP | 1437.68 | 719.85 | 2 |
| Zinc finger protein 831 | ESEGEGGPGPGPGVAGAEP | 1649.78 | 550.93 | 3 |
| Collagen alpha-2(IV) chain | PGEKGDAGLPGLSGK | 1363.64 | 682.83 | 2 |
| Collagen alpha-2(I) chain | GPTGNGGPVGA | 882.42 | 442.22 | 2 |
| Translation initiation factor IF-2 | GGGGGAPGRPGGGGGGGGAP | 1405.65 | 703.83 | 2 |
| Collagen alpha-2(I) chain | GPAGPHGPP | 785.38 | 393.7 | 2 |
| Serine/threonine-protein kinase ATG1 | ESNMFVSEYL | 1217.56 | 609.79 | 2 |
| ATP-dependent RNA helicase DBP7 | REGKWDIHATT | 1312.67 | 657.34 | 2 |
| Nucleoside diphosphate kinase B | ETNPADSKPGSI | 1214.58 | 608.3 | 2 |
| Glucosyl-3-phosphoglycerate synthase | VAGDLAGGRAPGALP | 1320.64 | 661.33 | 2 |
| Collagen alpha-6(IV) chain | VGPLGPSG | 682.33 | 342.17 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPLGP | 973.53 | 487.77 | 2 |
| 5’-3’ exoribonuclease 2 | NNGGGGGGYGGQP | 1090.51 | 546.26 | 2 |
| PE-PGRS family protein PE_PGRS30 | NGGAAGLIGNGGAGGAGGAGGAG | 1639.72 | 820.87 | 2 |
| Protein FAM81B | DTNVNKSASPTATAEEQPVEP | 2184.09 | 1093.05 | 2 |
| E3 ubiquitin-protein ligase TOPORS | DQGLFMGPSTSGAAANR | 1679.7 | 560.91 | 3 |
| (R)-2-hydroxyglutaryl-CoA-dehydratase | ||||
| activating ATPase | GIADKQMSELSCHA | 1488.7 | 745.36 | 2 |
| Uncharacterized TPR repeat-containing | ||||
| protein At1g05150 | DALGLELNADE | 1158.57 | 580.29 | 2 |
| Collagen alpha-1(I) chain | DGNPGLPGPPGPPGPPG | 1492.69 | 747.35 | 2 |
| Golgin subfamily A member 6A | GNHEGHG | 706.28 | 354.15 | 2 |
| Collagen alpha-2(IV) chain | EVLGAQPGTRGDAGLPGQPG | 1875.93 | 626.32 | 3 |
| MTOR-associated protein MEAK7 | DVDGLFDTLSGSSSSAAAKNGK | 2126.05 | 1064.03 | 2 |
| Transforming protein Maf | GSAAAVVSAVIAAA | 1156.53 | 579.27 | 2 |
| Glycine dehydrogenase (decarboxylating) | PGAMGADIAIG | 971.4 | 486.71 | 2 |
| L-lactate dehydrogenase A-like 6B | SVADLTESILK | 1174.65 | 392.56 | 3 |
| CTP synthase | PDGKLVEICEVTGHPF | 1739.83 | 870.92 | 2 |
| Collagen alpha-3(V) chain | GIPGPLGPL | 819.45 | 410.73 | 2 |
| T-related protein | VSGGGGGGGAGGGAGSGSPQ | 1429.68 | 715.85 | 2 |
| Glyceraldehyde-3-phosphate dehydrogenase 1 | TVDGPSGK | 759.37 | 380.69 | 2 |
| UDP-3-O-acylglucosamine N-acyltransferase | ADGFGFAPDFGPQGGEW | 1753.78 | 877.9 | 2 |
| Protein prickle | GGGAGGSSGGPGGADAAAAPAAGQ | 1767.76 | 884.89 | 2 |
| Histone H2A | AQGGVLPNIQ | 995.54 | 498.78 | 2 |
| Putative cuticle collagen 155 | GPSGPNGNPGAPGAPGQ | 1430.71 | 716.36 | 2 |
| BTB/POZ domain and ankyrin repeat- | ||||
| containing protein NH5.1 | GGAGGGGGAP | 656.34 | 329.18 | 2 |
| PE-PGRS family protein PE_PGRS5 | GAGGKGGNGGTGGAGGPGG | 1341.64 | 671.83 | 2 |
| Collagen alpha-5(IV) chain | PGIPGIGLPGPPGPKGFPGIP | 1947 | 974.51 | 2 |
| Glutamate dehydrogenase 1, mitochondrial | IGPGIDVPAPDMSTGE | 1554.73 | 778.37 | 2 |
| Collagen alpha-2(IV) chain | SGPSGIPGLPGPKGEPGY | 1665.76 | 833.89 | 2 |
| Collagen alpha-1(I) chain | GLPGSPGPAGEAGK | 1193.6 | 597.81 | 2 |
| TRPM8 channel-associated factor homolog | SEAVQTNLVPFFEAWGWPI | 2190.1 | 1096.06 | 2 |
| Collagen alpha-4(IV) chain | GPPGIPGPNGEDGLPGLP | 1639.76 | 820.89 | 2 |
| Elastin | VPGAVPGGVP | 848.44 | 425.23 | 2 |
| Multidrug resistance protein PE_PGR46 | IMVVVQPFVLVAI | 1426.82 | 714.42 | 2 |
| Uncharacterized PE-PGRS family protein PE_PGRS46 | GDGAPGGDGGAGPLLIGNG | 1550.68 | 776.35 | 2 |
| POTE ankyrin domain family member E | SGDGVTH | 671.29 | 336.65 | 2 |
| Actin-related protein 3 | SEVVDEVIQN | 1130.54 | 566.28 | 2 |
| Actin-related protein 3 | SGDGVTH | 671.29 | 336.65 | 2 |
| Protein Wiz | GPERLPGPAPRENIEGGAE | 1944.94 | 973.48 | 2 |
| DNA-directed RNA polymerase subunit beta | GKPIPESGLPE | 1122.53 | 562.27 | 2 |
| Histone H2A | AQGGVLPNIQ | 995.54 | 498.78 | 2 |
| Ribulose bisphosphate carboxylase/oxygenase activase 2, chloroplastic | TLMNIADNPTNVQLP | 1639.72 | 820.87 | 2 |
| FT-interacting protein 1 | PEVFVKAQVGNQILK | 1668.86 | 835.43 | 2 |
| Collagen alpha-2(I) chain | GAVGPVGPVG | 808.44 | 405.23 | 2 |
| Collagen alpha-2(I) chain | GPIGPPGNPGA | 932.47 | 467.24 | 2 |
| Polyribonucleotide nucleotidyltransferase | TEAVVAEGLEAAKP | 1383.75 | 692.88 | 2 |
| Putative cuticle collagen 145 | EGPAGPAGPAGPDGQPGA | 1501.64 | 751.83 | 2 |
| Contactin-3 | VSGGGGSRSELVITWDPVP | 1911.97 | 956.99 | 2 |
| Collagen alpha-1(III) chain | EPGQAGPAGPPGPPG | 1285.6 | 1286.61 | 1 |
| Collagen alpha-2(I) chain | SIGEPGPIGIAG | 1066.51 | 534.26 | 2 |
| Collagen alpha-2(I) chain isoform X3 | GDPGPGGPQGEPGAVGPAGITGDKGPSGES | 2601.2 | 868.08 | 3 |
| Uncharacterized protein | DIKPVTEIQQNGNDFVITSK | 2245.16 | 749.4 | 3 |
| Calmodulin | IDQLTEEQIAEF | 1434.65 | 718.33 | 2 |
| Mitochondrial fission regulator | HLSLPRFFPSRTGE | 1643.18 | 548.73 | 3 |
| Collagen, type V, alpha 3a | LIDVLRVLELSEDMEGVSV | 2114.92 | 1058.47 | 2 |
| Si:dkey-237h12.3 | ELDASNMGGWSLDK | 1521.81 | 761.91 | 2 |
| Uncharacterized protein Salmo truta | AGAEGFDDIK | 1021.47 | 511.74 | 2 |
| Fatty acid-binding protein, liver | AIGLPDDLIQK | 1181.67 | 591.84 | 2 |
| Uncharacterized protein Sinocyclocheilus | ||||
| anshuiensis | DVFRDGFTMDT | 1302.61 | 652.31 | 2 |
| Collagen alpha-4(IV) chain | GSSPIGPPGSPGSPGASGQ | 1592.74 | 797.38 | 2 |
| Mucin-5AC-like | GGPTSGSEGGDNESIK | 1490.65 | 746.33 | 2 |
| D-dopachrome decarboxylase | MIVVVKPGLPMLM | 1426.82 | 714.42 | 2 |
| Uncharacterized protein OS = Echeneis naucrates | PKPLPFFGTMLSYR | 1653 | 827.51 | 2 |
| Fumarylacetoacetase | IGVAIGDQILDLSVIK | 1652.97 | 827.49 | 2 |
| Protein of Origin of the Identified Peptide | Sequence | Obs MW | Obs m/z | Theor z |
|---|---|---|---|---|
| Adenosylhomocysteinase | GVSEETTTGVH | 1115.51 | 558.76 | 2 |
| Hemoglobin subunit alpha | AIHFPADFTPEVH | 1479.71 | 494.24 | 3 |
| Forkhead box protein K1 | PQPPPGPPPPP | 1076.57 | 539.29 | 2 |
| 40S ribosomal protein | ADGYEPPIQET | 1218.54 | 610.28 | 2 |
| WW domain-binding protein 11 | PGPPPGPPPP | 908.48 | 455.24 | 2 |
| Filamin-A | VITPEEIVDPNVDEH | 1704.81 | 569.28 | 3 |
| Collagen alpha-1(X) chain | ISVPGKPGPQ | 978.47 | 490.24 | 2 |
| Fatty acid-binding protein 10-A, liver basic | AQENYEEFLR | 1297.59 | 649.8 | 2 |
| Methionine import ATP-binding protein MetN | IDEIGGQHVGSLVLGVP | 1688.81 | 845.41 | 2 |
| Probable tRNA pseudouridine synthase | ENNVDFVNRKIKEGEAMVSGPI | 2445.24 | 816.09 | 3 |
| Mediator of RNA polymerase II transcription subunit 30 | LAASGMAPGPFAGPQ | 1370.71 | 686.36 | 2 |
| 1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)- methylideneamino] imidazole-4- | ||||
| carboxamide isomerase | HWVDQGGKRLHL | 1444.89 | 723.45 | 2 |
| Quinolinate synthase A | EGADEVHVDPGI | 1236.58 | 619.29 | 2 |
| 40S ribosomal protein S17 | DQEIIEVDPDT | 1272.58 | 637.3 | 2 |
| Uncharacterized PE-PGRS family protein | NGGNGGDGGNGGDGGNGAP | 1627.66 | 814.84 | 2 |
| PE_PGRS54 | GPPPPGPPPEVVI | 1251.65 | 626.83 | 2 |
| Prostaglandin reductase 1 | LVGAGNNGGDALLAAAELAR | 1851.87 | 926.94 | 2 |
| NAD(P)H-hydrate epimerase | VLRFFMATTQYR | 1531.9 | 766.96 | 2 |
| Cysteine--tRNA ligase | DSGDGVTH | 786.32 | 787.33 | 1 |
| Actin, cytoplasmic 1 | LDRMKNSCIVCNIGH | 1701.92 | 851.97 | 2 |
| Putative adenosylhomocysteinase 3 | SSSSILVVIATL | 1188.79 | 595.4 | 2 |
| Spore membrane assembly protein 2 | IPAINVNDSVT | 1141.6 | 571.81 | 2 |
| Adenosylhomocysteinase | IHFPADFTPEVH | 1408.68 | 470.57 | 3 |
| Hemoglobin subunit alpha | VFASYPQPLG | 1077.53 | 539.77 | 2 |
| Uncharacterized protein y4iR | ||||
| 2-C-methyl-D-erythritol 4-phosphate- | ||||
| cytidylyltransferase | LQSVIAVVPAAGV | 1222.84 | 612.43 | 2 |
| Zinc finger C2HC domain-containing | ||||
| protein 1A | NQVIKDGGPLPPPPPP | 1621.8 | 811.91 | 2 |
| Trichodiene synthase | VSEGITLNQALE | 1272.58 | 637.3 | 2 |
| 60 kDa heat shock protein, mitochondrial | GTSDVEVNEK | 1076.5 | 539.25 | 2 |
| pH-response regulator protein palF/RIM8 | PIRITHLTVAL | 1232.8 | 617.41 | 2 |
| Leucine-rich repeat-containing protein 56 | LEQLEVLDLEGNS | 1457.65 | 729.83 | 2 |
| Tungsten-containing formylmethanofuran- | ||||
| dehydrogenase 2 subunit C | DVDVRVGGEMKAG | 1331.66 | 666.84 | 2 |
| Cyclic pyranopterin monophosphate synthase | NTNGEANMVDVSMKQ | 1636.8 | 819.41 | 2 |
| Acetylcholinesterase | FRHPRPAEKWTGV | 1579.88 | 790.95 | 2 |
| Uncharacterized PE-PGRS family protein PE | NGGNGGIGGP | 798.43 | 400.22 | 2 |
| Sulfocyanin | SPSASSSTGTSTGP | 1222.59 | 612.3 | 2 |
| Actin, cytoplasmic | VMDSGDGVTH | 1016.42 | 509.22 | 2 |
| 40S ribosomal protein S3a | GEGGGSSAAKPSG | 1060.47 | 531.24 | 2 |
| DNA repair protein crb2 | DSLYDRLLARKGPLFGK | 1948.23 | 975.12 | 2 |
| Argininosuccinate synthase | IEGGRLEDPSFVPP | 1511.82 | 756.92 | 2 |
| Collagen alpha-2(I) chain | GAVGPVGPVG | 808.44 | 405.23 | 2 |
| Thiazole synthase | GVLLNTAVSGAKDP | 1340.73 | 671.37 | 2 |
| Histone-lysine N-methyltransferase 2D | PLSPPPEDSPLSPPP | 1525.79 | 509.61 | 3 |
| Probable transcriptional regulatory | ||||
| protein Ecaj_0351 | NFDSLFNIAI | 1152.59 | 577.3 | 2 |
| 50S ribosomal protein L29 | HAKKAELFELRVK | 1567.85 | 784.93 | 2 |
| Forkhead box protein K1 | QPPPGPPPPPP | 1076.57 | 539.29 | 2 |
| Probable GPI-anchored adhesin-like | ||||
| protein PGA32 | ATAAGTEVQGFTPI | 1361.6 | 681.81 | 2 |
| Replicase polyprotein 1ab | MAKMGKYGLGFK | 1329.9 | 665.96 | 2 |
| Stonin-2 | VVDGGSQDHS | 999.35 | 500.68 | 2 |
| SLAIN motif-containing protein | AGGGGPEPGGAGTPPGAAAAP | 1615.84 | 808.93 | 2 |
| Structural maintenance of chromosomes | ||||
| protein 4 | EIQNSILNVGGPQ | 1367.67 | 684.84 | 2 |
| Golgin-84 | TPEIH | 595.3 | 298.66 | 2 |
| Keratin, type II cytoskeletal 5 | LGGGAGFGGGYGGP | 1122.54 | 562.28 | 2 |
| Translation initiation factor IF-2 | VEEGLTSDEPDLE | 1431.6 | 716.81 | 2 |
| Genome polyprotein | IDLSANAAGSDPP | 1226.61 | 614.31 | 2 |
| Collagen alpha-1(X) chain | ISVPGKPGPQ | 978.47 | 490.24 | 2 |
| Transcription-associated protein 1 | VASVQPYAMPP | 1158.53 | 580.27 | 2 |
| MAM and LDL-receptor class A domain- | ||||
| containing- protein 2 | LDDSPCPPE | 971.37 | 972.38 | 1 |
| Coiled-coil domain-containing protein CG32809 | SSSKKKRKGRE | 1289.85 | 645.93 | 2 |
| Large tegument protein deneddylase | SVPAPPTLPP | 974.52 | 488.27 | 2 |
| Adenylyl cyclase-associated protein | GPPPPGPPPPP | 1004.53 | 503.27 | 2 |
| Neuroblast differentiation-associated protein AHNAK | VDIEGPDVDIEGSGG | 1457.65 | 729.83 | 2 |
| Mediator of RNA polymerase II transcription subunit 28 | QPPGPPPPPPP | 1076.56 | 539.29 | 2 |
| Protein S100 | DLDANSDGSVDFQ | 1381.56 | 691.79 | 2 |
| LisH domain-containing protein | VISYALDLIEVKHDSARVH | 2164.32 | 1083.16 | 2 |
| Adenylyl cyclase-associated protein | DGDYTEIPVPEQ | 1361.59 | 681.8 | 2 |
| Guanylate cyclase domain-containing protein | LISPGDAL | 784.35 | 393.18 | 2 |
| Insulin receptor substrate 2 | VCGGSGPG | 632.26 | 317.14 | 2 |
| Sequence_Modification | Sequence in BIOPEP-UWM Database | Identity of Sequences with Antioxidant Potential |
|---|---|---|
| GPAGPHGPPG | PHG | ID 8026 synthetic peptide |
| GPP | ID 8987 | |
| GPAGHPGPPG | GPP | ID 8987 |
| GYAKDGLPGIPGPQGET | KD | ID 8134 peptide from dried bonito |
| GGGEGYGGGGANGGGY | GGE | ID 8114 peptide from sardinella byproducts |
| GPLGPPGGMPGH | GPP | ID 8987 |
| GPPGLPGPPGPPGHKGF_ Carbamyl(K)@15 | GPP | ID 8987 |
| GGGGGGGGGGGGSGGNFGGGGPP | GPP | ID 8987 |
| QPPPGPPPPPP | GPP | ID 8987 |
| TALGGAAGGMGGGGGMGGGM_ Oxidation(M)@20 | GAA | ID 8983 |
| ACAGMIGPPGPQGFP_ Deamidated(Q)@12 | GPP | ID 8987 |
| ACA | ID 10038 | |
| QAGEGGAGAGAGAAG | GAA | ID 8983 |
| LPGPPGPPGPPGPRGYPG | GPP | ID 8987 |
| AIQPDTEFTPPELDASS | EL | ID 7888 |
| PEL | ID 8139 synthetic peptide | |
| GPP | ID 8987 | |
| LSLVVSGGHTELVL | EL | ID 7888 |
| QNLVGPPGPPGPPGVSGD_ Gln->pyro-Glu@N-term | GPP | ID 8987 |
| DINAGGGACASVGLL | ACA | ID 10038 |
| KGDRFLEAAGVNKLWPE | LW | ID 8462 peptide from marine bivalve |
| RGDGGPPGVTGFPGAA | GAA | ID 8983 |
| GPP | ID 8987 | |
| GPAGPHGPP | PHG | ID 8026 |
| GPP | ID 8987 | |
| ETNPADSKPGSI | KP | ID 8218 |
| NGGAAGLIGNGGAGGAGGAGGAG | GAA | ID 8983 |
| DQGLFMGPSTSGAAANR_ Deamidated(N)@16 | GAA | ID 8983 |
| GIADKQMSELSCHA | EL | ID 7888 |
| DALGLELNADE | EL | ID 7888 |
| DGNPGLPGPPGPPGPPG_ Pro->pyro-Glu(P)@16 | GPP | ID 8987 |
| SVADLTESILK | LK | ID 8217 |
| ADGFGFAPDFGPQGGEW | GGE | ID 8114 peptide from sardinella by-products |
| ADGF | ID 9328 | |
| PGIPGIGLPGPPGPKGFPGIP_ Delta:H(2)C(2)(K)@15 | GPP | ID 8987 |
| SEAVQTNLVPFFEAWGWPI | WG | ID 9082 |
| EAVQ | ID 9881 | |
| GPPGIPGPNGEDGLPGLP | GPP | ID 8987 |
| GKPIPESGLPE | KP | ID 8218 |
| PEVFVKAQVGNQILK | LK | ID 8217 |
| GPIGPPGNPGA | GPP | ID 8987 |
| TEAVVAEGLEAAKP | KP | ID 8218 |
| VSGGGGSRSELVITWDPVP | EL | ID 7888 |
| TW | ID 8459 peptide from marine bivalve | |
| EPGQAGPAGPPGPPG_ Deamidated(Q)@4 | GPP | ID 8987 |
| DIKPVTEIQQNGNDFVITSK | KP | ID 8218 |
| HLSLPRFFPSRTGE | HL | ID 3317 |
| LIDVLRVLELSEDMEGVSV | EL | ID 7888 |
| ELDASNMGGWSLDK | EL | ID 7888 |
| GGPTSGSEGGDNESIK | GPP | ID 8987 |
| MIVVVKPGLPMLM | KP | ID 8218 |
| VKP | ID 8434 peptide from jellyfish | |
| PKPLPFFGTMLSYR | LPM | ID 9360 |
| IGVAIGDQILDLSVIK | KP | ID 8218 |
| Sequence_Modification | Sequence in BIOPEP-UWM Database | Identity of Sequences with Antioxidant Potential |
|---|---|---|
| AIHFPADFTPEVH | ADF | ID 7868 peptide from Okara protein |
| PQPPPGPPPPP | GPP | ID 8987 |
| PGPPPGPPPP | GPP | ID 8987 |
| ISVPGKPGPQ | KP | ID 8218 |
| HWVDQGGKRLHL | LH | ID 3305 |
| HL | ID 3317 | |
| LHL | ID 7995 synthetic peptide | |
| GPPPPGPPPEVVI | GPP | ID 8987 |
| LVGAGNNGGDALLAAAELAR | EL | ID 7888 |
| IHFPADFTPEVH | ADF | ID 7868 peptide from Okara protein |
| NQVIKDGGPLPPPPPP | KD | ID 8134 peptide from dried bonito |
| PIRITHLTVAL | HL | ID 3317 |
| IR | ID 8215 | |
| DVDVRVGGEMKAG | GGE | ID 8114 peptide from sardinella by-products |
| GEGGGSSAAKPSG | KP | ID 8217 |
| GVLLNTAVSGAKDP | KD | ID 8134 peptide from dried bonito |
| HAKKAELFELRVK | EL | ID 7888 |
| QPPPGPPPPPP | GPP | ID 8987 |
| AGGGGPEPGGAGTPPGAAAAP | GAA | ID 8983 |
| Salmon Side Streams | Heavy Metals (µg/g of Wet Weight) | |||
|---|---|---|---|---|
| As | Hg | Cd | Pb | |
| Muscle | 0.5413 ± 0.0068 | 0.0238 ± 0.0005 | 0.0004 ± 0.0001 | 0.0269 ± 0.0002 |
| Head | 0.6922 ± 0.0072 | 0.0157 ± 0.0005 | 0.0011 ± 0.0001 | 0.0190 ± 0.0001 |
| Viscera | 0.4617 ± 0.0055 | 0.0095 ± 0.0002 | 0.0044 ± 0.0002 | 0.0071 ± 0.0001 |
| Skin | 0.4504 ± 0.0032 | 0.0077 ± 0.0003 | 0.0019 ± 0.0001 | 0.0247 ± 0.0001 |
| Tailfin | 0.4186 ± 0.0054 | 0.0408 ± 0.0015 | 0.0104 ± 0.0003 | 0.0859 ± 0.0016 |
| (Legislation *) | <13.5 | <0.50 | <0.05 | <0.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Fuente, B.; Pallarés, N.; Berrada, H.; Barba, F.J. Salmon (Salmo salar) Side Streams as a Bioresource to Obtain Potential Antioxidant Peptides after Applying Pressurized Liquid Extraction (PLE). Mar. Drugs 2021, 19, 323. https://doi.org/10.3390/md19060323
de la Fuente B, Pallarés N, Berrada H, Barba FJ. Salmon (Salmo salar) Side Streams as a Bioresource to Obtain Potential Antioxidant Peptides after Applying Pressurized Liquid Extraction (PLE). Marine Drugs. 2021; 19(6):323. https://doi.org/10.3390/md19060323
Chicago/Turabian Stylede la Fuente, Beatriz, Noelia Pallarés, Houda Berrada, and Francisco J. Barba. 2021. "Salmon (Salmo salar) Side Streams as a Bioresource to Obtain Potential Antioxidant Peptides after Applying Pressurized Liquid Extraction (PLE)" Marine Drugs 19, no. 6: 323. https://doi.org/10.3390/md19060323
APA Stylede la Fuente, B., Pallarés, N., Berrada, H., & Barba, F. J. (2021). Salmon (Salmo salar) Side Streams as a Bioresource to Obtain Potential Antioxidant Peptides after Applying Pressurized Liquid Extraction (PLE). Marine Drugs, 19(6), 323. https://doi.org/10.3390/md19060323