
Protective Effects of Fucoxanthin on Hydrogen Peroxide-Induced Calcification of Heart Valve Interstitial Cells

 , ,
, ,  ,
,  ,
, 
Abstract
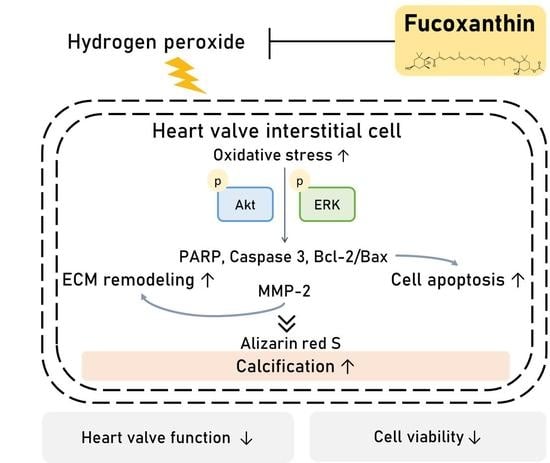
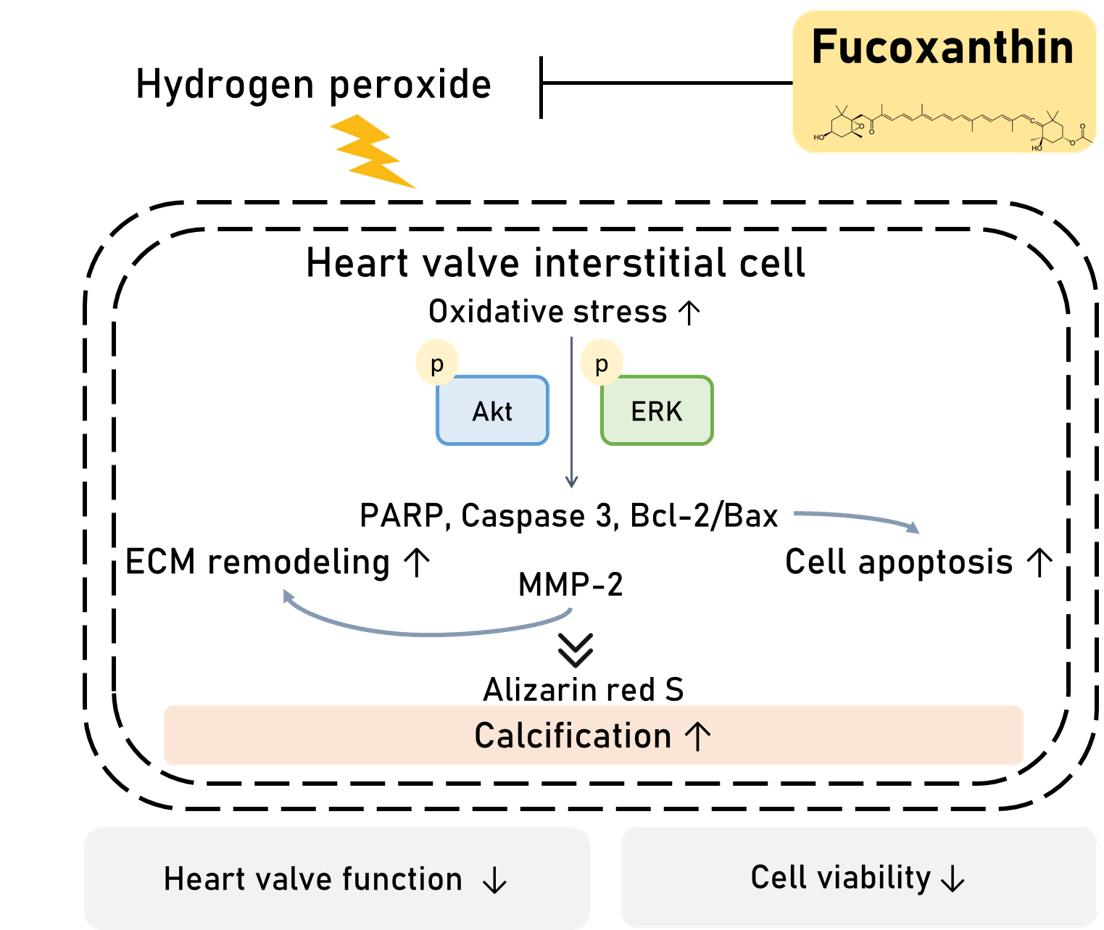
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
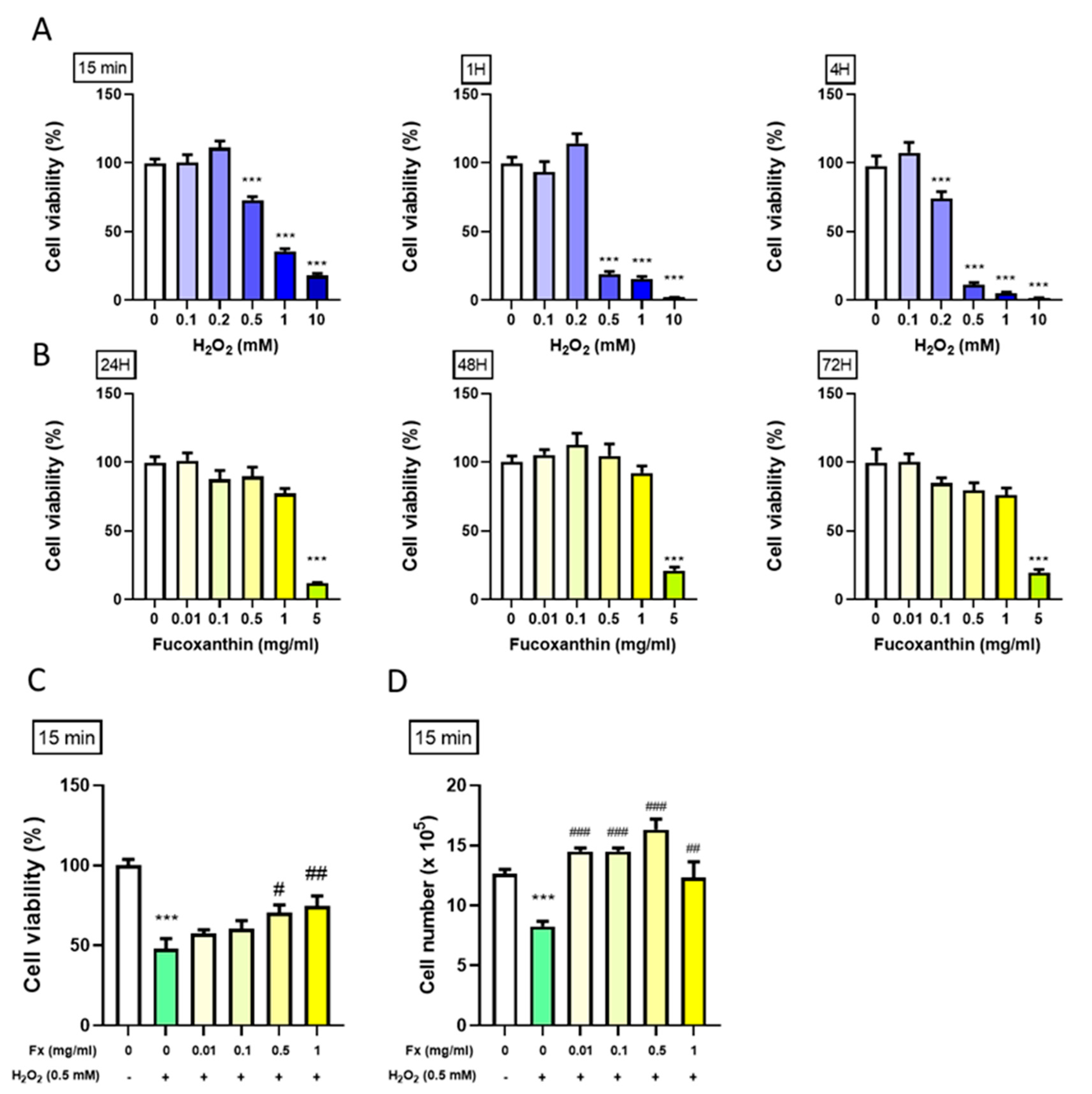
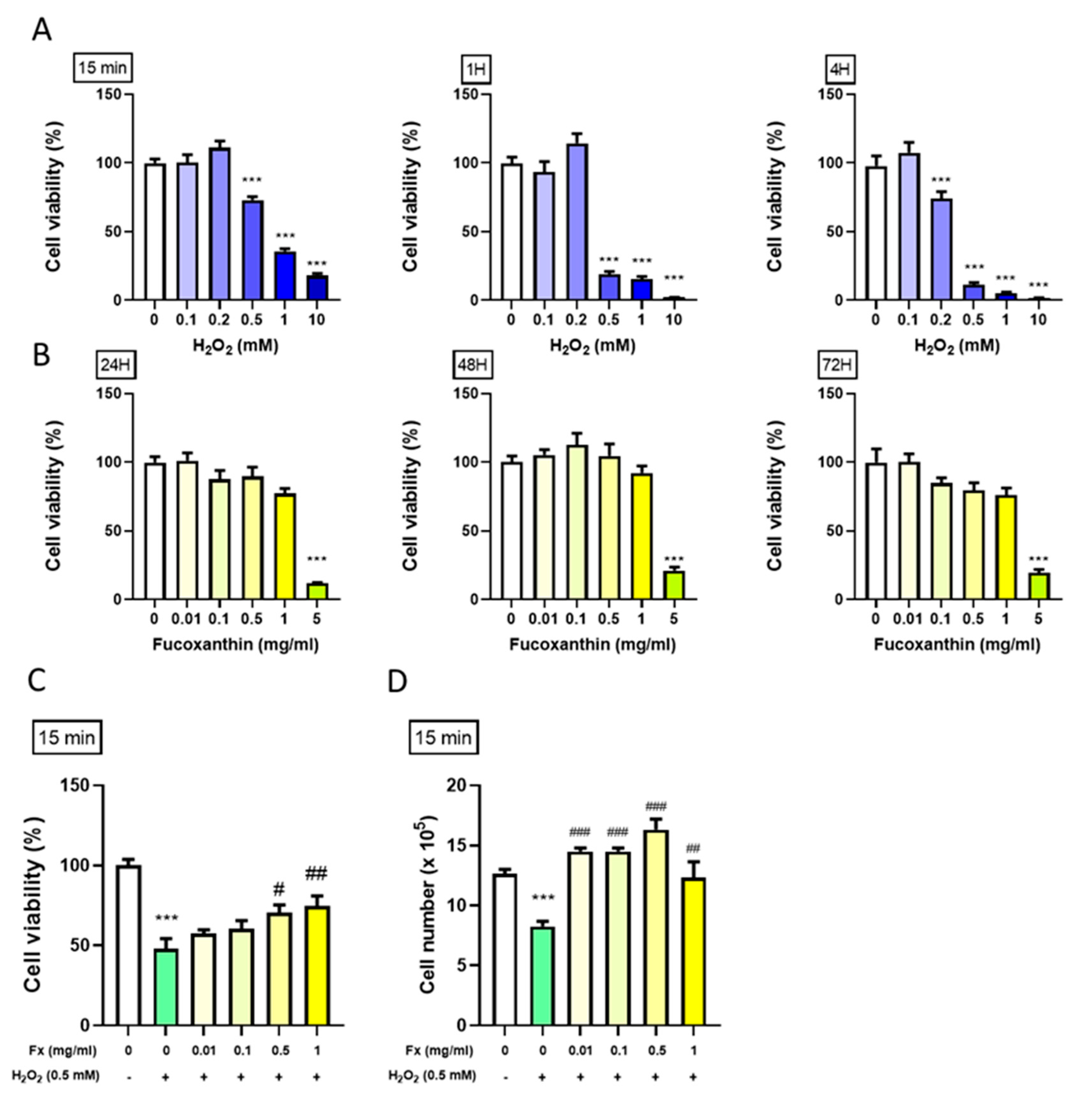
2.1. Protective Effect of Fx on VIC Cell Viability from H2O2-Induced Oxidative Stress
2.1.1. Effect of H2O2 on the VIC Cell Viability
2.1.2. Effect of Fx on VIC Cell Viability
2.1.3. Fx-Abrogated H2O2-Induced VIC Viability Change
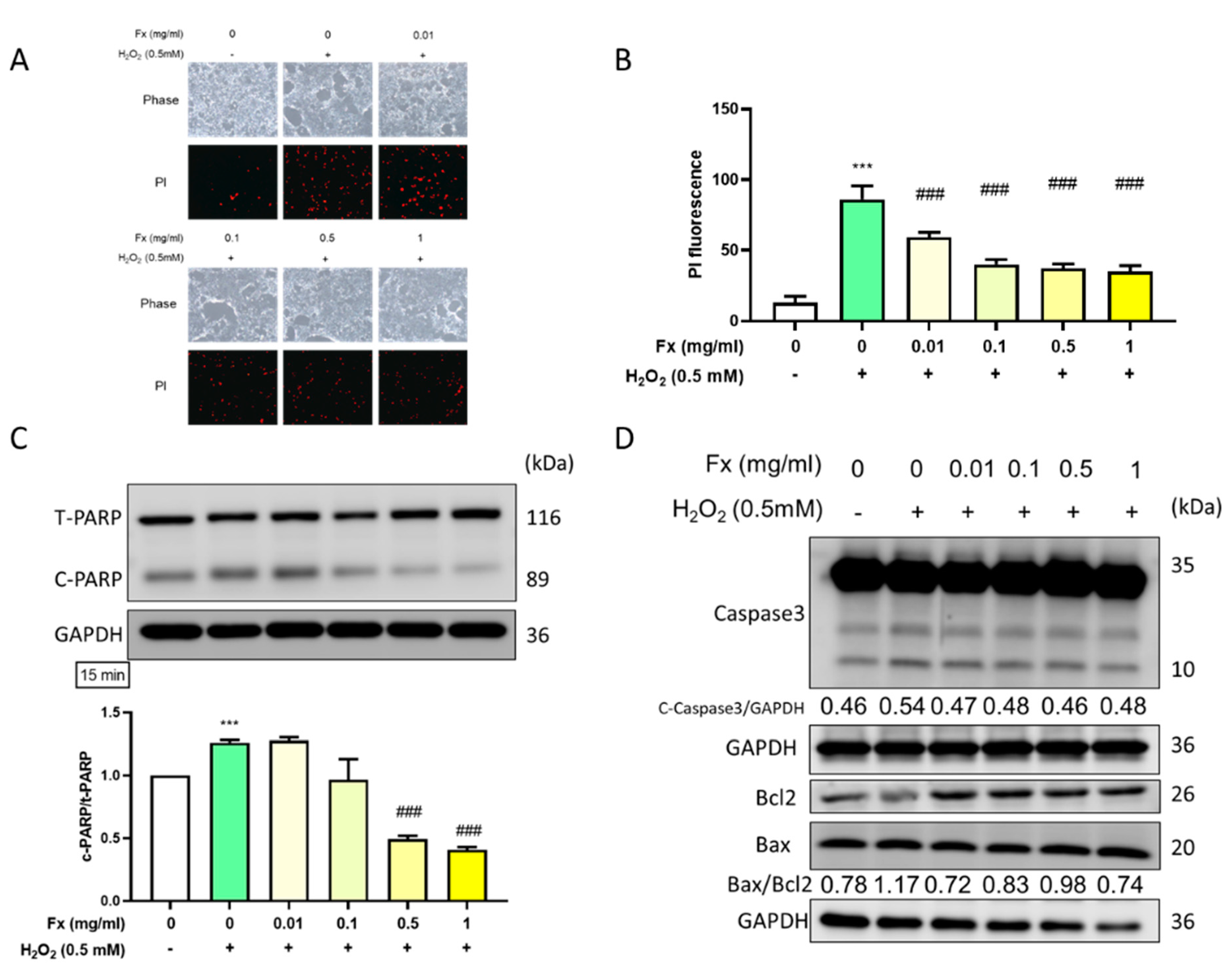
2.2. Fx-Ameliorated H2O2-Induced DNA Damage and Apoptosis-Related Protein Expression
2.2.1. Fx Decreased H2O2-Induced VIC Cell Morphology Changes and DNA Damage
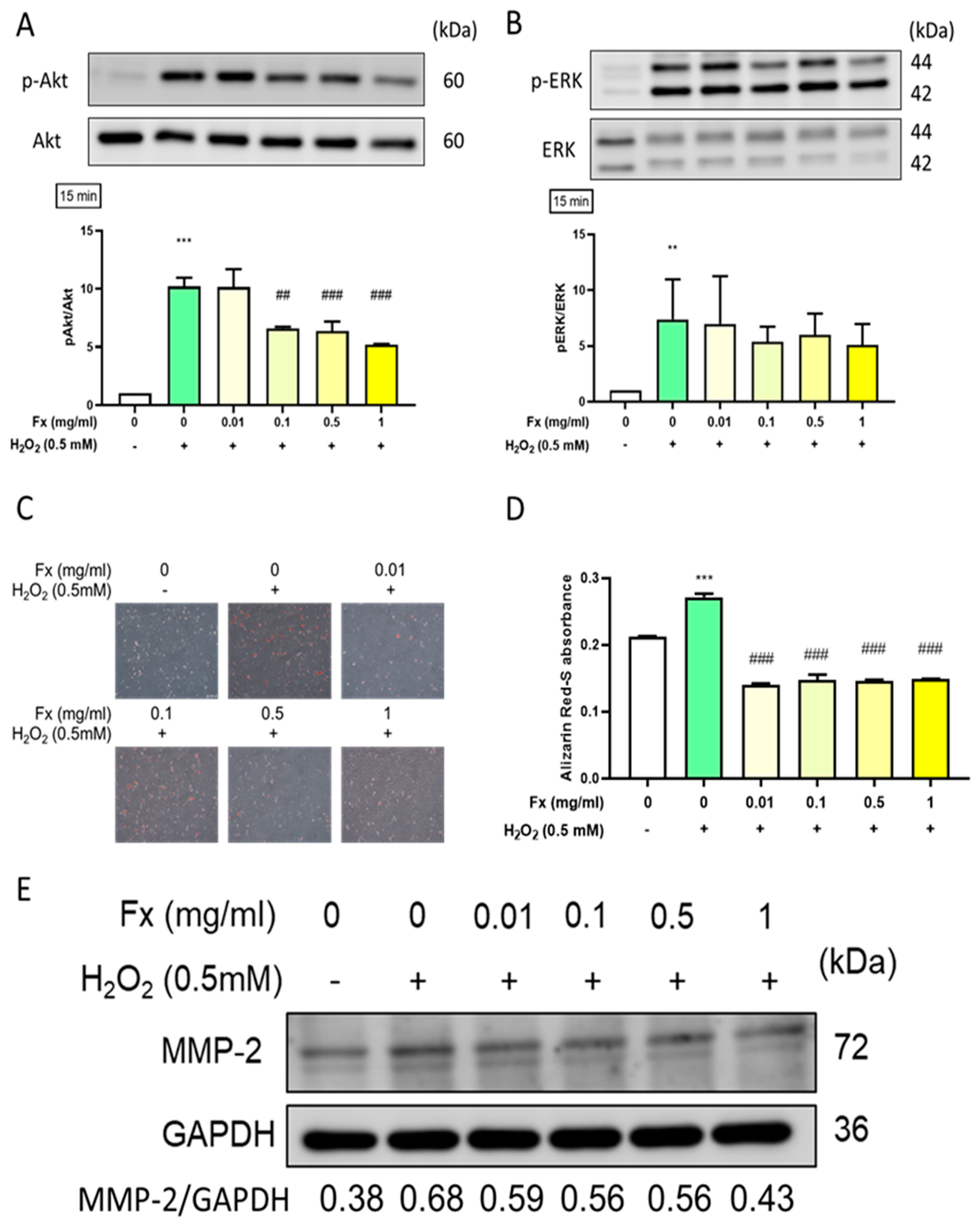
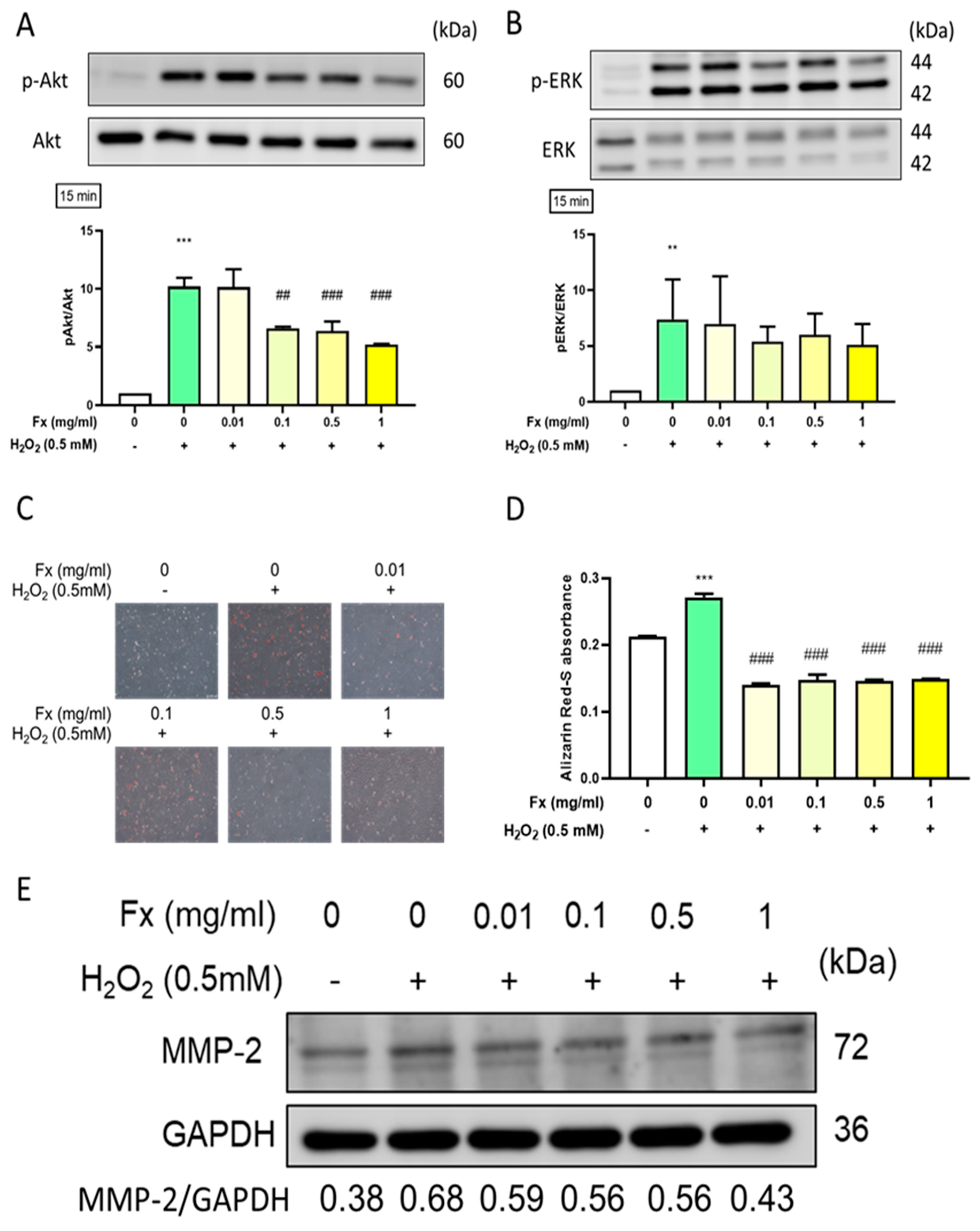
2.2.2. Fx-Ameliorated H2O2-Induced Apoptosis-Related Protein Expression
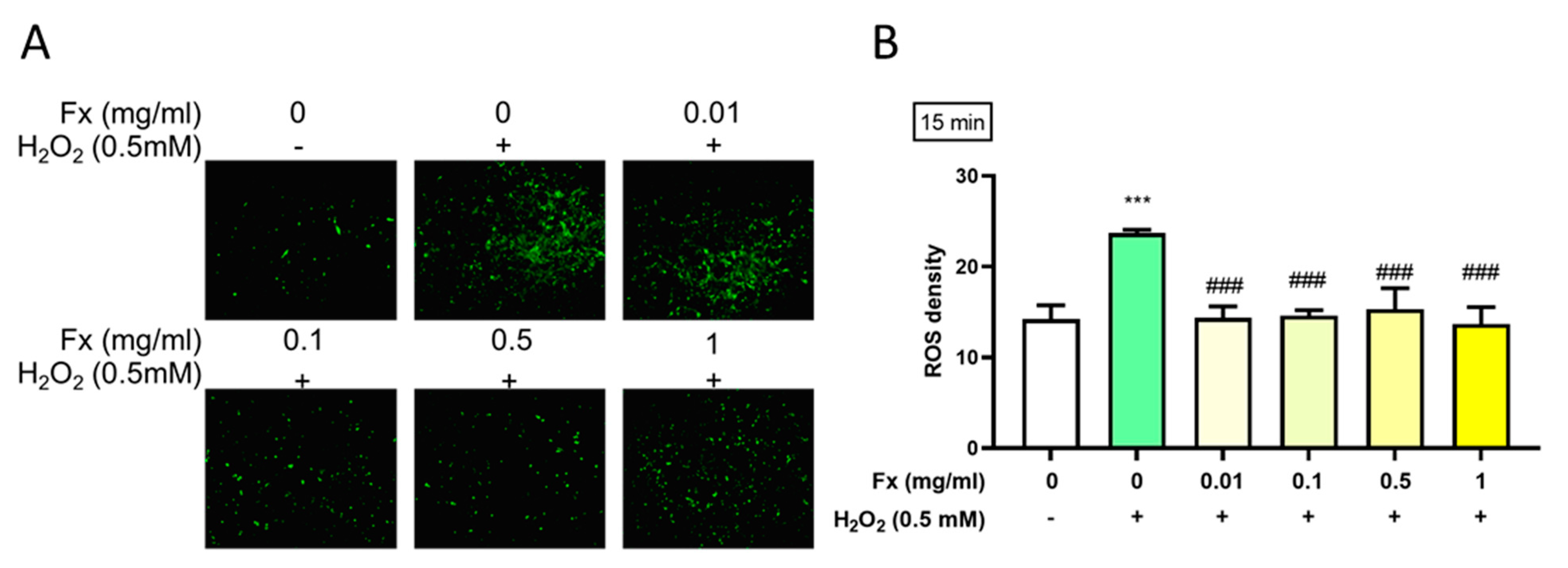
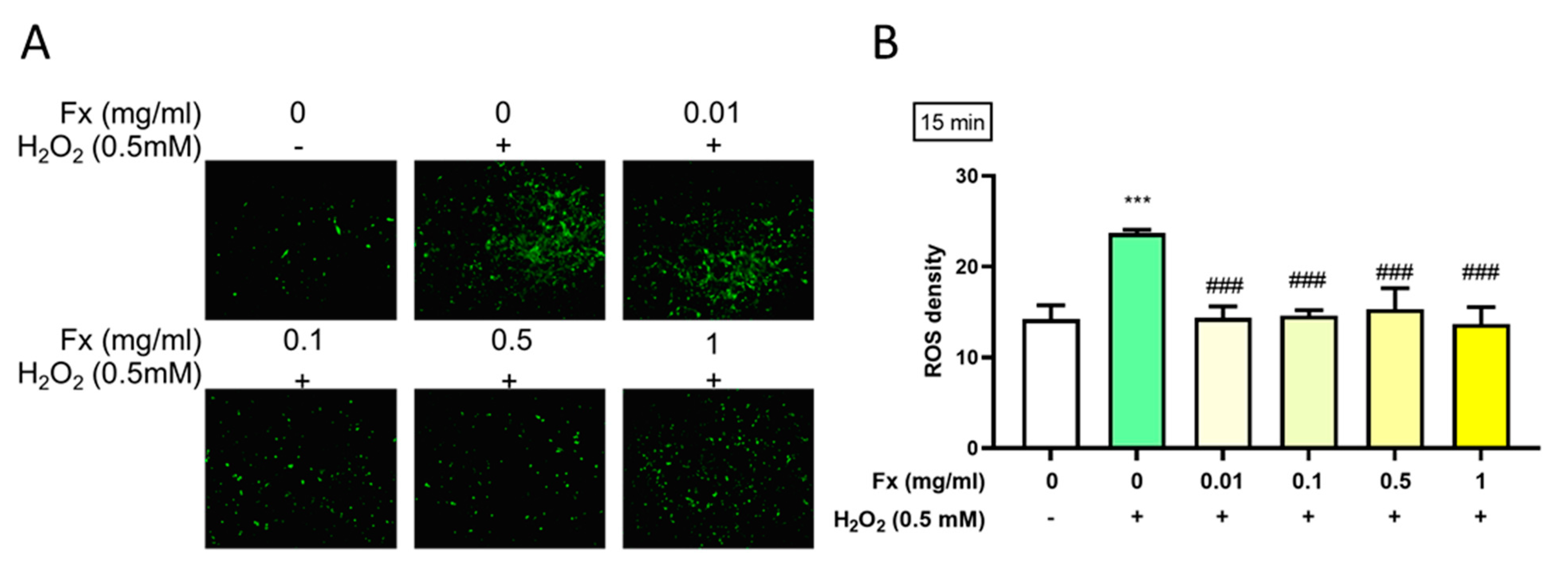
2.3. Effect of Fx on H2O2-Induced Reactive Oxygen Species
2.4. Effect of Fx on Oxidative Stress-Induced Calcification and the Expression of Its Related Markers in VIC
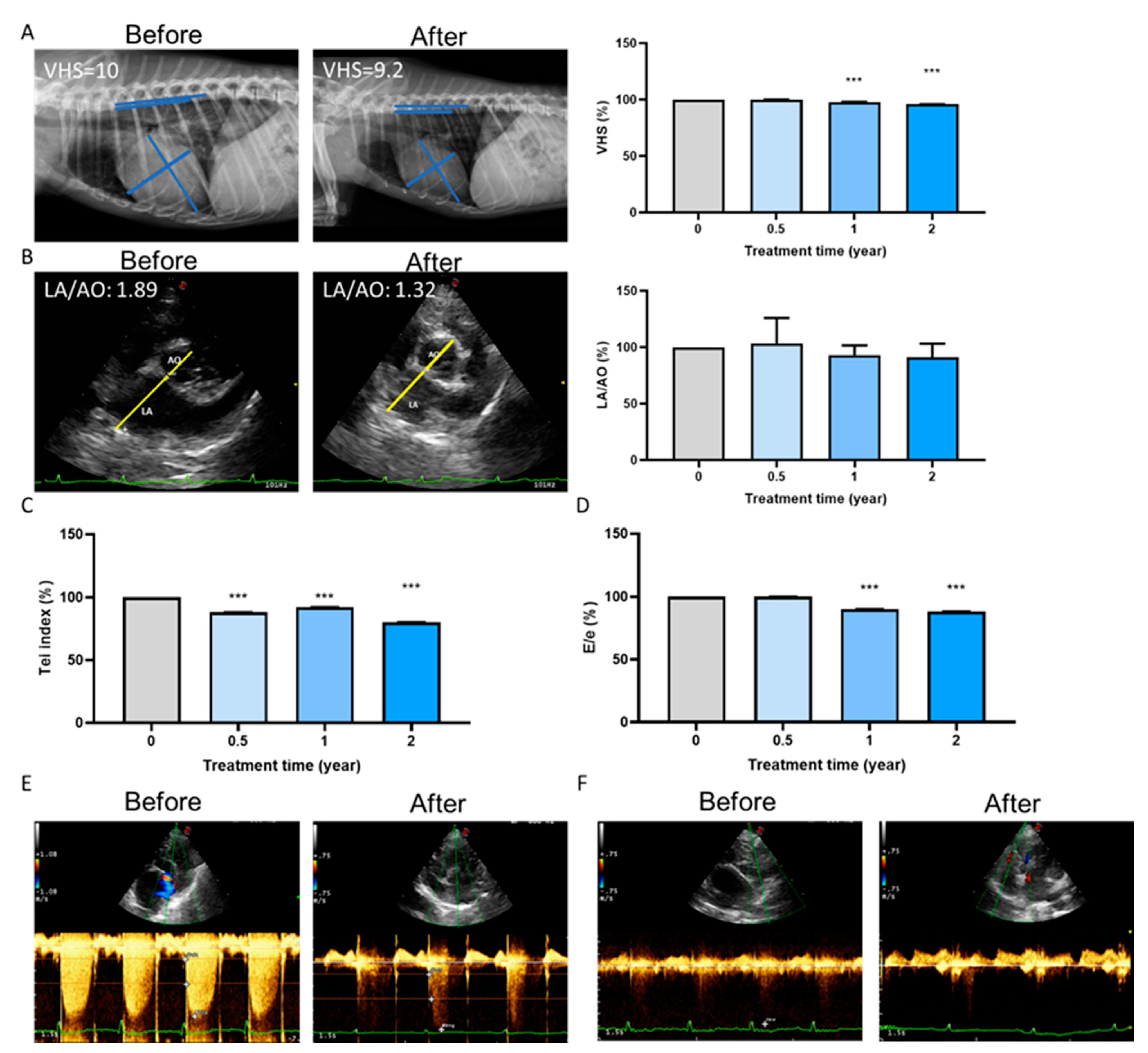
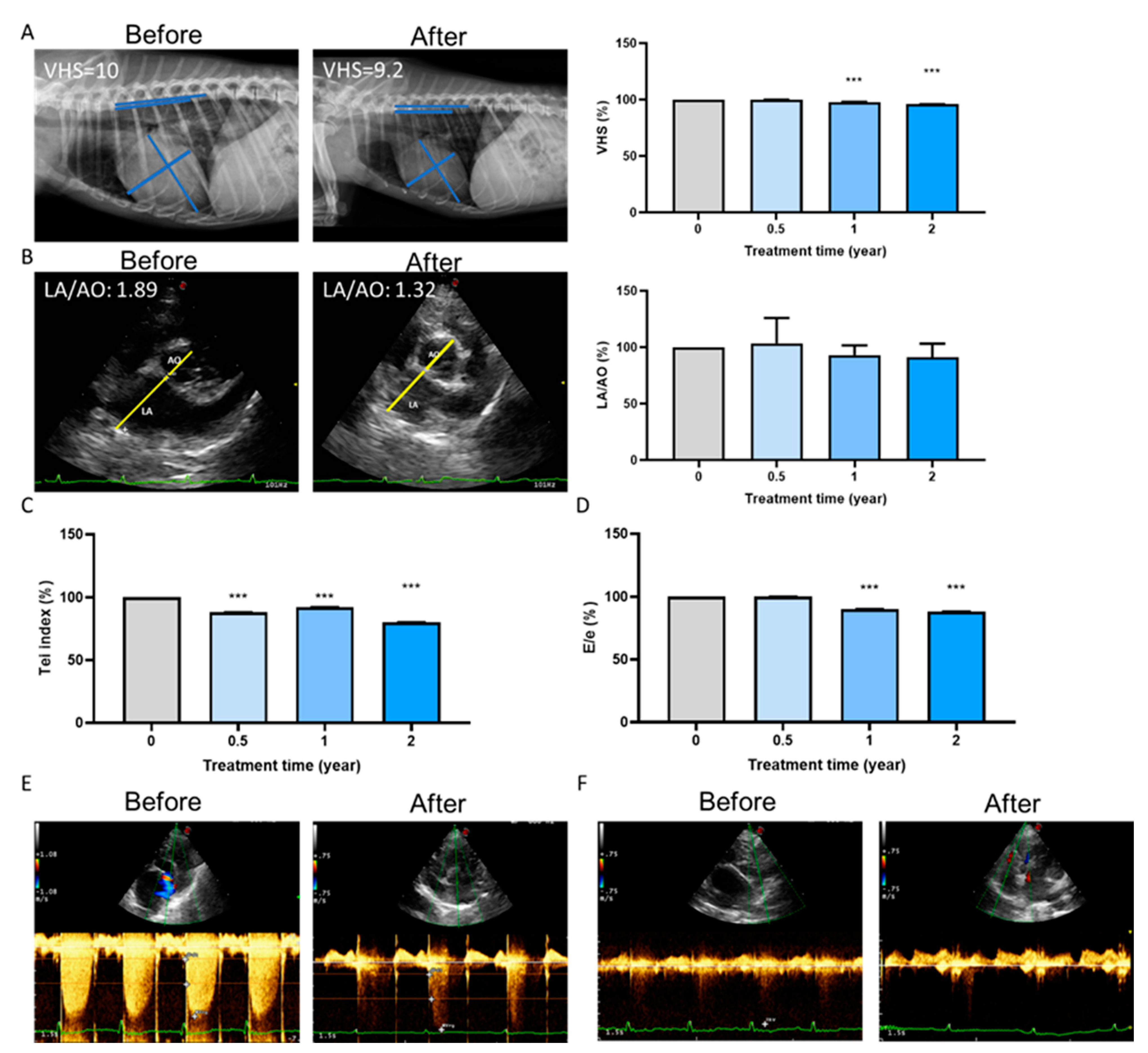
2.5. Long-Term Cardioprotective Effect of Fx Treatment in Dogs
3. Discussion
4. Materials and Methods
4.1. Reagents Preparation
4.2. Cell Extraction and Treatment
4.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
4.4. Cell Counting
4.5. PI Staining
4.6. ROS Density Measurement
4.7. Alizarin Red-S Staining Assay
4.8. Protein Extraction and Western Blot
4.9. In Vivo Animal Experiments
4.10. Statistical Analysis
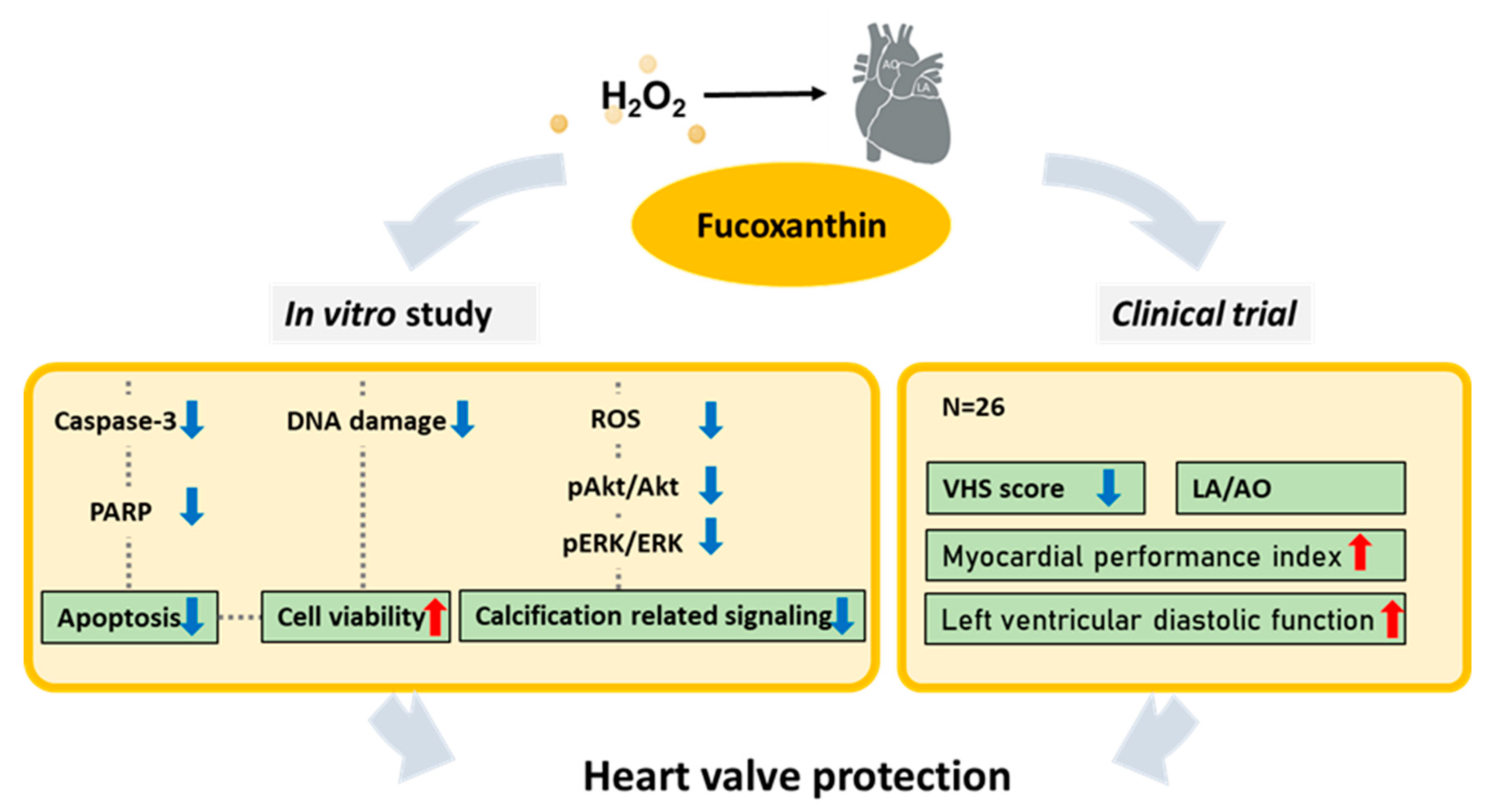
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Approval for Experiments on Animals
References
- Wasmus, C.; Dudek, J. Metabolic Alterations Caused by Defective Cardiolipin Remodeling in Inherited Cardiomyopathies. Life 2020, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.T.; Srigunapalan, S.; Simmons, C.A.; Anseth, K.S. Hemodynamic and Cellular Response Feedback in Calcific Aortic Valve Disease. Circ. Res. 2013, 113, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Towler, D.A. Oxidation, inflammation, and Aortic Valve Calcification Peroxide Paves an Osteogenic Path. J. Am. Coll. Cardiol. 2008, 52, 851–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaitanya, G.V.; Steven, A.J.; Babu, P.P. PARP-1 Cleavage Fragments: Signatures of Cell-death Proteases in Neurodegeneration. CCS 2010, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Vito, A.; Donato, A.; Presta, I.; Mancuso, T.; Brunetti, F.S.; Mastroroberto, P.; Amorosi, A.; Malara, N.; Donato, G. Extracellular Matrix in Calcific Aortic Valve Disease: Architecture, Dynamic and Perspectives. Int. J. Mol. Sci. 2021, 22, 913. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, D.; Yuan, P.; Li, J.; Yun, Y.; Cui, Y.; Zhang, T.; Ma, J.; Sun, L.; Ma, H.; et al. Endothelial-to-Mesenchymal Transition in Calcific Aortic Valve Disease. Acta Cardiol. Sin. 2020, 36, 183–194. [Google Scholar] [PubMed]
- Nagy, E.; Eriksson, P.; Yousry, M.; Caidahl, K.; Ingelsson, E.; Hansson, G.K.; Franco-Cereceda, A.; Bäck, M. Valvular Osteoclasts in Calcification and Aortic Valve Stenosis Severity. Int. J. Cardiol. 2013, 168, 2264–2271. [Google Scholar] [CrossRef] [Green Version]
- Nsaibia, M.J.; Boulanger, M.C.; Bouchareb, R.; Mkannez, G.; Le Quang, K.; Hadji, F.; Argaud, D.; Dahou, A.; Bossé, Y.; Koschinsky, M.L.; et al. OxLDL-derived Lysophosphatidic Acid Promotes the Progression of Aortic Valve Stenosis Through A LPAR1-Rhoa-NF-ΚB Pathway. Cardiovasc. Res. 2017, 113, 1351–1363. [Google Scholar] [CrossRef] [Green Version]
- Keene, B.W.; Atkins, C.E.; Bonagura, J.D.; Fox, P.R.; Häggström, J.; Fuentes, V.L.; Oyama, M.A.; Rush, J.E.; Stepien, R.; Uechi, M. ACVIM Consensus Guidelines for the Diagnosis and Treatment of Myxomatous Mitral Valve Disease in Dogs. J. Vet. Intern Med. 2019, 33, 1127–1140. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Chen, H.-Y.; Hsia, S.-M. Four case reports: Effects of Fucoidan and Fucoxanthin on the Treatment of Degenerative Heart Valve Disease in Dogs. J. Anim. Sci. Vet. Med. 2020, 5, 114–122. [Google Scholar]
- Liu, Y.; Liu, M.; Zhang, X.; Chen, Q.; Chen, H.; Sun, L.; Liu, G. Protective Effect of Fucoxanthin Isolated from Laminaria japonica against Visible Light-Induced Retinal Damage Both in Vitro and in Vivo. J. Agric. Food Chem. 2016, 64, 416–424. [Google Scholar] [CrossRef]
- Iwasaki, S.; Widjaja-Adhi, M.A.K.; Koide, A.; Kaga, T.; Nakano, S.; Beppu, F.; Hosokawa, M.; Miyashita, K. In Vivo Antioxidant Activity of Fucoxanthin on Obese/Diabetes KK-Ay Mice. Food Nutr. Sci. 2012, 3, 1491–1499. [Google Scholar]
- Prasedya, E.S.; Martyasari, N.W.R.; Abidin, A.S.; Pebriani, S.A.; Ilhami, B.T.K.; Frediansyah, A.; Sunarwidhi, A.L.; Widyastuti, S.; Sunarpi, H. Macroalgae Sargassum cristaefolium Extract Inhibits Proinflammatory Cytokine Expression in BALB/C Mice. Scientifica (Cairo) 2020, 2020, 9769454. [Google Scholar] [PubMed]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal Carotenoids And Phytosterols Regulate Biochemical Mechanisms Involved in Human Health and Disease Prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Chau, Y.-T.; Chen, H.-Y.; Lin, P.-H.; Hsia, S.-M. Preventive Effects of Fucoidan and Fucoxanthin on Hyperuricemic Rats Induced by Potassium Oxonate. Mar. Drugs 2019, 17, 343. [Google Scholar] [CrossRef] [Green Version]
- Hentati, F.; Tounsi, L.; Djomdi, D.; Pierre, G.; Delattre, C.; Ursu, A.V.; Fendri, I.; Abdelkafi, S.; Michaud, P. Bioactive Polysaccharides from Seaweeds. Molecules 2020, 25, 3152. [Google Scholar] [CrossRef] [PubMed]
- Branchetti, E.; Sainger, R.; Poggio, P.; Grau, J.B.; Patterson-Fortin, J.; Bavaria, J.E.; Chorny, M.; Lai, E.; Gorman, R.C.; Levy, R.J.; et al. Antioxidant Enzymes Reduce DNA Damage and Early Activation of Valvular Interstitial Cells in Aortic Valve Sclerosis. Arterioscler. Thromb. Vasc. Biol. 2013, 33, e66–e74. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Torzewski, M. Fibroblasts and Their Pathological Functions in the Fibrosis of Aortic Valve Sclerosis and Atherosclerosis. Biomolecules 2019, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, Y.; Liu, M.; Huang, Y.; Shi, J.; Dong, N.; Xu, K. Curcumin Inhibits Calcification of Human Aortic Valve Interstitial Cells by Interfering NF-κB, AKT, and ERK Pathways. Phytother. Res. 2020, 34, 2074–2081. [Google Scholar] [CrossRef]
- Nakayama, H.; Nakayama, T.; Hamlin, R.L. Correlation of Cardiac Enlargement as Assessed by Vertebral Heart Size and Echocardiographic and Electrocardiographic Findings in Dogs with Evolving Cardiomegaly due to Rapid Ventricular Pacing. J. Vet. Intern. Med. 2001, 15, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Galizzi, A.; Bagardi, M.; Stranieri, A.; Zanaboni, A.M.; Malchiodi, D.; Borromeo, V.; Brambilla, P.G.; Locatelli, C. Factors Affecting the Urinary Aldosterone-to-Creatinine Ratio in Healthy Dogs and Dogs with Naturally Occurring Myxomatous Mitral Valve Disease. BMC Vet. Res. 2021, 17, 15. [Google Scholar] [CrossRef]
- Heistad, D.D. Oxidative stress and Vascular Disease: 2005 Duff lecture. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Griendling, K.K.; FitzGerald, G.A. Oxidative Stress and Cardiovascular Injury: Part II: Animal and Human studies. Circulation 2003, 108, 2034–2040. [Google Scholar] [CrossRef]
- Wei, E.P.; Kontos, H.A.; Christman, C.W.; DeWitt, D.S.; Povlishock, J.T. Superoxide Generation and Reversal of Acetylcholine-induced Cerebral Arteriolar Dilation after Acute Hypertension. Circ. Res. 1985, 57, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Weiss, R.M.; Ohashi, M.; Miller, J.D.; Young, S.G.; Heistad, D.D. Calcific Aortic Valve Stenosis in Old Hypercholesterolemic Mice. Circulation 2006, 114, 2065–2069. [Google Scholar] [CrossRef]
- Misteli, T.; Soutoglou, E. The Emerging Role of Nuclear Architecture in DNA Repair and Genome Maintenance. Nat. Rev. Mol. Cell Biol. 2009, 10, 243–254. [Google Scholar] [CrossRef]
- Byon, C.H.; Javed, A.; Dai, Q.; Kappes, J.C.; Clemens, T.L.; Darley-Usmar, V.M.; McDonald, J.M.; Chen, Y. Oxidative Stress Induces Vascular Calcification through Modulationo of the Osteogenic Transcription Factor Runx2 by AKT Signaling. J. Biol. Chem. 2008, 283, 15319–15327. [Google Scholar] [CrossRef] [Green Version]
- Scott, M.; Vesely, I. Aortic Valve Cusp Microstructure: The Role of Elastin. Ann. Thorac. Surg. 1995, 60, S391–S394. [Google Scholar] [CrossRef]
- O’Brien, K.D.; Reichenbach, D.D.; Marcovina, S.M.; Kuusisto, J.; Alpers, C.E.; Otto, C.M. Apolipoproteins B, (a), and E accumulate in the Morphologically Early Lesion of ‘Degenerative’ Valvular Aortic Stenosis. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 523–532. [Google Scholar] [CrossRef]
- Farrar, E.J.; Huntley, G.D.; Butcher, J. Endothelial-derived Oxidative Stress drives Myofibroblastic Activation and Calcification of the Aortic Valve. PLoS ONE 2015, 10, e0123257. [Google Scholar] [CrossRef]
- Pawade, T.A.; Newby, D.E.; Dweck, M.R. Calcification in Aortic Stenosis: The Skeleton Key. J. Am. Coll. Cardiol. 2015, 66, 561–577. [Google Scholar] [CrossRef] [Green Version]
- Helske, S.; Syväranta, S.; Lindstedt, K.A.; Lappalainen, J.; Oörni, K.; Mäyränpää, M.I.; Lommi, J.; Turto, H.; Werkkala, K.; Kupari, M.; et al. Increased Expression of Elastolytic Cathepsins S, K, and V and their Inhibitor Cystatin C in Stenotic Aortic Valves. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1791–1798. [Google Scholar] [CrossRef] [Green Version]
- Kee, H.J.; Cho, S.N.; Kim, G.R.; Choi, S.Y.; Ryu, Y.; Kim, I.K.; Hong, Y.J.; Park, H.W.; Ahn, Y.; Cho, J.G.; et al. Gallic Acid Inhibits Vascular Calcification Through the Blockade of BMP2-Smad1/5/8 Signaling Pathway. Vascul. Pharmacol. 2014, 63, 71–78. [Google Scholar] [CrossRef]
- Roman-Garcia, P.; Barrio-Vazquez, S.; Fernandez-Martin, J.L.; Ruiz-Torres, M.P.; Cannata-Andia, J.B. Natural Antioxidants and Vascular Calcification: A Possible Benefit. J. Nephrol. 2011, 24, 669–672. [Google Scholar] [CrossRef]
- Lu, T.S.; Lim, K.; Molostvov, G.; Yang, Y.C.; Yiao, S.Y.; Zehnder, D.; Hsiao, L.L. Induction of Intracellular Heat-shock protein 72 prevents the Development of Vascular Smooth Muscle Cell Calcification. Cardiovasc. Res. 2012, 96, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.-T.; Yeh, H.-Y.; Tsai, Y.-T.; Chuang, P.-H.; Yuan, T.-H.; Huang, J.-W.; Chen, H.-W. Natural and Non-Natural Antioxidative Compounds: Potential Candidates for Treatment of Vascular Calcification. Cell Death Discov. 2019, 5, 145. [Google Scholar] [CrossRef]
- Menichini, D.; Alrais, M.; Liu, C.; Xia, Y.; Blackwell, S.C.; Facchinetti, F.; Sibai, B.M.; Longo, M. Maternal Supplementation of Inositols, Fucoxanthin, and Hydroxytyrosol in Pregnant Murine Models of Hypertension. Am. J. Hypertens. 2020, 33, 652–659. [Google Scholar] [CrossRef]
- Chiang, Y.F.; Chen, H.Y.; Chang, Y.J.; Shih, Y.H.; Shieh, T.M.; Wang, K.L.; Hsia, S.M. Protective Effects of Fucoxanthin on High Glucose- and 4-Hydroxynonenal (4-HNE)-Induced Injury in Human Retinal Pigment Epithelial Cells. Antioxidants (Basel) 2020, 9, 1176. [Google Scholar] [CrossRef]
- Ou, H.C.; Chou, W.C.; Chu, P.M.; Hsieh, P.L.; Hung, C.H.; Tsai, K.L. Fucoxanthin Protects Against oxLDL-Induced Endothelial Damage via Activating the AMPK-Akt-CREB-PGC1α Pathway. Mol. Nutr. Food Res. 2019, 63, e1801353. [Google Scholar] [CrossRef]
- Casamián-Sorrosal, D.; Barrera-Chacón, R.; Fonfara, S.; Belinchón-Lorenzo, S.; Gómez-Gordo, L.; Galapero-Arroyo, J.; Fernández-Cotrina, J.; Cristobal-Verdejo, J.I.; Duque, F.J. Association of Myocardial Parasitic Load with Cardiac Biomarkers and Other Selected Variables in 10 Dogs with Advanced Canine Leishmaniasis. Vet. Rec. 2021, e198. [Google Scholar] [CrossRef]
- Lee, S.W.; Park, M.C.; Park, Y.B.; Lee, S.K. E/E’ Ratio is more Sensitive Than E/A Ratio for Detection of Left Ventricular Diastolic Dysfunction in Systemic Lupus Erythematosus. LUPUS 2008, 17, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhu, D.; Markby, G.; Corcoran, B.M.; Farquharson, C.; Macrae, V.E. Isolation and Characterization of Primary Rat Valve Interstitial Cells: A New Model to Study Aortic Valve Calcification. J. Vis. Exp. 2017, 56126. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, Y.-F.; Tsai, C.-H.; Chen, H.-Y.; Wang, K.-L.; Chang, H.-Y.; Huang, Y.-J.; Hong, Y.-H.; Ali, M.; Shieh, T.-M.; Huang, T.-C.; et al. Protective Effects of Fucoxanthin on Hydrogen Peroxide-Induced Calcification of Heart Valve Interstitial Cells. Mar. Drugs 2021, 19, 307. https://doi.org/10.3390/md19060307
Chiang Y-F, Tsai C-H, Chen H-Y, Wang K-L, Chang H-Y, Huang Y-J, Hong Y-H, Ali M, Shieh T-M, Huang T-C, et al. Protective Effects of Fucoxanthin on Hydrogen Peroxide-Induced Calcification of Heart Valve Interstitial Cells. Marine Drugs. 2021; 19(6):307. https://doi.org/10.3390/md19060307
Chicago/Turabian StyleChiang, Yi-Fen, Chih-Hung Tsai, Hsin-Yuan Chen, Kai-Lee Wang, Hsin-Yi Chang, Yun-Ju Huang, Yong-Han Hong, Mohamed Ali, Tzong-Ming Shieh, Tsui-Chin Huang, and et al. 2021. "Protective Effects of Fucoxanthin on Hydrogen Peroxide-Induced Calcification of Heart Valve Interstitial Cells" Marine Drugs 19, no. 6: 307. https://doi.org/10.3390/md19060307
APA StyleChiang, Y.-F., Tsai, C.-H., Chen, H.-Y., Wang, K.-L., Chang, H.-Y., Huang, Y.-J., Hong, Y.-H., Ali, M., Shieh, T.-M., Huang, T.-C., Lin, C.-I., & Hsia, S.-M. (2021). Protective Effects of Fucoxanthin on Hydrogen Peroxide-Induced Calcification of Heart Valve Interstitial Cells. Marine Drugs, 19(6), 307. https://doi.org/10.3390/md19060307