
Enzymatic Degradation of Gracilariopsis lemaneiformis Polysaccharide and the Antioxidant Activity of Its Degradation Products


Abstract
1. Introduction
2. Results and Discussion
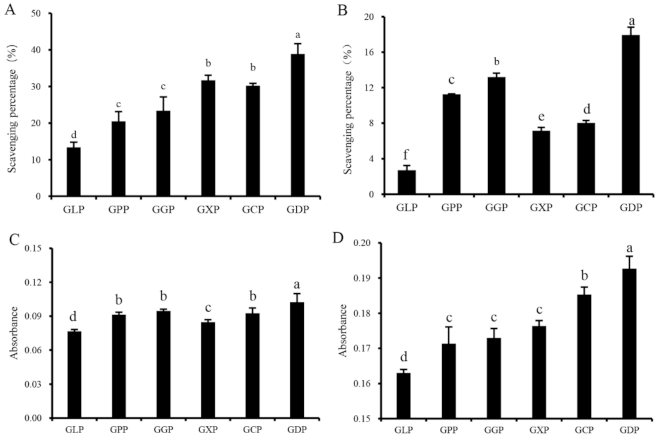
2.1. Chemical Analysis of Polysaccharides
2.2. Fourier Transform Infrared Spectra (FT-IR) Analysis
2.3. Screening of Degradation Enzymes
2.4. Single-Factor Experiment
2.5. Orthogonal Optimization Experiment and Verification Test
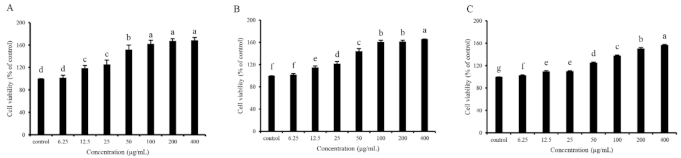
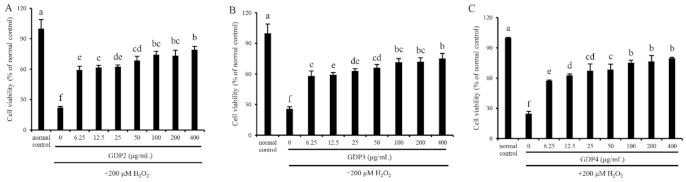
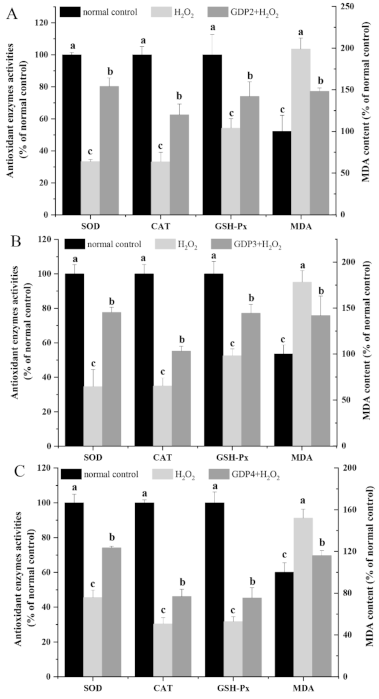
2.6. GDP Alleviated H2O2-Induced Oxidative Injury in HFL1 Cells
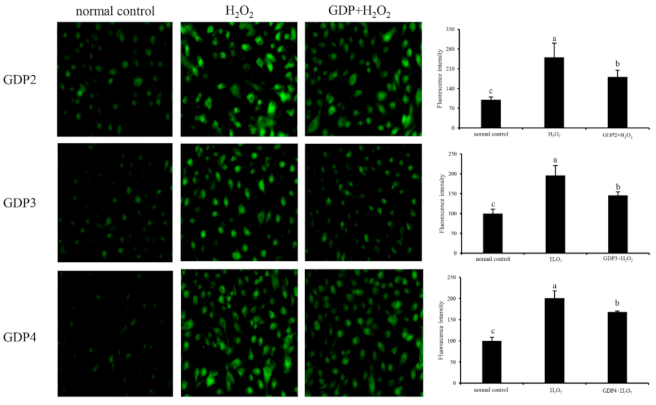
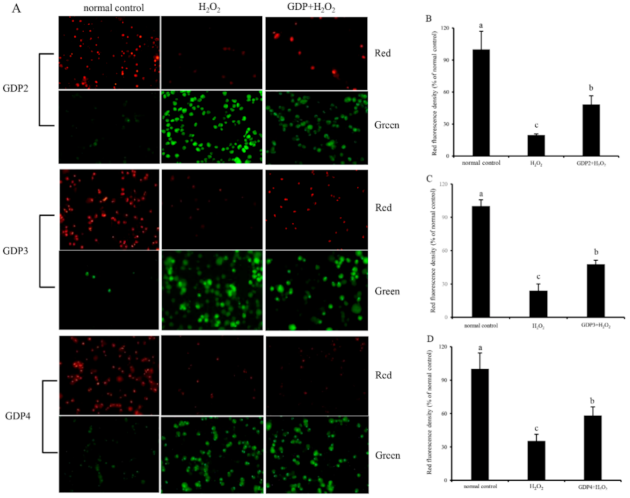
2.7. GDP Reduced ROS Generation in the HFL1 Cells
2.8. GDP Alleviated Lipid Peroxidation Level and Increased the Cellular Antioxidant Enzymes Activities of HFL1 Cells
2.9. GDP Inhibited Mitochondrial Membrane Potential (MMP) Decline in HFL1 Cells
3. Materials and Methods
3.1. Materials and Reagents
3.2. GLP Extraction
3.3. Enzymatic Hydrolysis of GLP
3.4. Physicochemical Properties Analysis of Polysaccharides
3.5. Single-Factor and Orthogonal Optimization Experiments
3.6. In Vitro Chemical Assays of Antioxidant Activity
3.6.1. Hydroxyl Radical Scavenging Activity
3.6.2. DPPH Radical Scavenging Activity
3.6.3. Reduction Capacity
3.6.4. Total Antioxidant Capacity
3.7. In Vitro Biological Assays of Antioxidant Activity
3.7.1. Cell Culture and Cell Viability Assay
3.7.2. Measurement of Intracellular ROS Generation
3.7.3. Cellular Antioxidant Activity Measurement
3.7.4. Determination of MMP
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.T.; Jan, C.R.; Liang, W.Z. Protective effects of a phenolic glycoside compound curculigoside on H2O2-induced oxidative stress and cytotoxicity in normal human breast epithelial cells. J. Funct. Foods 2018, 41, 171–182. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.W.; Wei, B.; Wang, S.J.; Ke, S.Z.; Chen, J.W.; Zhang, H.W.; Wang, H. The antioxidant activity of polysaccharides derived from marine organisms: An overview. Mar. Drugs. 2019, 17, 674. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.X.; Ge, Z.F.; Fan, Y.J.; Luo, A.S.; Chun, Z.; He, X.J. In vitro and in vivo antioxidant activity of a water-soluble polysaccharide from Dendrobium denneanum. Molecules 2011, 16, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, H.H.; Ko, J.Y.; Jang, J.H.; Kim, G.H.; Lee, J.S.; Nah, J.W.; Jeon, Y.J. Rapid preparation of functional polysaccharides from Pyropia yezoensis by microwave-assistant rapid enzyme digest system. Carbohydr. Polym. 2016, 153, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.N.; Wang, Z.J.; Xie, D.S.; Sun, X.; Yang, W.G.; Zhao, X.D.; Xu, N.J. Characterization and potential antitumor activity of polysaccharide from Gracilariopsis lemaneiformis. Mar. Drugs 2017, 15, 100. [Google Scholar] [CrossRef]
- Han, R.; Pang, D.; Wen, L.; You, L.; Huang, R.; Kulikouskaya, V. In vitro digestibility and prebiotic activities of a sulfated polysaccharide from Gracilaria Lemaneiformis. J. Funct. Foods 2020, 64, 103652. [Google Scholar] [CrossRef]
- Li, X.; Huang, S.M.; Chen, X.; Xu, Q.J.; Ma, Y.X.; You, L.J.; Kulikouskaya, V.; Xiao, J.B.; Piao, J.H. Structural characteristic of a sulfated polysaccharide from Gracilaria lemaneiformis and its lipid metabolism regulation effect. Food Funct. 2020, 11, 10876–10885. [Google Scholar] [CrossRef]
- Ju, T.; Xi, J. Continuous extraction optimization, molecular structures and antioxidant activities of polysaccharide from Gracilariopsis lemaneiformis using liquid-phase pulsed discharge. Sep. Purif. Technol. 2020, 236, 116241. [Google Scholar] [CrossRef]
- Wu, S.; Lu, M.; Wang, S. Amylase-assisted extraction and antioxidant activity of polysaccharides from Gracilaria lemaneiformis. Biotech 2017, 7, 38. [Google Scholar] [CrossRef]
- Guo, D.; Yu, K.; Sun, X.Y.; Ouyang, J.M. Structural characterization and repair mechanism of Gracilaria lemaneiformis sulfated polysaccharides of different molecular weights on damaged renal epithelial cells. Oxid. Med. Cell. Longev. 2018, 2018, 7410389. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol 2002, 60, 258–274. [Google Scholar]
- Jin, M.; Zhao, K.; Huang, Q.; Xu, C.; Shang, P. Isolation, structure and bioactivities of the polysaccharides from Angelica sinensis (Oliv.) Diels: A review. Carbohydr. Polym. 2012, 89, 713–722. [Google Scholar] [CrossRef]
- Alban, S.; Franz, G. Characterization of the anticoagulant actions of a semisynthetic curdlan sulfate. Thromb. Res. 2000, 99, 377–388. [Google Scholar] [CrossRef]
- Sun, L.; Chu, J.; Sun, Z.; Chen, L. Physicochemical properties, immunomodulation and antitumor activities of polysaccharide from Pavlova viridis. Life Sci. 2016, 144, 156–161. [Google Scholar] [CrossRef]
- Wu, G.J.; Shiu, S.M.; Hsieh, M.C.; Tsai, G.J. Anti-inflammatory activity of a sulfated polysaccharide from the brown alga Sargassum cristaefolium. Food Hydrocolloid. 2016, 53, 16–23. [Google Scholar] [CrossRef]
- Bhadja, P.; Tan, C.Y.; Ouyang, J.M.; Yu, K. Repair effect of seaweed polysaccharides with different contents of sulfate group and molecular weights on damaged HK-2 cells. Polymers 2016, 8, 188. [Google Scholar] [CrossRef]
- Zhou, C.S.; Yu, X.J.; Zhang, Y.Z.; He, R.H.; Ma, H. Ultrasonic degradation, purification and analysis of structure and antioxidant activity of polysaccharide from Porphyra yezoensis Udea. Carbohydr. Polym. 2012, 87, 2046–2051. [Google Scholar] [CrossRef]
- Barreteau, H.; Delattre, C.; Michaud, P. Production of oligosaccharides as promising new food additive generation. Food Technol. Biotech. 2006, 44, 323–333. [Google Scholar]
- Xiong, F.; Li, X.; Zheng, H.; Hu, N.; Li, H. Characterization and antioxidant activities of polysaccharides from Passiflora edulis Sims peel under different degradation methods. Carbohydr. Polym. 2019, 218, 46–52. [Google Scholar] [CrossRef]
- Liao, X.B.; Yang, L.W.; Chen, M.Z.; Yu, J.; Zhang, S.M.; Ju, Y.Y. The hypoglycemic effect of a polysaccharide (GLP) from Gracilaria lemaneiformis and its degradation products in diabetic mice. Food Funct. 2015, 6, 2542–2549. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Gao, Y.; Daniel, G. Effects of epigallocatechin gallate, epigallocatechin and epicatechin gallate on the chemical and cell-based antioxidant activity, sensory properties, and cytotoxicity of a catechin-free model beverage. Food Chem. 2021, 339, 128060. [Google Scholar] [CrossRef]
- Ouyang, J.M.; Wang, M.; Lu, P.; Tan, J. Degradation of sulfated polysaccharide extracted from algal Laminaria japonica and its modulation on calcium oxalate crystallization. Mater. Sci. Eng. C 2010, 30, 1022–1029. [Google Scholar] [CrossRef]
- Shi, F.L.; Yan, X.L.; Cheong, K.L.; Liu, Y. Extraction, purification, and characterization of polysaccharides from marine algae Gracilaria lemaneiformis with anti-tumor activity. Process Biochem. 2018, 73, 197–203. [Google Scholar] [CrossRef]
- Ji, C.F.; Ji, Y.B.; Meng, D.Y. Sulfated modification and anti-tumor activity of laminarin. Exp. Ther. Med. 2013, 6, 1259–1264. [Google Scholar] [CrossRef]
- Qiao, D.; Ke, C.; Hu, B.; Luo, J.; Ye, H.; Sun, Y.; Yan, X.; Zeng, X. Antioxidant activities of polysaccharides from Hyriopsis cumingii. Carbohydr. Polym. 2009, 78, 199–204. [Google Scholar] [CrossRef]
- Zheng, L.; Lin, L.; Su, G.; Zhao, Q.; Zhao, M. Pitfalls of using 1,1-diphenyl-2-picrylhydrazyl (DPPH) assay to assess the radical scavenging activity of peptides: Its susceptibility to interference and low reactivity towards peptides. Food Res. Int. 2015, 76, 359–365. [Google Scholar] [CrossRef]
- Lo, C.T.; Chang, C.A.; Chiu, K.H.; Tsay, P.K.; Jen, J.F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.S.; Wu, Y.; Sun, X.; Xu, N.J. Synthesized sulfated and acetylated derivatives of polysaccharide extracted from Gracilariopsis lemaneiformis and their potential antioxidant and immunological activity. Int. J. Biol. Macromol. 2019, 124, 568–572. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Y.C.; Jiang, Z.B.; Shi, G.G. Purification, partial characterization and bioactivity of sulfated polysaccharides from Grateloupia livida. Int. J. Biol. Macromol. 2017, 94, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Fan, L.; Ai, L.; Shan, L. Antioxidant activities of polysaccharides from the fruiting bodies of Zizyphus jujuba cv. jinsixiaozao. Carbohydr. Polym. 2011, 84, 390–394. [Google Scholar] [CrossRef]
- Sibakov, J.; Myllymki, O.; Suortti, T.; Kaukovirta-Norja, A.; Lehtinen, P.; Poutanen, K. Comparison of acid and enzymatic hydrolyses of oat bran β-glucan at low water content. Food Res. Int. 2013, 52, 99–108. [Google Scholar] [CrossRef]
- Chavan, S.; Gaikwad, A. Optimization of enzymatic hydrolysis of bamboo biomass for enhanced saccharification of cellulose through Taguchi orthogonal design. J. Environ. Chem. Eng. 2021, 9, 104807. [Google Scholar] [CrossRef]
- Kurd, F.; Samavati, V. Water soluble polysaccharides from Spirulina platensis: Extraction and in vitro anti-cancer activity. Int. J. Biol. Macromol. 2015, 74, 498–506. [Google Scholar] [CrossRef]
- Hu, J.; Jia, X.J.; Fang, X.B.; Li, P.; He, C.W.; Chen, M.W. Ultrasonic extraction, antioxidant and anticancer activities of novel polysaccharides from Chuanxiong rhizome. Int. J. Biol. Macromol. 2016, 85, 277–284. [Google Scholar] [CrossRef]
- De Oliveira Carvalho, J.; Orlanda, J.F.F. Heat stability and effect of pH on enzyme activity of polyphenol oxidase in buriti (Mauritia flexuosa linnaeus f.) fruit extract. Food Chem. 2017, 233, 159–163. [Google Scholar] [CrossRef]
- Pan, S.; Wu, S. Cellulase-assisted extraction and antioxidant activity of the polysaccharides from garlic. Carbohydr. Polym. 2014, 111, 606–609. [Google Scholar] [CrossRef]
- Yang, W.G.; Xie, G.H.; Xu, D.L.; Zhu, Q.L.; Lu, J.F.; Zhou, X.Y. Degradation of Gracilaria lemaneiformis polysaccharide and antioxidation of its degraded products. J. Fish. China 2009, 33, 342–347. [Google Scholar]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Ji, D.; You, L.; Ren, Y.; Wen, L.; Zheng, G.; Li, C. Protective effect of polysaccharides from Sargassum fusiforme against UVB-induced oxidative stress in HaCaT human keratinocytes. J. Funct. Foods 2017, 36, 332–340. [Google Scholar] [CrossRef]
- Liu, Z.; Li, G.; Long, C.; Xu, J.; Cen, J.; Yang, X. The antioxidant activity and genotoxicity of isogarcinol. Food Chem. 2018, 253, 5–12. [Google Scholar] [CrossRef]
- Fatma, G.; Bahri, G.; Beyzagül, E.; Zekai, H.; Akar, K. Investigation of serum and brain superoxide dismutase levels depending on atomoxetine used in attention-deficit/hyperactivity disorder treatment: A combination of in vivo and molecular docking studies. Bioorg. Chem. 2020, 105, 104435. [Google Scholar]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Ruth, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012, 645460. [Google Scholar] [CrossRef]
- Luo, X.; Zhang, H.; Duan, Y.; Chen, G. Protective effects of radish (Raphanus sativus L.) leaves extract against hydrogen peroxide-induced oxidative damage in human fetal lung fibroblast (MRC-5) cells. Biomed. Pharmacother. 2018, 103, 406–414. [Google Scholar] [CrossRef]
- Bresolí-Obach, R.; Busto-Moner, L.; Muller, L.; Reina, M.; Nonell, S. NanoDCFH-DA: A silica-based nanostructured fluorogenic probe for the detection of reactive oxygen species. Photochem. Photobiol. 2018, 94, 1143–1150. [Google Scholar] [CrossRef]
- He, R.R.; Tsoi, B.; Lan, F.; Yao, N.; Yao, X.S.; Kurihara, H. Antioxidant properties of lutein contribute to the protection against lipopolysaccharide-induced uveitis in mice. Chin. Med. 2011, 6, 38. [Google Scholar] [CrossRef]
- Jeon, G.; Kim, C.; Cho, U.M.; Hwang, E.T.; Hwang, H.S.; Min, J. Melanin-decolorizing activity of antioxidant enzymes, glutathione peroxidase, thiol peroxidase, and catalase. Mol. Biotechnol. 2021, 63, 150–155. [Google Scholar] [CrossRef]
- Fuhrmann, D.C.; Brüne, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, J.; Zhu, P.J.; Zhu, H.; Sam, T.; Hu, S.Y.; Ren, J.; Chen, Y.D. NR4A1 aggravates the cardiac microvascular ischemia reperfusion injury through suppressing FUNDC1-mediated mitophagy and promoting Mff-required mitochondrial fission by CK2α. Basic Res. Cardiol. 2018, 113, 23. [Google Scholar] [CrossRef]
- Deng, H.; Jia, Y.; Pan, D.; Ma, Z.G. Berberine alleviates rotenone-induced cytotoxicity by antioxidation and activation of PI3K/Akt signaling pathway in SH-SY5Y cells. Neuroreport 2020, 31, 41–47. [Google Scholar] [CrossRef]
- Kumari, A.; Kakkar, P. Lupeol prevents acetaminophen-induced in vivo hepatotoxicity by altering the Bax/Bcl-2 and oxidative stress-mediated mitochondrial signaling cascade. Life Sci. 2012, 90, 561–570. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lloyd, A.G.; Dodgson, K.S.; Price, R.G.; Rose, F.A. Infrared studies on sulphate esters. i. polysaccharide sulphates. Biochim. Biophys. Acta 1961, 46, 116–120. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, W.; Song, W.; Chen, H.; Teng, A.; Liu, A. Partial characterization and anti-tumor activity of an acidic polysaccharide from Gracilaria lemaneiformis. Carbohydr. Polym. 2012, 88, 1313–1318. [Google Scholar] [CrossRef]
- Hu, W.W.; Ye, X.Q.; Chantapakul, T.; Chen, S.G.; Zheng, J.Q. Manosonication extraction of RG-I pectic polysaccharides from citrus waste: Optimization and kinetics analysis—Sciencedirect. Carbohydr. Polym. 2020, 235, 115982. [Google Scholar] [CrossRef]
- Ren, Y.; Zheng, G.; You, L.; Wen, L.; Li, C.; Fu, X.; Zhou, L. Structural characterization and macrophage immunomodulatory activity of a polysaccharide isolated from Gracilaria lemaneiformis. J. Funct. Foods 2017, 33, 286–296. [Google Scholar] [CrossRef]
- Yang, W.; Wang, Y.; Li, X.; Yu, P. Purification and structural characterization of Chinese yam polysaccharide and its activities. Carbohydr. Polym. 2015, 117, 1021–1027. [Google Scholar] [CrossRef]
- You, L.J.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Sugar/% | Sulfate/% | Uronic Acid/% | Mw/kDa |
|---|---|---|---|---|
| GLP | 69.33 ± 0.61 | 18.71 ± 1.31 | 5.55 ± 0.20 | 106 |
| GPP | 74.29 ± 1.48 * | 13.91 ± 0.89 ** | 6.40 ± 0.08 ** | 66 |
| GGP | 78.21 ± 4.21 ** | 18.91 ± 1.12 | 5.32 ± 0.38 | 93 |
| GCP | 74.29 ± 2.86 * | 4.56 ± 0.48 ** | 4.13 ± 0.09 ** | 89 |
| GXP | 72.68 ± 2.35 | 7.33 ± 0.10 ** | 6.01 ± 0.11 * | 95 |
| GDP | 79.13 ± 0.91 ** | 10.11 ± 0.56 ** | 5.46 ± 0.14 | 81 |
| Monosaccharide Composition (mol %) | GLP | GPP | GGP | GCP | GXP | GDP |
|---|---|---|---|---|---|---|
| Fucose (Fuc) | - | - | - | - | 0.2 | - |
| Rhamnose (Rha) | - | 1.4 | 1.2 | - | - | 0.7 |
| Arabinose (Ara) | - | 0.2 | 0.5 | - | - | 0.1 |
| Galactose (Gal) | 89.7 | 58.8 | 68.0 | 20.0 | 70.9 | 41.9 |
| Glucose (Glu) | 2.3 | 33 | 13.8 | 74.5 | 23.4 | 42.1 |
| Xylose (Xyl) | 1.2 | 1.5 | 1.0 | 0.1 | 0.6 | 0.8 |
| Mannose (Man) | - | - | 10.9 | 4.2 | 0.8 | 11.1 |
| Galacturonic acid (GalA) | 3.4 | 2.2 | 2.6 | 0.4 | 1.6 | 1.3 |
| Glucuronic acid (GluA) | 3.4 | 2.1 | 1.3 | 0.6 | 1.4 | 0.6 |
| Mannuronic acid (ManA) | - | - | - | - | 0.7 | 0.5 |
| Number | A (Temperature/°C) | B (pH) | C (Enzyme Dosage/U·g−1) | Hydroxyl Radical Scavenging Rate/% | DPPH Radical Scavenging Rate/% | Reducing Capacity | Total Antioxidant Capacity | Comprehensive Score | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1(30) | 1(4.4) | 1(250,000) | 48.33 ± 0.26 | 29.60 ± 0.38 | 0.096 ± 0.002 | 0.322 ± 0.002 | 19.59 | |
| 2 | 1(30) | 2(5.0) | 2(300,000) | 44.22 ± 0.85 | 28.42 ± 0.14 | 0.108 ± 0.002 | 0.329 ± 0.001 | 18.27 | |
| 3 | 1(30) | 3(5.6) | 3(350,000) | 35.65 ± 0.64 | 28.79 ± 0.38 | 0.104 ± 0.001 | 0.320 ± 0.001 | 16.22 | |
| 4 | 2(40) | 1(4.4) | 2(300,000) | 43.39 ± 1.43 | 19.65 ± 1.08 | 0.109 ± 0.004 | 0.324 ± 0.001 | 15.87 | |
| 5 | 2(40) | 2(5.0) | 3(350,000) | 40.27 ± 2.71 | 20.74 ± 0.05 | 0.118 ± 0.004 | 0.326 ± 0.001 | 15.36 | |
| 6 | 2(40) | 3(5.6) | 1(250,000) | 38.92 ± 0.17 | 22.45 ± 0.35 | 0.117 ± 0.001 | 0.324 ± 0.001 | 15.45 | |
| 7 | 3(50) | 1(4.4) | 3(350,000) | 37.12 ± 1.13 | 22.23 ± 0.11 | 0.124 ± 0.003 | 0.322 ± 0.001 | 14.95 | |
| 8 | 3(50) | 2(5.0) | 1(250,000) | 41.94 ± 0.16 | 17.69 ± 0.53 | 0.117 ± 0.003 | 0.322 ± 0.001 | 15.02 | |
| 9 | 3(50) | 3(5.6) | 2(300,000) | 39.30 ± 0.61 | 15.92 ± 0.14 | 0.124 ± 0.006 | 0.322 ± 0.001 | 13.92 | |
| Hydroxyl radical scavenging rate/% | K1 | 42.73 | 42.95 | 43.06 | |||||
| K2 | 40.86 | 42.14 | 42.30 | ||||||
| K3 | 39.45 | 37.96 | 37.68 | ||||||
| R | 3.28 | 4.99 | 5.38 | ||||||
| DPPH radical scavenging rate/% | K1 | 28.94 | 23.83 | 23.25 | |||||
| K2 | 20.95 | 22.28 | 21.33 | ||||||
| K3 | 18.61 | 22.39 | 23.92 | ||||||
| R | 10.32 | 1.54 | 2.59 | ||||||
| Reducing capacity | K1 | 0.103 | 0.110 | 0.110 | |||||
| K2 | 0.115 | 0.114 | 0.114 | ||||||
| K3 | 0.122 | 0.115 | 0.115 | ||||||
| R | 0.019 | 0.005 | 0.005 | ||||||
| Total antioxidant capacity | K1 | 0.324 | 0.323 | 0.323 | |||||
| K2 | 0.325 | 0.326 | 0.325 | ||||||
| K3 | 0.322 | 0.322 | 0.323 | ||||||
| R | 0.003 | 0.004 | 0.002 | ||||||
| Comprehensive score | K1 | 18.03 | 16.80 | 16.69 | |||||
| K2 | 15.56 | 16.22 | 16.02 | ||||||
| K3 | 14.63 | 15.20 | 15.51 | ||||||
| R | 3.40 | 1.61 | 1.18 | ||||||
| Samples | Scheme | Hydroxyl Radical Scavenging Rate/% | DPPH Radical Scavenging Rate/% | Reducing Capacity | Total Antioxidant Capacity |
|---|---|---|---|---|---|
| GDP1 | A1B1C1 | 52.20 ± 0.82b | 29.73 ± 0.71b | 0.117 ± 0.002c | 0.325 ± 0.001d |
| GDP2 | A1B1C3 | 39.92 ± 0.63d | 32.68 ± 0.75a | 0.125 ± 0.004b | 0.334 ± 0.002c |
| GDP3 | A2B2C2 | 61.99 ± 2.44a | 24.28 ± 1.08d | 0.129 ± 0.002b | 0.345 ± 0.002b |
| GDP4 | A3B3C3 | 44.27 ± 0.16c | 26.15 ± 0.23c | 0.140 ± 0.001a | 0.355 ± 0.002a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, T.; Zhang, X.; Hu, S.; Yu, Y.; Sun, X.; Xu, N. Enzymatic Degradation of Gracilariopsis lemaneiformis Polysaccharide and the Antioxidant Activity of Its Degradation Products. Mar. Drugs 2021, 19, 270. https://doi.org/10.3390/md19050270
Fang T, Zhang X, Hu S, Yu Y, Sun X, Xu N. Enzymatic Degradation of Gracilariopsis lemaneiformis Polysaccharide and the Antioxidant Activity of Its Degradation Products. Marine Drugs. 2021; 19(5):270. https://doi.org/10.3390/md19050270
Chicago/Turabian StyleFang, Tian, Xiaoqian Zhang, Shanshan Hu, Yanyan Yu, Xue Sun, and Nianjun Xu. 2021. "Enzymatic Degradation of Gracilariopsis lemaneiformis Polysaccharide and the Antioxidant Activity of Its Degradation Products" Marine Drugs 19, no. 5: 270. https://doi.org/10.3390/md19050270
APA StyleFang, T., Zhang, X., Hu, S., Yu, Y., Sun, X., & Xu, N. (2021). Enzymatic Degradation of Gracilariopsis lemaneiformis Polysaccharide and the Antioxidant Activity of Its Degradation Products. Marine Drugs, 19(5), 270. https://doi.org/10.3390/md19050270