
Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity

Abstract
1. Introduction
2. Results
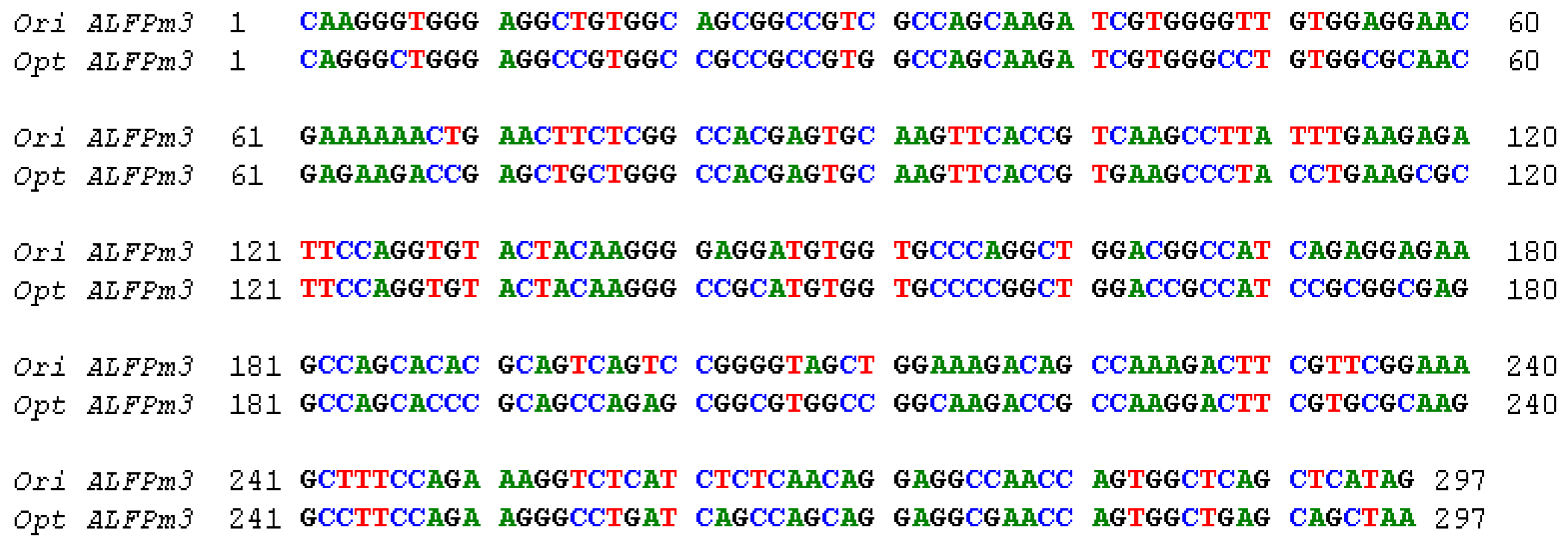
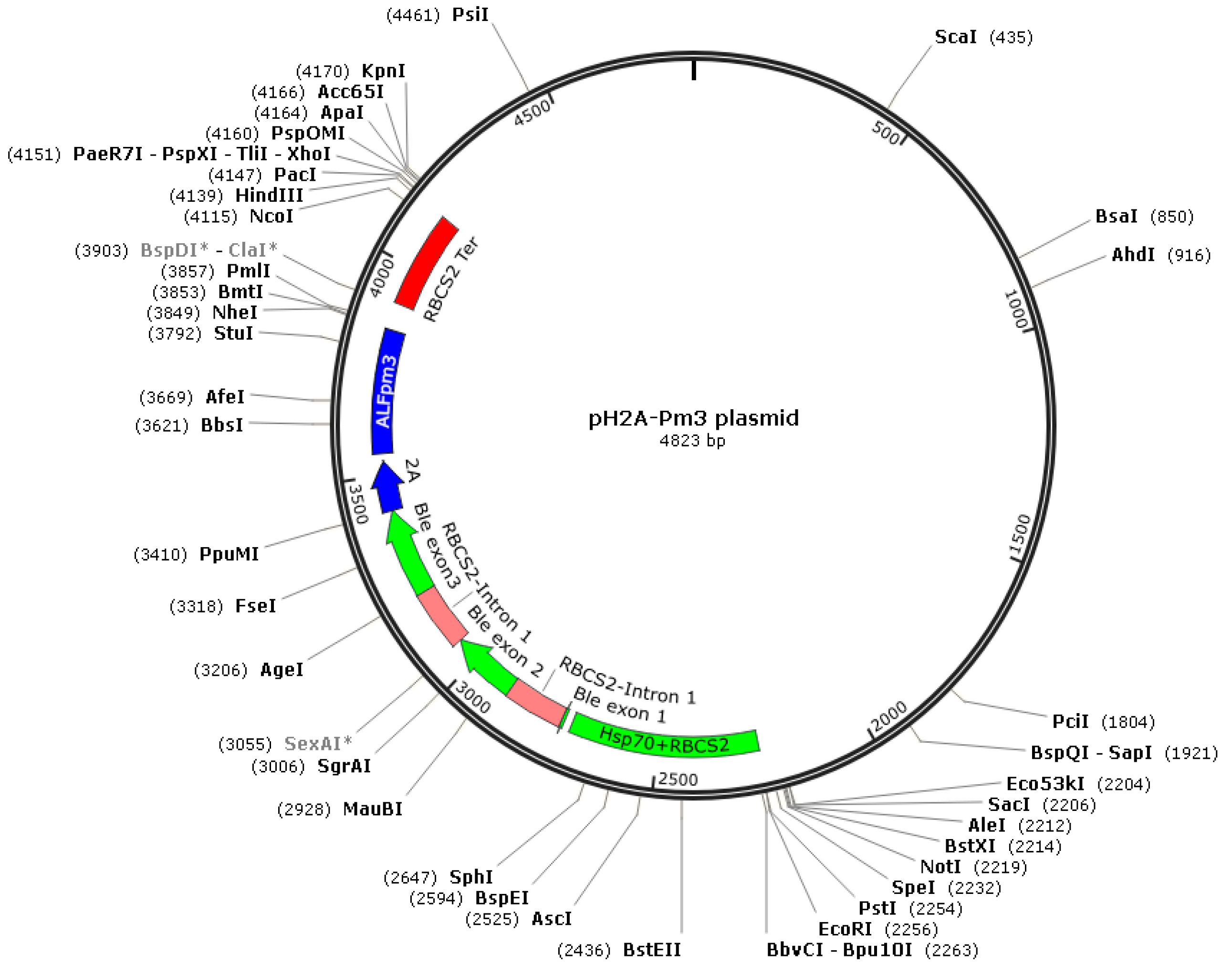
2.1. The Design of ALFPm3 Expression Cassette
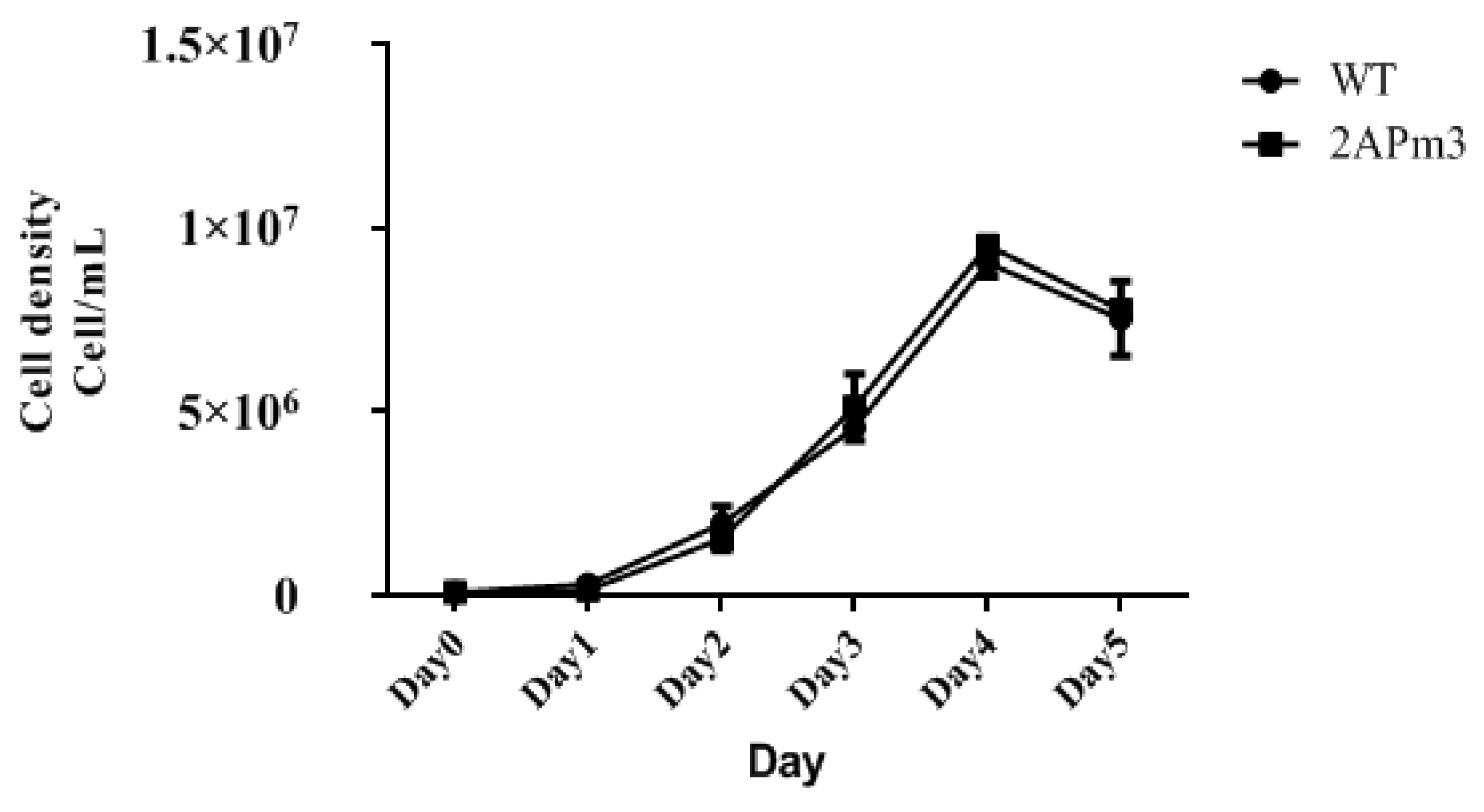
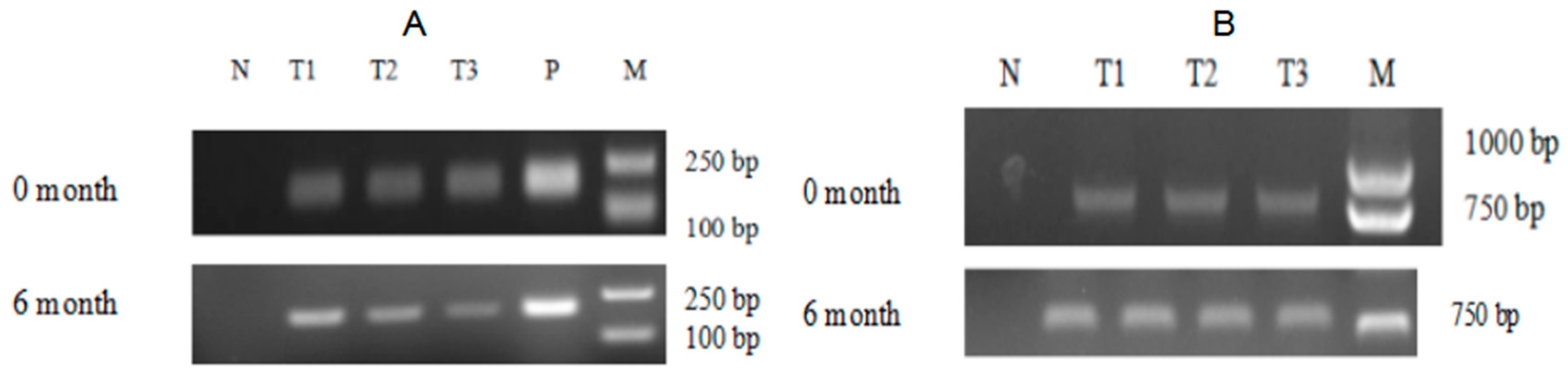
2.2. The Screening of Transgenic C. reinhardtii
2.3. The Analysis of ALFPm3 Protein Production in Transgenic C. reinhardtii
2.4. The Expression Analysis of ALFPm3 Gene in C. reinhardtii
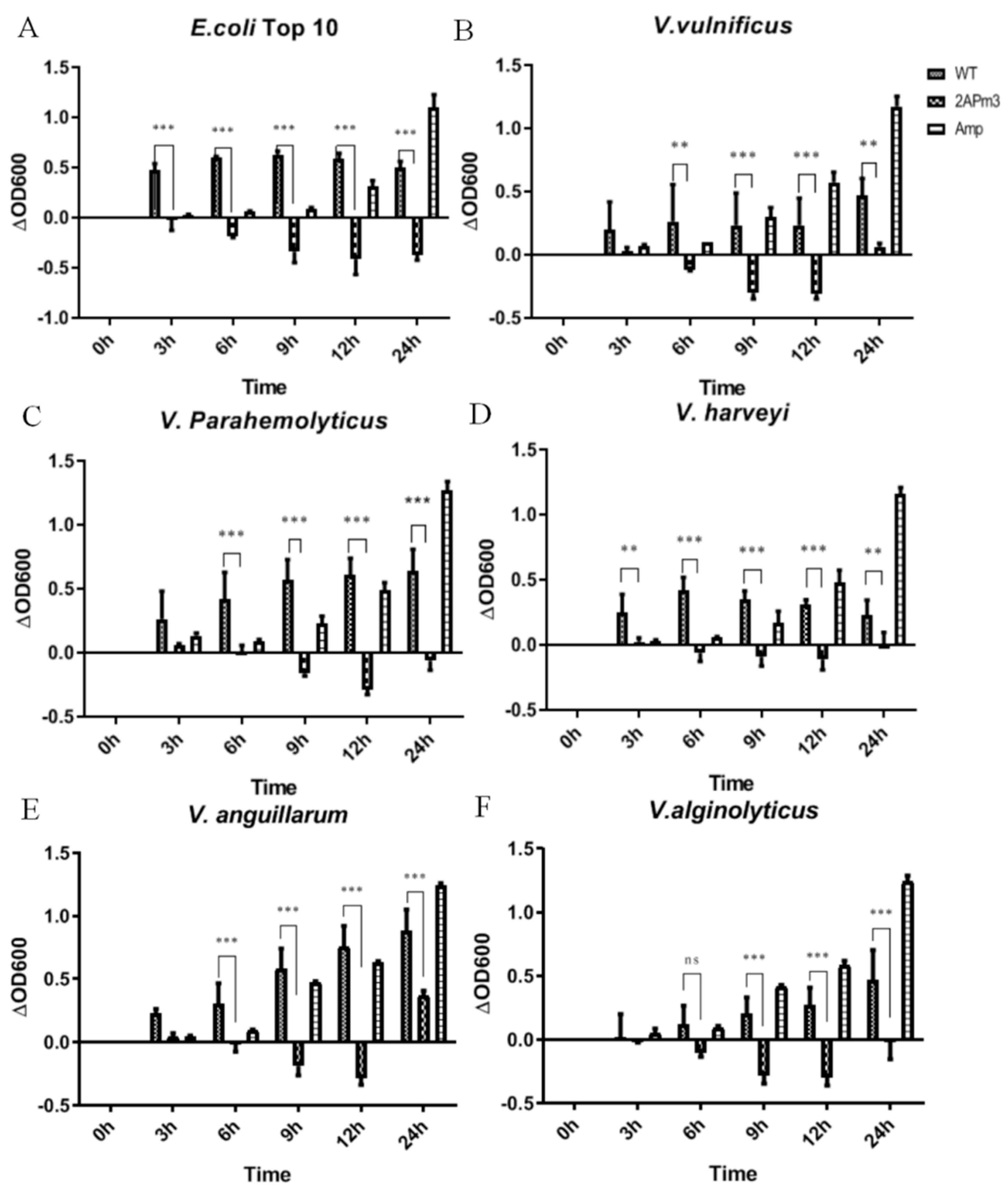
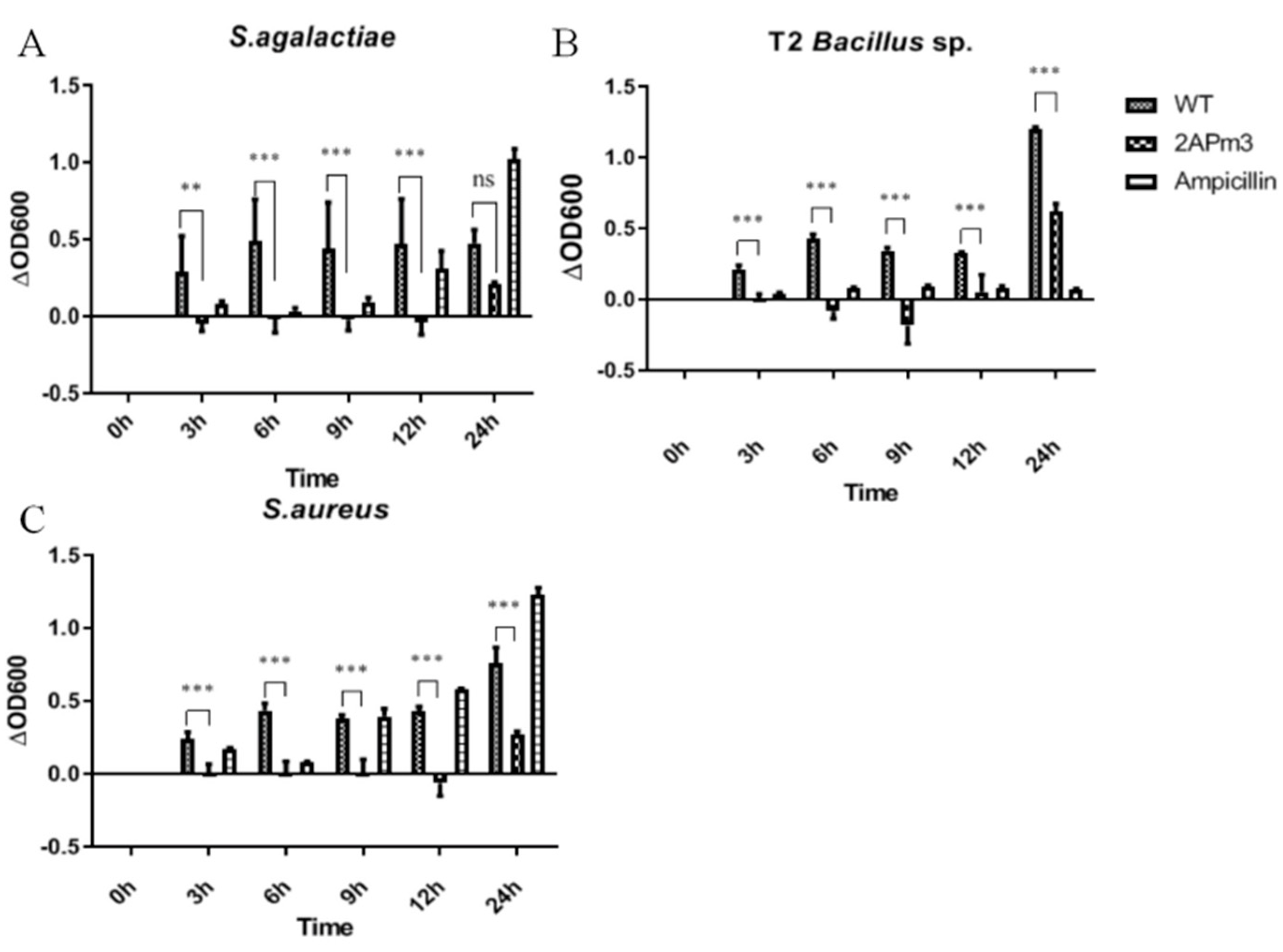
2.5. cALFPm3 Showed High Anti-Bacterial Activity
3. Discussion
4. Materials and Methods
4.1. Algal Strain and Culture Conditions
4.2. Plasmid Construction and Genetic Transformation
4.3. Genomic PCR and RT-PCR Analysis
4.4. Protein Extraction and Immunoblot Analysis
4.5. Anti-Bacterial Assay
4.6. Expression Analysis of ALFPm3 in C. reinhardtii Transformants
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- López-Montesinos, I.; Domínguez-Guasch, A.; Gómez-Zorrilla, S.; Duran-Jordà, X.; Siverio-Parès, A.; Arenas-Miras, M.; Montero, M.; Redó, L.S.; Grau, S.; Horcajada, J.; et al. Clinical and economic burden of community-onset multidrug-resistant infections requiring hospitalization. J. Infect. 2020, 80, 271–278. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Bhopale, G.M. Antimicrobial Peptides: A Promising Avenue for Human Healthcare. Curr. Pharm. Biotechnol. 2020, 21, 90–96. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms: My Perspective. Adv. Exp. Med. Biol. 2019, 1117, 3–6. [Google Scholar] [CrossRef]
- Yi, H.-Y.; Chowdhury, M.; Huang, Y.-D.; Yu, X.-Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Andreev, Y.A.; Odintsova, T.I. Hevein-like antimicrobial peptides of plants. Biochemistry 2017, 82, 1659–1674. [Google Scholar] [CrossRef]
- Chen, X.; Yi, Y.; Bian, C.; You, X.; Shi, Q. Putative Antimicrobial Peptides in Fish: Using Zebrafish as a Representative. Protein Pept. Lett. 2020, 27, 1059–1067. [Google Scholar] [CrossRef]
- Balandin, S.V.; Sheremeteva, E.V.; Ovchinnikova, T.V. Pediocin-Like Antimicrobial Peptides of Bacteria. Biochemistry 2019, 84, 464–478. [Google Scholar] [CrossRef]
- Raeder, S.B.; Sandbakken, E.T.; Nepal, A.; Løseth, K.; Bergh, K.; Witsø, E.; Otterlei, M. Novel Peptides Targeting the β-Clamp Rapidly Kill Planktonic and Biofilm Staphylococcus epidermidis Both in vitro and in vivo. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Mar. Drugs 2021, 19, 176. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Tanaka, S.; Nakamura, T.; Morita, T.; Iwanaga, S. Limulus anti-LPS factor: An anticoagulant which inhibits the endotoxin-mediated activation of Limulus coagulation system. Biochem. Biophys. Res. Commun. 1982, 105, 717–723. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Jiao, Y.; Huang, D.; Li, A.; Chen, H.; Liu, Y.; Li, S.; Li, H.; Wang, C. Molecular and antimicrobial characterization of a group G anti-lipopolysaccharide factor (ALF) from Penaeus monodon. Fish. Shellfish. Immunol. 2019, 94, 149–156. [Google Scholar] [CrossRef]
- Soonthornchai, W.; Chaiyapechara, S.; Klinbunga, S.; Thongda, W.; Tangphatsornruang, S.; Yoocha, T.; Jarayabhand, P.; Jiravanichpaisal, P. Differentially expressed transcripts in stomach of Penaeus monodon in response to AHPND infection. Dev. Comp. Immunol. 2016, 65, 53–63. [Google Scholar] [CrossRef]
- Tharntada, S.; Somboonwiwat, K.; Rimphanitchayakit, V.; Tassanakajon, A. Anti-lipopolysaccharide factors from the black tiger shrimp, Penaeus monodon, are encoded by two genomic loci. Fish. Shellfish. Immunol. 2008, 24, 46–54. [Google Scholar] [CrossRef]
- Methatham, T.; Boonchuen, P.; Jaree, P.; Tassanakajon, A.; Somboonwiwat, K. Antiviral action of the antimicrobial peptide ALFPm3 from Penaeus monodon against white spot syndrome virus. Dev. Comp. Immunol. 2017, 69, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Jaree, P.; Tassanakajon, A.; Somboonwiwat, K. Effect of the anti-lipopolysaccharide factor isoform 3 (ALFPm3) from Penaeus monodon on Vibrio harveyi cells. Dev. Comp. Immunol. 2012, 38, 554–560. [Google Scholar] [CrossRef]
- Ponprateep, S.; Tharntada, S.; Somboonwiwat, K.; Tassanakajon, A. Gene silencing reveals a crucial role for anti-lipopolysaccharide factors from Penaeus monodon in the protection against microbial infections. Fish. Shellfish. Immunol. 2012, 32, 26–34. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Li, A.; Jiao, Y.; Li, S.; Huang, J.; Yang, L.; Wang, C. Characterization of a group D anti-lipopolysaccharide factor (ALF) involved in anti-Vibrio response in Penaeus monodon. Fish. Shellfish. Immunol. 2019, 89, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.-L.; Wei, Y.; Wang, Z.-Z. Functional expression, purification, and antimicrobial activity of a novel antimicrobial peptide MLH in Escherichia coli. Prep. Biochem. Biotechnol. 2018, 48, 57–63. [Google Scholar] [CrossRef]
- Corrales-García, L.L.; Serrano-Carreón, L.; Corzo, G. Improving the heterologous expression of human β-defensin 2 (HBD2) using an experimental design. Protein Expr. Purif. 2020, 167, 105539. [Google Scholar] [CrossRef]
- Yi, T.; Sun, S.; Huang, Y.; Chen, Y. Prokaryotic expression and mechanism of action of α-helical antimicrobial peptide A20L using fusion tags. BMC Biotechnol. 2015, 15, 69. [Google Scholar] [CrossRef]
- Ishida, H.; Nguyen, L.T.; Gopal, R.; Aizawa, T.; Vogel, H.J. Overexpression of Antimicrobial, Anticancer, and Transmembrane Peptides inEscherichia colithrough a Calmodulin-Peptide Fusion System. J. Am. Chem. Soc. 2016, 138, 11318–11326. [Google Scholar] [CrossRef]
- Su, G.; Tang, F.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; et al. Expression, Purification and Characterization of a Novel Antimicrobial Peptide: Gloverin A2 from Bombyx mori. Int. J. Pept. Res. Ther. 2019, 25, 827–833. [Google Scholar] [CrossRef]
- Cao, J.; De La Fuente-Nunez, C.; Ou, R.W.; Torres, M.D.T.; Pande, S.G.; Sinskey, A.J.; Lu, T.K. Yeast-Based Synthetic Biology Platform for Antimicrobial Peptide Production. ACS Synth. Biol. 2018, 7, 896–902. [Google Scholar] [CrossRef]
- Li, X.; Fan, Y.; Lin, Q.; Luo, J.; Huang, Y.; Bao, Y.; Xu, L. Expression of chromogranin A-derived antifungal peptide CGA-N12 in Pichia pastoris. Bioengineered 2020, 11, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Klinbunga, S.; Aumelas, A.; Romestand, B.; Gueguen, Y.; Boze, H.; Moulin, G.; Bachère, E. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef]
- Tang, T.; Liu, J.; Li, S.; Li, H.; Liu, F. Recombinant expression of an oriental river prawn anti-lipopolysaccharide factor gene in Pichia pastoris and its characteristic analysis. Fish. Shellfish. Immunol. 2020, 98, 414–419. [Google Scholar] [CrossRef]
- Prabhu, A.A.; Boro, B.; Bharali, B.; Chakraborty, S.; Dasu, V.V. Gene and process level modulation to overcome the bottlenecks of recombinant proteins expression in Pichia pastoris. Curr. Pharm. Biotechnol. 2018, 18, 1200–1223. [Google Scholar] [CrossRef]
- Puxbaum, V.; Mattanovich, D.; Gasser, B. Quo vadis? The challenges of recombinant protein folding and secretion in Pichia pastoris. Appl. Microbiol. Biotechnol. 2015, 99, 2925–2938. [Google Scholar] [CrossRef]
- Rabert, C.; Weinacker, D., Jr.; Pessoa, A.; Farías, J.G. Recombinants proteins for industrial uses: Utilization of Pichia pastoris expression system. Braz. J. Microbiol. 2013, 44, 351–356. [Google Scholar] [CrossRef]
- Zhang, M.-P.; Wang, M.; Wang, C. Nuclear transformation of Chlamydomonas reinhardtii: A review. Biochimie 2021, 181, 1–11. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kwon, S.G.; Bae, S.J.; Park, S.J.; Im, J. Optimization of the droplet electroporation method for wild type Chlamydomonas reinhardtii transformation. Bioelectrochemistry 2019, 126, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Purton, S. Tools and Techniques for Chloroplast Transformation of Chlamydomonas. Adv. Exp. Med. Biol. 2007, 616, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Vargas, J.M.; Jiménez-Clemente, L.A.; Macedo-Osorio, K.S.; Oliver-Salvador, M.C.; Fernández-Linares, L.C.; Durán-Figueroa, N.V.; Badillo-Corona, J.A. Use of the ptxD gene as a portable selectable marker for chloroplast transformation in Chlamydomonas reinhardtii. Mol. Biotechnol. 2019, 61, 461–468. [Google Scholar] [CrossRef]
- LaRosa, V.; Remacle, C. Transformation of the mitochondrial genome. Int. J. Dev. Biol. 2013, 57, 659–665. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef]
- Rasala, B.A.; Lee, P.A.; Shen, Z.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust Expression and Secretion of Xylanase1 in Chlamydomonas reinhardtii by Fusion to a Selection Gene and Processing with the FMDV 2A Peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef] [PubMed]
- Soria-Guerra, R.E.; Ramírez-Alonso, J.I.; Ibáñez-Salazar, A.; Govea-Alonso, D.O.; Paz-Maldonado, L.M.T.; Bañuelos-Hernández, B.; Korban, S.S.; Rosales-Mendoza, S. Expression of an HBcAg-based antigen carrying angiotensin II in Chlamydomonas reinhardtii as a candidate hypertension vaccine. Plant Cell Tissue Organ Cult. 2014, 116, 133–139. [Google Scholar] [CrossRef]
- Gregory, J.A.; Li, F.; Tomosada, L.M.; Cox, C.J.; Topol, A.B.; Vinetz, J.M.; Mayfield, S. Algae-Produced Pfs25 Elicits Antibodies That Inhibit Malaria Transmission. PLoS ONE 2012, 7, e37179. [Google Scholar] [CrossRef] [PubMed]
- Barahimipour, R.; Neupert, J.; Bock, R. Efficient expression of nuclear transgenes in the green alga Chlamydomonas: Synthesis of an HIV antigen and development of a new selectable marker. Plant. Mol. Biol. 2016, 90, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.-C.; Liang, M.-H.; Jiang, J.-G. Transgenic microalgae as bioreactors. Crit. Rev. Food Sci. Nutr. 2020, 60, 3195–3213. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. The microalgaChlamydomonas reinhardtiias a platform for the production of human protein therapeutics. Bioeng. Bugs 2011, 2, 50–54. [Google Scholar] [CrossRef]
- Schroda, M. Good News for Nuclear Transgene Expression in Chlamydomonas. Cells 2019, 8, 1534. [Google Scholar] [CrossRef]
- De Felipe, P.; Izquierdo, M. Construction and characterization of pentacistronic retrovirus vectors. J. Gen. Virol. 2003, 84, 1281–1285. [Google Scholar] [CrossRef]
- Agca, C.; Seye, C.; Benson, C.M.K.; Rikka, S.; Chan, A.W.; Weisman, G.A.; Agca, Y. Development of a Novel Transgenic Rat Overexpressing the P2Y2 Nucleotide Receptor Using a Lentiviral Vector. J. Vasc. Res. 2009, 46, 447–458. [Google Scholar] [CrossRef]
- Rasala, B.A.; Barrera, D.J.; Ng, J.; Plucinak, T.M.; Rosenberg, J.N.; Weeks, N.P.; Oyler, G.A.; Peterson, T.C.; Haerizadeh, F.; Mayfield, S.P. Expanding the spectral palette of fluorescent proteins for the green microalgaChlamydomonas reinhardtii. Plant. J. 2013, 74, 545–556. [Google Scholar] [CrossRef]
- Dong, B.; Hu, H.-H.; Li, Z.-F.; Cheng, R.-Q.; Meng, D.-M.; Wang, J.; Fan, Z.-C. A novel bicistronic expression system composed of the intraflagellar transport protein gene ift25 and FMDV 2A sequence directs robust nuclear gene expression in Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2017, 101, 4227–4245. [Google Scholar] [CrossRef] [PubMed]
- Plucinak, T.M.; Horken, K.M.; Jiang, W.; Fostvedt, J.; Nguyen, S.T.; Weeks, D.P. Improved and versatile viral 2A platforms for dependable and inducible high-level expression of dicistronic nuclear genes in Chlamydomonas reinhardtii. Plant. J. 2015, 82, 717–729. [Google Scholar] [CrossRef]
- Dong, B.; Cheng, R.-Q.; Liu, Q.-Y.; Wang, J.; Fan, Z.-C. Multimer of the antimicrobial peptide Mytichitin-A expressed in Chlamydomonas reinhardtii exerts a broader antibacterial spectrum and increased potency. J. Biosci. Bioeng. 2018, 125, 175–179. [Google Scholar] [CrossRef]
- Neupert, J.; Shao, N.; Lu, Y.; Bock, R. Genetic Transformation of the Model Green Alga Chlamydomonas reinhardtii. In Advanced Structural Safety Studies; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2012; Volume 847, pp. 35–47. [Google Scholar]
- Cao, M.; Fu, Y.; Guo, Y.; Pan, J. Chlamydomonas (Chlorophyceae) colony PCR. Protoplasma 2009, 235, 107–110. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Target Gene |
|---|---|---|
| Ble-F | TTAAATCTAGAAAAATGGCCAG | ble |
| Ble-R | GTCCTGCTCCTCGGCCACG | |
| ALF3-F | AAGTTCACCGTGAAGCCCTAC | ALFPm3 |
| ALF3-R | CTGCTCAGCCACTGGTTCGC | |
| actin-F | ACCCCGTGCTGCTGACTG | β-actin |
| actin-R | ACGTTGAAGGTCTCGAACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Huang, R.; Wang, C.; Hu, Q.; Li, H.; Li, X. Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity. Mar. Drugs 2021, 19, 239. https://doi.org/10.3390/md19050239
Li A, Huang R, Wang C, Hu Q, Li H, Li X. Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity. Marine Drugs. 2021; 19(5):239. https://doi.org/10.3390/md19050239
Chicago/Turabian StyleLi, Anguo, Ruihao Huang, Chaogang Wang, Qunju Hu, Hui Li, and Xiao Li. 2021. "Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity" Marine Drugs 19, no. 5: 239. https://doi.org/10.3390/md19050239
APA StyleLi, A., Huang, R., Wang, C., Hu, Q., Li, H., & Li, X. (2021). Expression of Anti-Lipopolysaccharide Factor Isoform 3 in Chlamydomonas reinhardtii Showing High Antimicrobial Activity. Marine Drugs, 19(5), 239. https://doi.org/10.3390/md19050239