Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction to Animal Venoms
2. Phylum Cnidaria
3. Venom Evolution across Cnidaria
4. Geographic, Ontogenetic and Prey-Associated Venom Variation
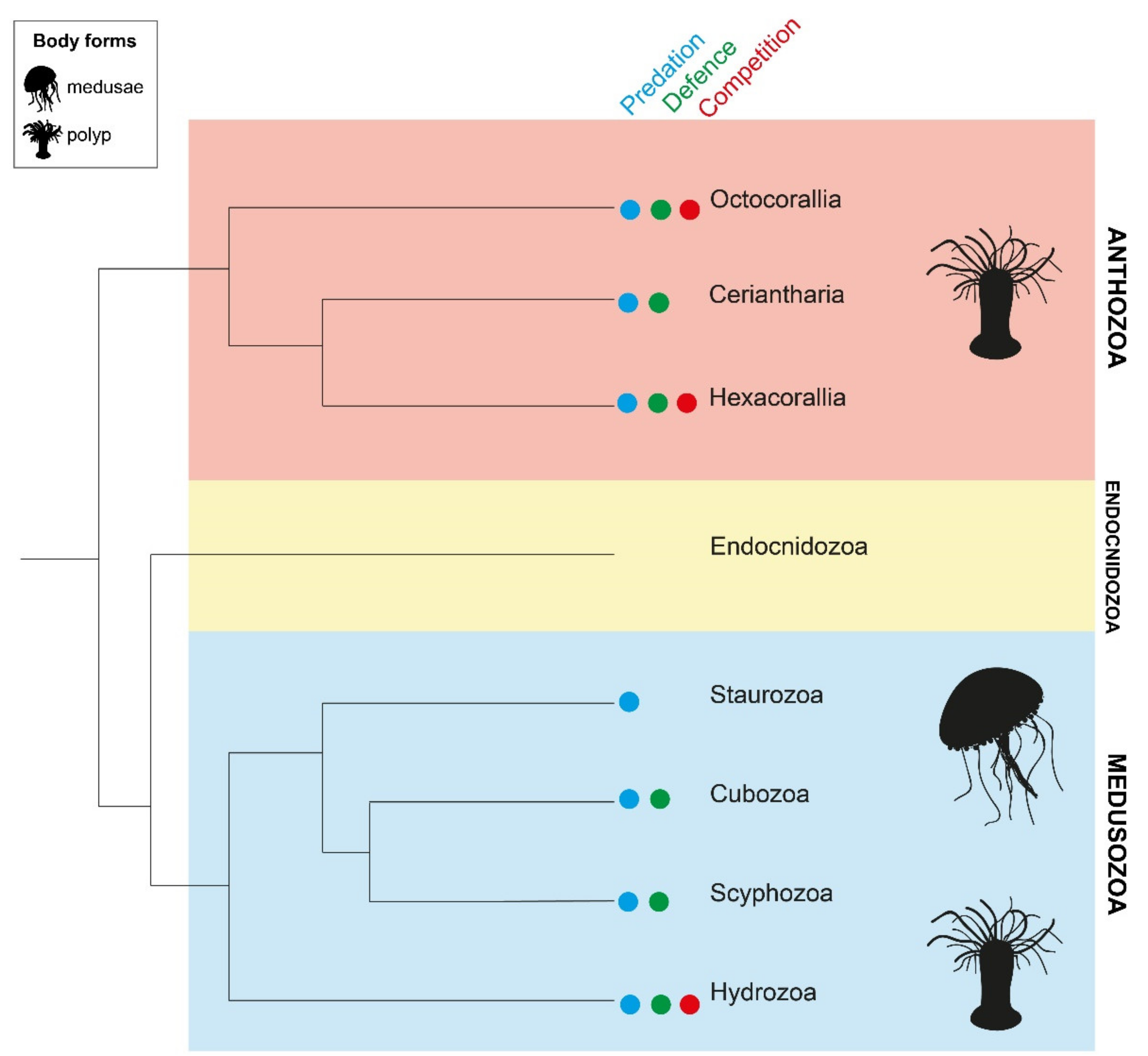
5. Colonial Regionalisation and Functional Divisions
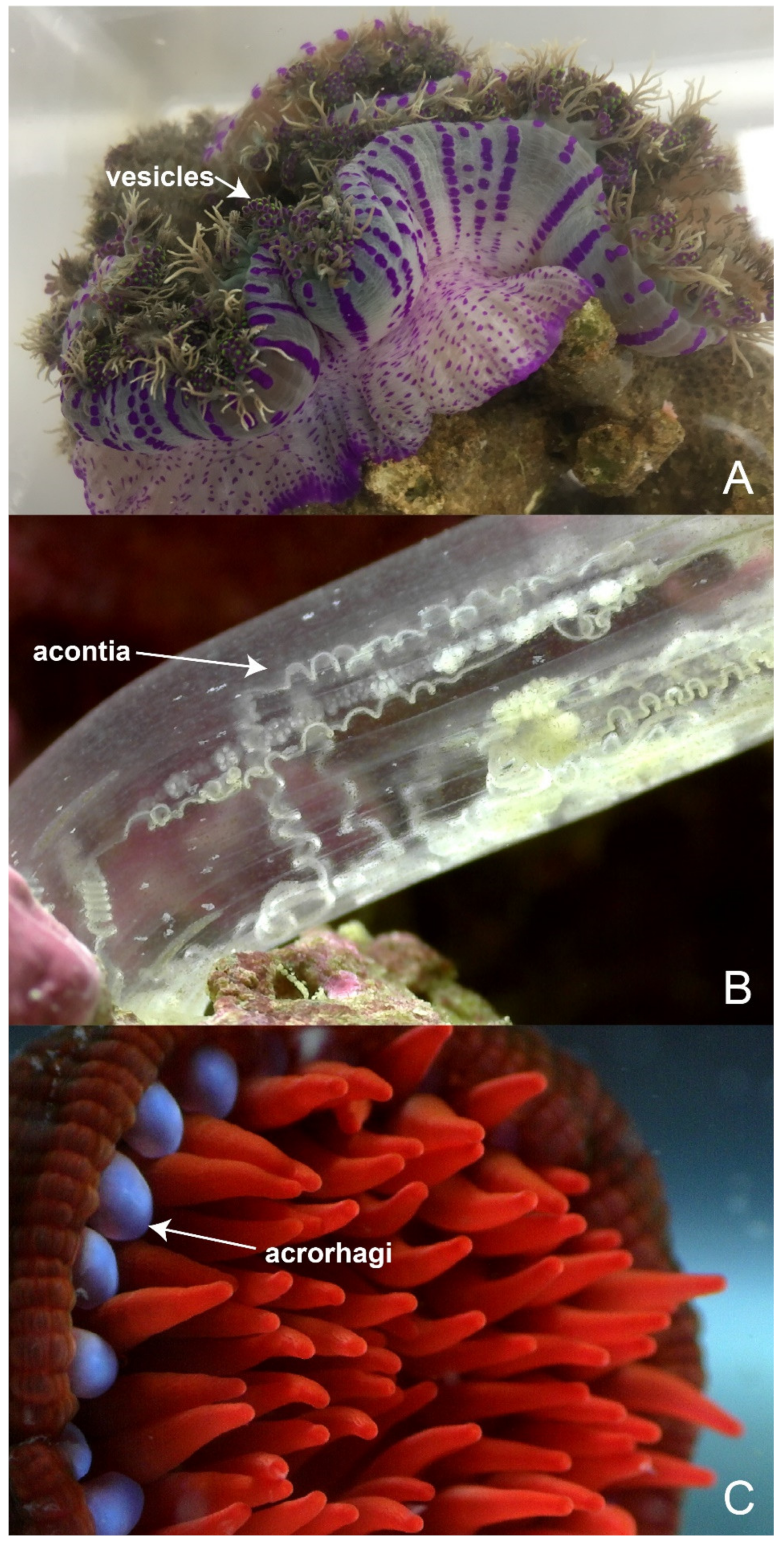
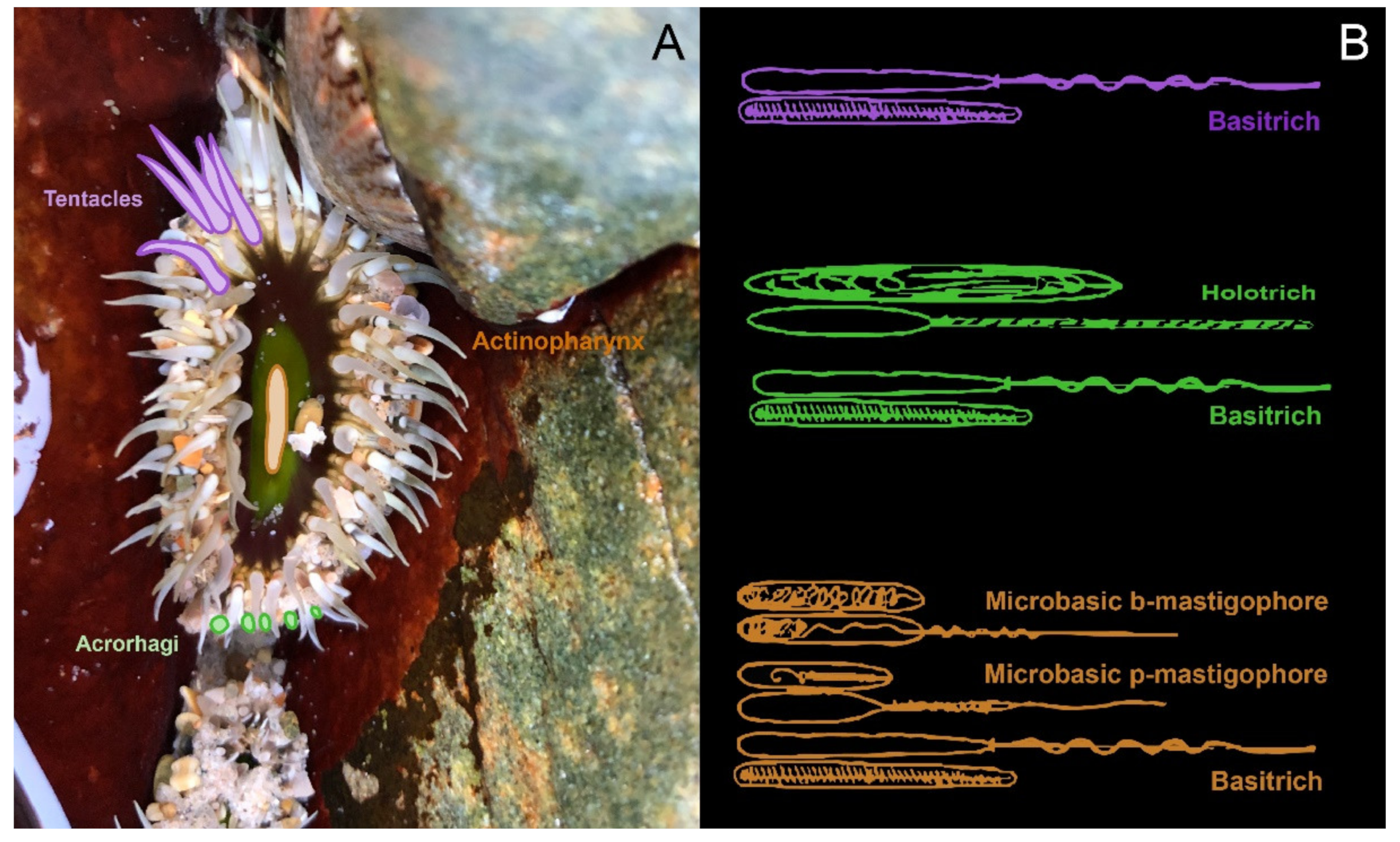
6. Shared and Specialised Morphology
7. Functional Anatomy and Venom Variation
8. Characterising Toxin Expression Patterns
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Paterson, S.; Vogwill, T.; Buckling, A.; Benmayor, R.; Spiers, A.; Thomson, N.; Quail, M.; Smith, F.; Walker, D.; Libberton, B.; et al. Antagonistic coevolution accelerates molecular evolution. Nature 2010, 464, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Langerhans, R.B. Evolutionary consequences of predation: avoidance, escape, reproduction, and diversification. In Predation in Organisms; Elewa, A.M.T., Ed.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, D.L.; Jenner, A.R.; Undheim, A.B.E. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Animal venom studies: Current benefits and future developments. World J. Biol. Chem. 2015, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Zhang, Y. Why do we study animal toxins? Zool. Res. 2015, 36, 183–222. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a platform for human drugs: translating toxins into therapeutics. Expert Opin. Biol. Th. 2011, 11, 1469–1484. [Google Scholar] [CrossRef]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discovery 2003, 2, 790–802. [Google Scholar] [CrossRef]
- Norton, R.S. Enhancing the therapeutic potential of peptide toxins. Expert Opin. Drug Discovery 2017, 12, 611–623. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: the evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Beress, L. Biologically active compounds from coelenterates. Pure Appl. Chem. 1982, 54, 1981–1994. [Google Scholar] [CrossRef]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef]
- Jenner, R.A.; von Reumont, B.M.; Campbell, L.I.; Undheim, E.A.B. Parallel Evolution of Complex Centipede Venoms Revealed by Comparative Proteotranscriptomic Analyses. Mol. Biol. Evol. 2019, 36, 2748–2763. [Google Scholar] [CrossRef]
- Surm, J.M.; Smith, H.L.; Madio, B.; Undheim, E.A.; King, G.F.; Hamilton, B.R.; Van der Burg, C.A.; Pavasovic, A.; Prentis, P.J. A process of convergent amplification and tissue-specific expression dominates the evolution of toxin and toxin-like genes in sea anemones. Mol. Ecol. 2019, 28, 2272–2289. [Google Scholar] [CrossRef]
- Fautin, D.G. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef]
- Church, J.E.; Hodgson, W.C. The pharmacological activity of fish venoms. Toxicon 2002, 40, 1083–1093. [Google Scholar] [CrossRef]
- Peiren, N.; Vanrobaeys, F.; de Graaf, D.C.; Devreese, B.; Van Beeumen, J.; Jacobs, F.J. The protein composition of honeybee venom reconsidered by a proteomic approach. Biochim. Biophys. Acta Proteins Proteom. 2005, 1752, 1–5. [Google Scholar] [CrossRef]
- de Graaf, D.C.; Aerts, M.; Danneels, E.; Devreese, B. Bee, wasp and ant venomics pave the way for a component-resolved diagnosis of sting allergy. J. Proteomics 2009, 72, 145–154. [Google Scholar] [CrossRef]
- Brodie, E.D. Toxins and venoms. Curr. Biol. 2009, 19, R931–R935. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef]
- Cecilia, A.P.; Kellee, M.M.; Joseph, R.S. Venom variation during prey capture by the cone snail, Conus textile. PLoS ONE 2014, 9, e98991. [Google Scholar] [CrossRef]
- Walker, A.A.; Mayhew, M.L.; Jin, J.; Herzig, V.; Undheim, E.A.B.; Sombke, A.; Fry, B.G.; Meritt, D.J.; King, G.F. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Madio, B.; Peigneur, S.; Chin, Y.K.; Hamilton, B.R.; Henriques, S.T.; Smith, J.J.; Cristofori-Armstrong, B.; Dekan, Z.; Boughton, B.A.; Alewood, P.F. PHAB toxins: a unique family of predatory sea anemone toxins evolving via intra-gene concerted evolution defines a new peptide fold. Cell Mol. Life Sci. 2018, 75, 4511–4524. [Google Scholar] [CrossRef]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: a review on cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef]
- Jungo, F.; Bairoch, A. Tox-Prot, the toxin protein annotation program of the Swiss-Prot protein knowledgebase. Toxicon 2005, 45, 293–301. [Google Scholar] [CrossRef]
- Watson, G.M.; Hessinger, D.A. Cnidocyte mechanoreceptors are tuned to the movements of swimming prey by chemoreceptors. Science 1989, 243, 1589–1591. [Google Scholar] [CrossRef]
- Kayal, E.; Bentlage, B.; Sabrina Pankey, M.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Shpirer, E.; Diamant, A.; Cartwright, P.; Huchon, D. A genome wide survey reveals multiple nematocyst-specific genes in Myxozoa. BMC Evol. Biol. 2018, 18, 138. [Google Scholar] [CrossRef]
- Sunagar, K.; Moran, Y. The Rise and Fall of an Evolutionary Innovation: Contrasting Strategies of Venom Evolution in Ancient and Young Animals. PLoS Genet 2015, 11, e1005596. [Google Scholar] [CrossRef]
- Daly, M. Functional and Genetic Diversity of Toxins in Sea Anemones. In Evolution of Venomous Animals and Their Toxins; Malhotra, A., Ed.; Springer: Dordrecht, The Netherlands, 2017; pp. 87–104. [Google Scholar] [CrossRef]
- Hines, D.E.; Pawlik, J.R. Assessing the antipredatory defensive strategies of Caribbean non-scleractinian zoantharians (Cnidaria): is the sting the only thing? Mar. Biol. 2012, 159, 389–398. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Lindquist, N. Hydroid defenses against predators: the importance of secondary metabolites versus nematocysts. Oecologia 2000, 124, 280–288. [Google Scholar] [CrossRef]
- Francis, L. Intraspecific aggression and its effect on the distribution of Anthopleura elegantissima and some related sea anemones. Biol. Bull. 1973, 144, 73–92. [Google Scholar] [CrossRef]
- Bigger, C.H. Interspecific and Intraspecific Acrorhagial Aggressive-Behavior among Sea-Anemones—A Recognition of Self and Not-Self. Biol. Bull. 1980, 159, 117–134. [Google Scholar] [CrossRef]
- Ayre, D. Inter-genotype aggression in the solitary sea anemone Actinia tenebrosa. Mar. Biol. 1982, 68, 199–205. [Google Scholar] [CrossRef]
- Lane, S.M.; Briffa, M. Immune function and the decision to deploy weapons during fights in the beadlet anemone. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef]
- Miranda, L.S.; Mills, C.E.; Hirano, Y.M.; Collins, A.G.; Marques, A.C. A review of the global diversity and natural history of stalked jellyfishes (Cnidaria, Staurozoa). Mar. Biodivers. 2018, 48, 1695–1714. [Google Scholar] [CrossRef]
- Rachamim, T.; Morgenstern, D.; Aharonovich, D.; Brekhman, V.; Lotan, T.; Sher, D. The dynamically evolving nematocyst content of an anthozoan, a scyphozoan, and a hydrozoan. Mol. Biol. Evol. 2015, 32, 740–753. [Google Scholar] [CrossRef]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) toxins: an overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef]
- Jaimes-Becerra, A.; Chung, R.; Morandini, A.C.; Weston, A.J.; Padilla, G.; Gacesa, R.; Ward, M.; Long, P.F.; Marques, A.C. Comparative proteomics reveals recruitment patterns of some protein families in the venoms of Cnidaria. Toxicon 2017, 137, 19–26. [Google Scholar] [CrossRef]
- Daly, M.; Chaudhuri, A.; Gusmão, L.; Rodríguez, E. Phylogenetic relationships among sea anemones (Cnidaria: Anthozoa: Actiniaria). Mol. Phylogenet. Evol. 2008, 48, 292–301. [Google Scholar] [CrossRef]
- Fautin, D.G.; Malarky, L.; Soberon, J. Latitudinal diversity of sea anemones (Cnidaria: Actiniaria). Biol. Bull. 2013, 224, 89–98. [Google Scholar] [CrossRef]
- Fautin, D.G. Hexacorallians of the World. Available online: http://geoportal.kgs.ku.edu/hexacoral/anemone2/index.cfm (accessed on 7 February 2020).
- Sunagar, K.; Morgenstern, D.; Reitzel, A.M.; Moran, Y. Ecological venomics: How genomics, transcriptomics and proteomics can shed new light on the ecology and evolution of venom. J. Proteomics 2016, 135, 62–72. [Google Scholar] [CrossRef]
- O’Hara, E.P.; Caldwell, G.S.; Bythell, J. Equistatin and equinatoxin gene expression is influenced by environmental temperature in the sea anemone Actinia equina. Toxicon 2018, 153, 12–16. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Macrander, J.; Surm, J.M.; Aharoni, R.; Menard-Harvey, S.S.; Klock, A.; Leach, W.B.; Reitzel, A.M.; Moran, Y. Population Specific Adaptations in Venom Production to Abiotic Stressors in a Widely Distributed Cnidarian. bioRxiv 2020. [Google Scholar] [CrossRef]
- Jouiaei, M.; Sunagar, K.; Federman Gross, A.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an Ancient Venom: Recognition of a Novel Family of Cnidarian Toxins and the Common Evolutionary Origin of Sodium and Potassium Neurotoxins in Sea Anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef]
- Macrander, J.; Broe, M.; Daly, M. Tissue-specific venom composition and differential gene expression in sea anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-seq approach to identify putative toxins from acrorhagi in aggressive and non-aggressive Anthopleura elegantissima polyps. BMC Genomics 2015, 16. [Google Scholar] [CrossRef]
- Macrander, J.; Daly, M. Evolution of the Cytolytic Pore-Forming Proteins (Actinoporins) in Sea Anemones. Toxins 2016, 8, 368. [Google Scholar] [CrossRef]
- Fenner, P.J.; Williamson, J.A. Worldwide deaths and severe envenomation from jellyfish stings. Med. J. Aust. 1996, 165, 658. [Google Scholar] [CrossRef]
- Currie, B.J.; Jacups, S.P. Prospective study of Chironex fleckeri and other box jellyfish stings in the “Top End” of Australia’s Northern Territory. Med. J. Aust. 2005, 183, 631–636. [Google Scholar] [CrossRef]
- Winter, K.L.; Isbister, G.K.; McGowan, S.; Konstantakopoulos, N.; Seymour, J.E.; Hodgson, W.C. A pharmacological and biochemical examination of the geographical variation of Chironex fleckeri venom. Toxicol. Lett. 2010, 192, 419–424. [Google Scholar] [CrossRef]
- Yue, Y.; Yu, H.; Li, R.; Liu, S.; Xing, R.; Li, P. Insights into individual variations in nematocyst venoms from the giant jellyfish Nemopilema nomurai in the Yellow Sea. Sci. Rep. 2019, 9, 3361. [Google Scholar] [CrossRef]
- Kitahara, M.V.; Jaimes-Becerra, A.; Gamero-Mora, E.; Padilla, G.; Doonan, L.B.; Ward, M.; Marques, A.C.; Morandini, A.C.; Long, P.F. Reciprocal transplantation of the heterotrophic coral Tubastraea coccinea (Scleractinia: Dendrophylliidae) between distinct habitats did not alter its venom toxin composition. Ecol. Evol. 2020, 00, 1–10. [Google Scholar] [CrossRef]
- Kintner, A.H.; Seymour, J.E.; Edwards, S.L. Variation in lethality and effects of two Australian chirodropid jellyfish venoms in fish. Toxicon 2005, 46, 699–708. [Google Scholar] [CrossRef]
- Carrette, T.; Alderslade, P.; Seymour, J. Nematocyst ratio and prey in two Australian cubomedusans, Chironex fleckeri and Chiropsalmus sp. Toxicon 2002, 40, 1547–1551. [Google Scholar] [CrossRef]
- Damian-Serrano, A.; Haddock, S.H.D.; Dunn, C.W. The Evolution of Siphonophore Tentilla as Specialized Tools for Prey Capture. bioRxiv. 2019. [CrossRef]
- Carrette, T.; Straehler-Pohl, I.; Seymour, J. Early Life History of Alatina cfmoseri Populations from Australia and Hawaii with Implications for Taxonomy (Cubozoa: Carybdeida, Alatinidae). PLoS ONE 2014, 9, e84377. [Google Scholar] [CrossRef]
- Straehler-Pohl, I.; Jarms, G. Morphology and life cycle of Carybdea morandinii, sp. nov. (Cnidaria), a cubozoan with zooxanthellae and peculiar polyp anatomy. Zootaxa 2011, 2755, 36–56. [Google Scholar] [CrossRef]
- Klompen, A.; Sanders, S.; Cartwright, P. Influences of functional variation on venom expression in hydractiniid hydrozoans. Presented at the Gordon Research Conference: Venom Evolution, Function and Biomedical Applications, Mount Snow, VT, USA, 5–10 August 2018. [Google Scholar]
- Courtney, R.; Browning, S.; Seymour, J. Early Life History of the ‘Irukandji’ Jellyfish Carukia barnesi. PLoS ONE 2016, 11, e0151197. [Google Scholar] [CrossRef]
- Underwood, A.H.; Seymour, J.E. Venom ontogeny, diet and morphology in Carukia barnesi, a species of Australian box jellyfish that causes Irukandji syndrome. Toxicon 2007, 49, 1073–1082. [Google Scholar] [CrossRef]
- Courtney, R.; Sachlikidis, N.; Jones, R.; Seymour, J. Prey Capture Ecology of the Cubozoan Carukia barnesi. PLoS One 2015, 10, e0124256. [Google Scholar] [CrossRef]
- Gershwin, L.-A. Two new species of jellyfishes (Cnidaria: Cubozoa: Carybdeida) from tropical Western Australia, presumed to cause Irukandji Syndrome. Zootaxa 2005, 1084, 1–30. [Google Scholar] [CrossRef]
- Augusto-de-Oliveira, C.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.; Liberato, T.; Fukushima, I.; Serrano, S.M.; Zelanis, A. Dynamic Rearrangement in Snake Venom Gland Proteome: Insights into Bothrops jararaca Intraspecific Venom Variation. J. Proteome Res. 2016, 15, 3752–3762. [Google Scholar] [CrossRef]
- Goncalves-Machado, L.; Pla, D.; Sanz, L.; Jorge, R.J.B.; Leitao-De-Araujo, M.; Alves, M.L.M.; Alvares, D.J.; De Miranda, J.; Nowatzki, J.; de Morais-Zani, K.; et al. Combined venomics, venom gland transcriptomics, bioactivities, and antivenomics of two Bothrops jararaca populations from geographic isolated regions within the Brazilian Atlantic rainforest. J. Proteomics 2016, 135, 73–89. [Google Scholar] [CrossRef]
- Huang, H.W.; Liu, B.S.; Chien, K.Y.; Chiang, L.C.; Huang, S.Y.; Sung, W.C.; Wu, W.G. Cobra venom proteome and glycome determined from individual snakes of Naja atra reveal medically important dynamic range and systematic geographic variation. J. Proteomics 2015, 128, 92–104. [Google Scholar] [CrossRef]
- Massey, D.J.; Calvete, J.J.; Sanchez, E.E.; Sanz, L.; Richards, K.; Curtis, R.; Boesen, K. Venom variability and envenoming severity outcomes of the Crotalus scutulatus scutulatus (Mojave rattlesnake) from Southern Arizona. J. Proteomics 2012, 75, 2576–2587. [Google Scholar] [CrossRef]
- Dowell, N.L.; Giorgianni, M.W.; Kassner, V.A.; Selegue, J.E.; Sanchez, E.E.; Carroll, S.B. The Deep Origin and Recent Loss of Venom Toxin Genes in Rattlesnakes. Curr. Biol. 2016, 26, 2434–2445. [Google Scholar] [CrossRef]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet Breadth Mediates the Prey Specificity of Venom Potency in Snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef]
- Margres, M.J.; Wray, K.P.; Hassinger, A.T.; Ward, M.J.; McGivern, J.J.; Moriarty Lemmon, E.; Lemmon, A.R.; Rokyta, D.R. Quantity, not quality: rapid adaptation in a polygenic trait proceeded exclusively through expression differentiation. Mol. Biol. Evol. 2017, 34, 3099–3110. [Google Scholar] [CrossRef]
- Dunn, C. Siphonophores. Curr. Biol. 2009, 19, R233–R234. [Google Scholar] [CrossRef]
- Dunn, C.W.; Wagner, G.P. The evolution of colony-level development in the Siphonophora (Cnidaria:Hydrozoa). Dev. Genes Evol. 2006, 216, 743–754. [Google Scholar] [CrossRef]
- Ramos, V.; Vasconcelos, V. Palytoxin and analogs: biological and ecological effects. Mar. Drugs 2010, 8, 2021–2037. [Google Scholar] [CrossRef]
- Wu, C.H. Palytoxin: Membrane mechanisms of action. Toxicon 2009, 54, 1183–1189. [Google Scholar] [CrossRef]
- Mahnir, V.M.; Kozlovskaya, E.P.; Kalinovsky, A.I. Sea anemone Radianthus macrodactylus—A new source of palytoxin. Toxicon 1992, 30, 1449–1456. [Google Scholar] [CrossRef]
- Guppy, R.; Ackbarali, C.; Ibrahim, D. Toxicity of crude organic extracts from the zoanthid Palythoa caribaeorum: A biogeography approach. Toxicon 2019, 167, 117–122. [Google Scholar] [CrossRef]
- Klompen, A.; Sanders, S.; Cartwright, P. Differentially expressed venoms in functionally specialized polyps of the colonial hydrozoan Hydractinia symbiolongicarpus. Presented at the 7th European Evolution and Development Conference, Galway, Ireland, 265–29 June 2018. [Google Scholar]
- Sanders, S.M.; Shcheglovitova, M.; Cartwright, P. Differential gene expression between functionally specialized polyps of the colonial hydrozoan Hydractinia symbiolongicarpus (Phylum Cnidaria). BMC Genom. 2014, 15, 406. [Google Scholar] [CrossRef]
- McCloskey, B. Illustrated Glossary of Sea Anemone Anatomy. Available online: http://archive.li/L7PMk#selection-187.0–186.1 (accessed on 7 February 2020).
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef]
- Bocharova, E.; Kozevich, I. Modes of reproduction in sea anemones (Cnidaria, Anthozoa). Biol. Bull. 2011, 38, 849–860. [Google Scholar] [CrossRef]
- Wallace, C.C. Hexacorals 1: Sea Anemones and Anemone-like Animals (Actiniaria, Zoanthidea, Corallimorpharia, Ceriantharia and Antipatharia). In The Great Barrier Reef: Biology, Environment and Management; Hutchings, P., Kingsford, M., Hoegh-Guldberg, O., Eds.; CSIRO: Collingwood, VIC, Australia, 2008; pp. 198–207. [Google Scholar]
- Schlesinger, A.; Zlotkin, E.; Kramarsky-Winter, E.; Loya, Y. Cnidarian internal stinging mechanism. Proc. R. Soc. B 2009, 276, 1063–1067. [Google Scholar] [CrossRef]
- Han, J.; Zhang, X.; Komiya, T. Integrated Evolution of Cnidarians and Oceanic Geochemistry Before and During the Cambrian Explosion. In The Cnidaria, Past, Present and Future: The World of Medusa and Her Sisters; Goffredo, S., Dubinsky, Z., Eds.; Springer International Publishing: Cham, Switherland, 2016; 2p. [Google Scholar]
- Crowther, A. Character Evolution in Light of Phylogenetic Analysis And Taxonomic Revision of the Zooxanthellate Sea Anemone Families Thalassianthidae and Aliciidae. Ph.D. dissertation, University of Kansas, Lawrence, KS, USA,, 2013. [Google Scholar]
- Östman, C.; Kultima, J.R.; Roat, C.; Rundblom, K. Acontia and mesentery nematocysts of the sea anemone Metridium senile (Linnaeus, 1761) (Cnidaria: Anthozoa). Sci. Mar. 2010, 74, 483–497. [Google Scholar] [CrossRef]
- Lam, J.; Cheng, Y.W.; Chen, W.U.; Li, H.H.; Chen, C.S.; Peng, S.E. A detailed observation of the ejection and retraction of defense tissue acontia in sea anemone (Exaiptasia pallida). PeerJ 2017, 5, 1–11. [Google Scholar] [CrossRef]
- Purcell, J.E. Aggresive function and induced development of catch tentacles in the sea anemone Metridium senile (Coelenterata, Actiniaria). Biol. Bull. 1977, 153, 355–368. [Google Scholar] [CrossRef]
- Watson, G.M.; Mariscal, R.N. Comparative ultrastructure of catch tentacles and feeding tentacles in the sea anemone Haliplanella. Tissue Cell 1983, 15, 939–953. [Google Scholar] [CrossRef]
- Williams, R.B. Acrorhagi, catch tentacles and sweeper tentacles: a synopsis of aggression of actiniarian and scleractinian cnidaria. Hydrobiologia 1991, 216, 539–545. [Google Scholar] [CrossRef]
- Einat, D.L.; Nanette, E.C. Long-term effects of competition on coral growth and sweeper tentacle development. Mar. Ecol.: Prog. Ser. 2006, 313, 115–123. [Google Scholar] [CrossRef][Green Version]
- Sebens, K.P.; Miles, J.S. Sweeper Tentacles in a Gorgonian Octocoral: Morphological Modifications for Interference Competition. Biol. Bull. 1988, 175, 378–387. [Google Scholar] [CrossRef]
- Daly, M. The anatomy, terminology, and homology of acrorhagi and pseudoacrorhagi in sea anemones. Zool. Verh. 2003, 345, 89–102. [Google Scholar]
- Ramasamy, S.; Isbister, G.K.; Seymour, J.E.; Hodgson, W.C. Pharmacologically distinct cardiovascular effects of box jellyfish (Chironex fleckeri) venom and a tentacle-only extract in rats. Toxicol. Lett. 2005, 155, 219–226. [Google Scholar] [CrossRef]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle Transcriptome and Venom Proteome of the Pacific Sea Nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef]
- Ponce, D.; López-Vera, E.; Aguilar, M.B.; Sánchez-Rodríguez, J. Preliminary Results of the in Vivo and in Vitro Characterization of a Tentacle Venom Fraction from the Jellyfish Aurelia aurita. Toxins 2013, 5, 2420–2433. [Google Scholar] [CrossRef]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef]
- Bastos, C.L., Jr.; Varela, A.S., Jr.; Ferreira, S.P.; Nornberg, B.F.; Boyle, R.T. Who knows not where an anemone does wear his sting? Could polypeptides released from the columnar vesicles of Bunodosoma cangicum induce apoptosis in the ZF-L cell line? Toxicon 2016, 124, 73–82. [Google Scholar] [CrossRef]
- Moran, Y.; Genikhovich, G.; Gordon, D.; Wienkoop, S.; Zenkert, C.; Özbek, S.; Technau, U.; Gurevitz, M. Neurotoxin localization to ectodermal gland cells uncovers an alternative mechanism of venom delivery in sea anemones. Proc. R. Soc. B 2012, 279, 1351–1358. [Google Scholar] [CrossRef]
- Roveta, C.; Pica, D.; Puce, S. The cnidome of Olindias muelleri (Cnidaria: Hydrozoa: Limnomedusae) from South Adriatic Sea. Zoomorphology 2019, 138, 437–442. [Google Scholar] [CrossRef]
- Acuña, F.H.; Excoffon, A.C.; Ricci, L. Composition, biometry and statistical relationships between the cnidom and body size in the sea anemone Oulactis muscosa (Cnidaria: Actiniaria). J. Mar. Biol. Assoc. UK 2007, 87, 415–419. [Google Scholar] [CrossRef]
- Ardelean, A.; Fautin, D. Variability in nematocysts from a single individual of the sea anemone Actinodendron arboreum (Cnidaria: Anthozoa: Actiniaria). Hydrobiologia 2004, 530, 189–197. [Google Scholar] [CrossRef]
- Reft, A.J.; Daly, M. Morphology, distribution, and evolution of apical structure of nematocysts in hexacorallia. J. Morphol. 2012, 273, 121–136. [Google Scholar] [CrossRef]
- Avian, M.; Malej, A. Aurelia polyps and medusae (Scyphozoa; Semaeostomeae;Ulmaridae) in the Northern Adriatic: their cnidome and ecology. In PERSEUS International Workshop “Coming to Grips with the Jellyfish Phenomenon in the Southern European and other Seas: Research to The Rescue of Coastal Managers”; Prieto, L., Deidun, A., Malej, A., Shiganova, T., Tirelli, V., Eds.; Perseus: Cadiz, Spain, 2015; p. 17. [Google Scholar]
- McClounan, S.; Seymour, J. Venom and cnidome ontogeny of the cubomedusae Chironex fleckeri. Toxicon 2012, 60, 1335–1341. [Google Scholar] [CrossRef]
- Di Camillo, C.; Bo, M.; Puce, S.; Tazioli, S.; Bavestrello, G. The cnidome of Carybdea marsupialis (Cnidaria: Cubomedusae) from the Adriatic Sea. J. Mar. Biol. Assoc. UK 2006, 86, 705–709. [Google Scholar] [CrossRef]
- Peach, M.B.; Pitt, K.A. Morphology of the nematocysts of the medusae of two scyphozoans, Catostylus mosaicus and Phyllorhiza punctata (Rhizostomeae): implications for capture of prey. Invertebr. Biol. 2005, 124, 98–108. [Google Scholar] [CrossRef]
- Spier, D.; Stampar, S.N.; Prantoni, A.L. New record of the endangered cerianthid Ceriantheomorphe brasiliensis (Cnidaria: Hexacorallia) in Paranaguá Bay, southern Brazil. Mar. Biodivers. Rec. 2012, 5. [Google Scholar] [CrossRef]
- Strömberg, S.M.; Östman, C. The cnidome and internal morphology of Lophelia pertusa (Linnaeus, 1758) (Cnidaria, Anthozoa). Acta Zool. 2017, 98, 191–213. [Google Scholar] [CrossRef]
- Acuña, F.; Excoffon, A.; McKinstry, S.; Martínez, D. Characterization of Aulactinia (Actiniaria: Actiniidae) species from Mar del Plata (Argentina) using morphological and molecular data. Hydrobiologia 2007, 592, 249–256. [Google Scholar] [CrossRef]
- Hwang, J.S.; Ohyanagi, H.; Hayakawa, S.; Osato, N.; Nishimiya-Fujisawa, C.; Ikeo, K.; David, C.N.; Fujisawa, T.; Gojobori, T. The evolutionary emergence of cell type-specific genes inferred from the gene expression analysis of Hydra. Proc. Natl. Acad. Sci. USA 2007, 104, 14735. [Google Scholar] [CrossRef]
- Sebé-Pedrós, A.; Saudemont, B.; Chomsky, E.; Plessier, F.; Mailhé, M.-P.; Renno, J.; Loe-Mie, Y.; Lifshitz, A.; Mukamel, Z.; Schmutz, S.; et al. Cnidarian Cell Type Diversity and Regulation Revealed by Whole-Organism Single-Cell RNA-Seq. Cell 2018, 173, 1520–1534. [Google Scholar] [CrossRef]
- Darling, J.A.; Reitzel, A.R.; Burton, P.M.; Mazza, M.E.; Ryan, J.F.; Sullivan, J.C.; Finnerty, J.R. Rising starlet: the starlet sea anemone, Nematostella vectensis. BioEssays 2005, 27, 211–221. [Google Scholar] [CrossRef]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef]
- Sunagar, K.; Columbus-Shenkar, Y.Y.; Fridrich, A.; Gutkovich, N.; Aharoni, R.; Moran, Y. Cell type-specific expression profiling unravels the development and evolution of stinging cells in sea anemone. BMC Biol. 2018, 16, 108. [Google Scholar] [CrossRef]
- St Pierre, L.; Woods, R.; Earl, S.; Masci, P.P.; Lavin, M.F. Identification and analysis of venom gland-specific genes from the coastal taipan (Oxyuranus scutellatus) and related species. Cell Mol. Life Sci. 2005, 62, 2679–2693. [Google Scholar] [CrossRef]
- Rattmann, Y.D.; Pereira, C.R.; Cury, Y.; Gremski, W.; Marques, M.C.; da Silva-Santos, J.E. Vascular permeability and vasodilation induced by the Loxosceles intermedia venom in rats: involvement of mast cell degranulation, histamine and 5-HT receptors. Toxicon 2008, 51, 363–372. [Google Scholar] [CrossRef]
- Sade, Y.B.; Boia-Ferreira, M.; Gremski, L.H.; da Silveira, R.B.; Gremski, W.; Senff-Ribeiro, A.; Chaim, O.M.; Veiga, S.S. Molecular cloning, heterologous expression and functional characterization of a novel translationally-controlled tumor protein (TCTP) family member from Loxosceles intermedia (brown spider) venom. Int J. Biochem Cell Biol. 2012, 44, 170–177. [Google Scholar] [CrossRef]
- Surm, J.M.; Stewart, Z.K.; Papanicolaou, A.; Pavasovic, A.; Prentis, P.J. The draft genome of Actinia tenebrosa reveals insights into toxin evolution. Ecol. Evol. 2019, 9, 11314–11328. [Google Scholar] [CrossRef]
- Ohdera, A.; Ames, C.L.; Dikow, R.B.; Kayal, E.; Chiodin, M.; Busby, B.; La, S.; Pirro, S.; Collins, A.G.; Medina, M.; et al. Box, stalked, and upside-down? Draft genomes from diverse jellyfish (Cnidaria, Acraspeda) lineages: Alatina alata (Cubozoa), Calvadosia cruxmelitensis (Staurozoa), and Cassiopea xamachana (Scyphozoa). GigaScience 2019, 8. [Google Scholar] [CrossRef]
- Helmkampf, M.; Bellinger, M.R.; Geib, S.M.; Sim, S.B.; Takabayashi, M. Draft Genome of the Rice Coral Montipora capitata Obtained from Linked-Read Sequencing. Genome Biol. Evol. 2019, 11, 2045–2054. [Google Scholar] [CrossRef]
- Ying, H.; Hayward, D.C.; Cooke, I.; Wang, W.; Moya, A.; Siemering, K.R.; Sprungala, S.; Ball, E.E.; Foret, S.; Miller, D.J. The Whole-Genome Sequence of the Coral Acropora millepora. Genome Biol. Evol. 2019, 11, 1374–1379. [Google Scholar] [CrossRef]
- Wilding, C.S.; Fletcher, N.; Smith, E.K.; Prentis, P.; Weedall, G.D.; Stewart, Z. The genome of the sea anemone Actinia equina (L.): Meiotic toolkit genes and the question of sexual reproduction. Mar. Genomics 2020, 100753. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; MacRaild, C.A.; Sunanda, P.; Morales, R.A.; Peigneur, S.; Macrander, J.; Heidi, H.Y.; Daly, M.; Raghothama, S.; Dhawan, V.; et al. Structure, folding and stability of a minimal homologue from Anemonia sulcata of the sea anemone potassium channel blocker ShK. Peptides 2018, 99, 169–178. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Villegas-Moreno, J.; Mitchell, M.L.; Csoti, A.; Peigneur, S.; Amero, C.; Pennington, M.W.; Tytgat, J.; Panyi, G.; Norton, R.S. Synthesis, folding, structure and activity of a predicted peptide from the sea anemone Oulactis sp. with an ShKT fold. Toxicon 2018, 150, 50–59. [Google Scholar] [CrossRef]
- Lachumanan, R.; Armugam, A.; Durairaj, P.; Gopalakrishnakone, P.; Tan, C.H.; Jeyaseelan, K. In situ hybridization and immunohistochemical analysis of the expression of cardiotoxin and neurotoxin genes in Naja naja sputatrix. J. Histochem. Cytochem. 1999, 47, 551–560. [Google Scholar] [CrossRef]
- Ogawa, T.; Onoue, H.; Nakagawa, K.; Nomura, S.; Sueishi, K.; Hattori, S.; Kihara, H.; Ohno, M. Localization and expression of phospholipases A2 in Trimeresurus flavoviridis (habu snake) venom gland. Toxicon 1995, 33, 1645–1652. [Google Scholar] [CrossRef]
- Wolenski, F.S.; Layden, M.J.; Martindale, M.Q.; Gilmore, T.D.; Finnerty, J.R. Characterizing the spatiotemporal expression of RNAs and proteins in the starlet sea anemone, Nematostella vectensis. Nat. Protoc. 2013, 8, 900–915. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The birth and death of toxins with distinct functions: a case study in the sea anemone Nematostella. Mol. Biol. Evol. 2019, 36. [Google Scholar] [CrossRef]
- Wikramanayake, A.H.; Hong, M.; Lee, P.N.; Pang, K.; Byrum, C.A.; Bince, J.M.; Xu, R.; Martindale, M.Q. An ancient role for nuclear β-catenin in the evolution of axial polarity and germ layer segregation. Nature 2003, 426, 446–450. [Google Scholar] [CrossRef]
- Richter, S.; Helm, C.; Meunier, F.A.; Hering, L.; Campbell, L.I.; Drukewitz, S.H.; Undheim, E.A.; Jenner, R.A.; Schiavo, G.; Bleidorn, C. Comparative analyses of glycerotoxin expression unveil a novel structural organization of the bloodworm venom system. BMC Evol. Biol. 2017, 17, 64. [Google Scholar] [CrossRef]
- Basulto, A.; Pérez, V.M.; Noa, Y.; Varela, C.; Otero, A.J.; Pico, M. Immunohistochemical targeting of sea anemone cytolysins on tentacles, mesenteric filaments and isolated nematocysts of Stichodactyla helianthus. J. Exp. Zool., Part. A 2006, 305, 253–258. [Google Scholar] [CrossRef]
- Horgan, R.P.; Kenny, L.C. ‘Omic’ technologies: genomics, transcriptomics, proteomics and metabolomics. Obstet. Gynaecol. 2011, 13, 189–195. [Google Scholar] [CrossRef]
- Chappell, L.; Russell, A.J.C.; Voet, T. Single-Cell (Multi)omics Technologies. Annu. Rev. Genomics Hum. Genet. 2018, 19, 15–41. [Google Scholar] [CrossRef]
- Angel, P.M.; Caprioli, R.M. Matrix-assisted laser desorption ionization imaging mass spectrometry: in situ molecular mapping. Biochemistry 2013, 52, 3818–3828. [Google Scholar] [CrossRef]
- Chen, X.; Leahy, D.; Van Haeften, J.; Hartfield, P.; Prentis, P.J.; van der Burg, C.A.; Surm, J.M.; Pavasovic, A.; Madio, B.; Hamilton, B.R. A versatile and robust serine protease inhibitor scaffold from Actinia tenebrosa. Mar. Drugs 2019, 17, 701. [Google Scholar] [CrossRef]
- Mitchell, M.L.; Hamilton, B.R.; Madio, B.; Morales, R.A.V.; Tonkin-Hill, G.Q.; Papenfuss, A.T.; Purcell, A.W.; King, G.F.; Undheim, E.A.B.; Norton, R.S. The use of imaging mass spectrometry to study peptide toxin distribution in Australian sea anemones. Aust. J. Chem. 2017, 70, 1235–1237. [Google Scholar] [CrossRef]
- Undheim, E.A.; Sunagar, K.; Hamilton, B.R.; Jones, A.; Venter, D.J.; Fry, B.G.; King, G.F. Multifunctional warheads: Diversification of the toxin arsenal of centipedes via novel multidomain transcripts. J. Proteomics 2014, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.; Hamilton, B.R.; Kurniawan, N.D.; Bowlay, G.; Cribb, B.W.; Merritt, D.J.; Fry, B.G.; King, G.F.; Venter, D.J. Production and packaging of a biological arsenal: Evolution of centipede venoms under morphological constraint. Proc. Natl. Acad. Sci. USA 2015, 112, 4026–4031. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.R.; Marshall, D.L.; Casewell, N.R.; Harrison, R.A.; Blanksby, S.J.; Undheim, E.A.B. Mapping enzyme activity on tissue by functional-mass spectrometry imaging. Angew. Chem. Int. Edit. 2019. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashwood, L.M.; Norton, R.S.; Undheim, E.A.B.; Hurwood, D.A.; Prentis, P.J. Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Mar. Drugs 2020, 18, 202. https://doi.org/10.3390/md18040202
Ashwood LM, Norton RS, Undheim EAB, Hurwood DA, Prentis PJ. Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Marine Drugs. 2020; 18(4):202. https://doi.org/10.3390/md18040202
Chicago/Turabian StyleAshwood, Lauren M., Raymond S. Norton, Eivind A. B. Undheim, David A. Hurwood, and Peter J. Prentis. 2020. "Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context" Marine Drugs 18, no. 4: 202. https://doi.org/10.3390/md18040202
APA StyleAshwood, L. M., Norton, R. S., Undheim, E. A. B., Hurwood, D. A., & Prentis, P. J. (2020). Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Marine Drugs, 18(4), 202. https://doi.org/10.3390/md18040202