Whole Genome Sequence of Dermacoccus abyssi MT1.1 Isolated from the Challenger Deep of the Mariana Trench Reveals Phenazine Biosynthesis Locus and Environmental Adaptation Factors

 ,
,  , , , ,
, , , ,  , and
, and 
Abstract
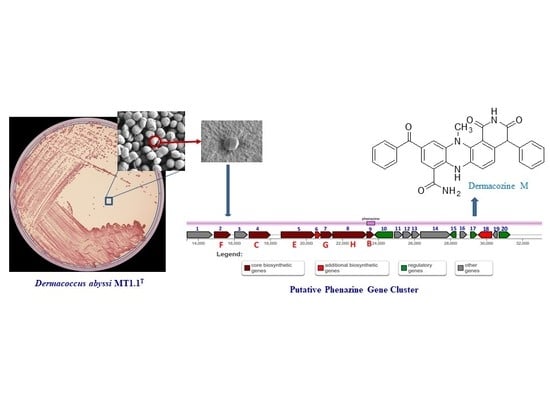
1. Introduction
2. Results and Discussion
2.1. Compound Identification
2.2. Genome Sequencing and Annotation
2.3. Secondary Metabolite-Biosynthetic Gene Clusters
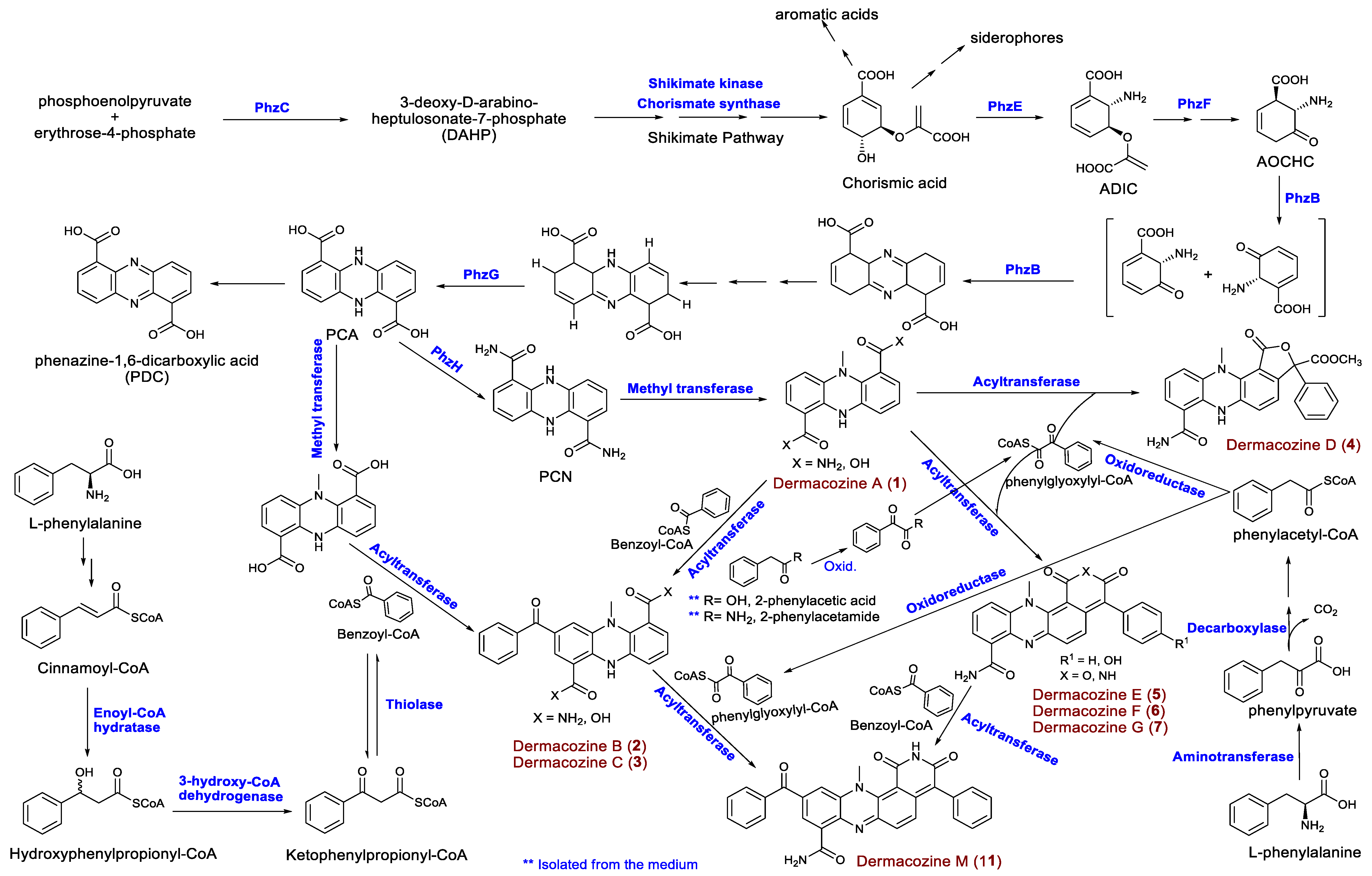
Phenazine Biosynthetic Gene Clusters
2.4. Genomic Insights Into Adaptation Strategies
2.4.1. Cold Shock Response
2.4.2. Osmotic Stress Response
2.4.3. Oxidative Stress Response
2.4.4. Respiration
2.4.5. Cell Wall/Membrane Alteration
2.4.6. Carbon Starvation and Storage
2.4.7. Remineralization of Organic Matter
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Microorganism
3.3. Fermentation Conditions
3.4. Purification of Dermacozine M
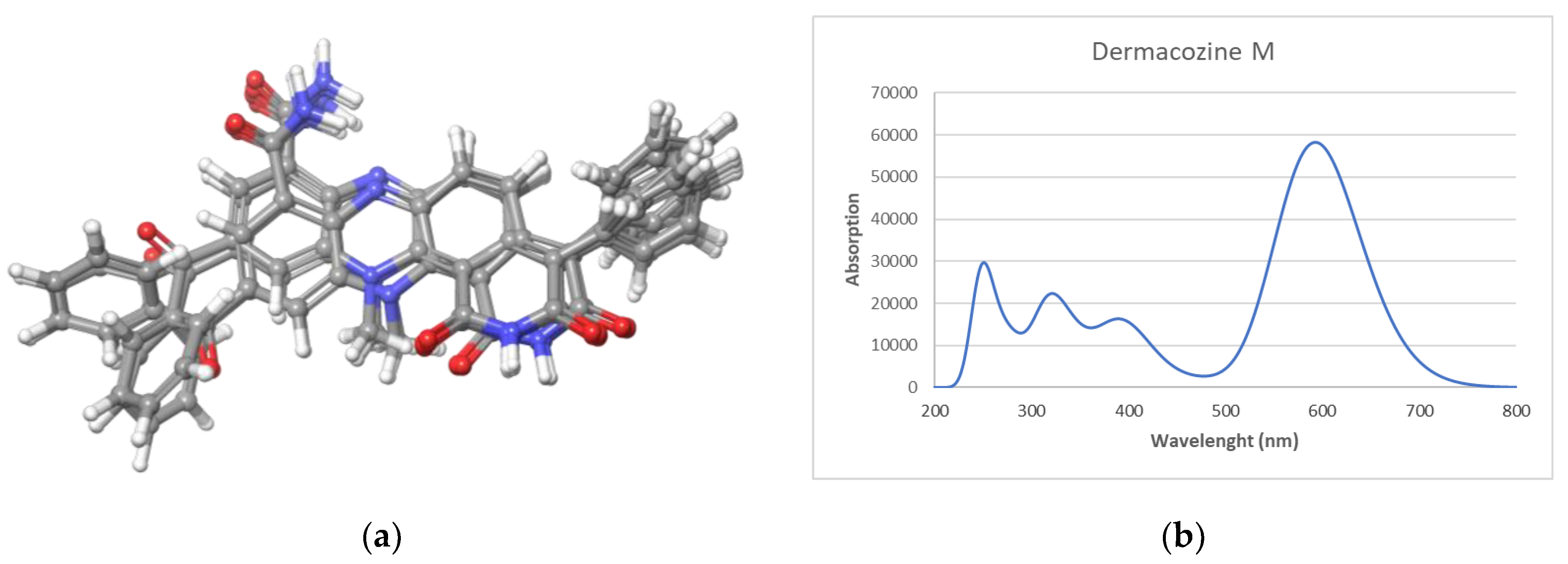
3.5. Computational Calculations
3.6. Genome Sequencing Information
3.6.1. DNA Extraction and Genome Sequencing
3.6.2. Detection of the Gene Clusters
3.6.3. Genbank Accession Number
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Li, S.J.; Li, T.J.; Liang, Z.Z.; Zhao, C.Q. Novel natural products from extremophilic fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Hassan, H.A.; Alhadrani, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme environments: microbiology leading to specialized metabolites. J. Appl. Microbiol. 2020, 128, 630–657. [Google Scholar] [CrossRef] [PubMed]
- Kamjam, M.; Sivalingam, P.; Deng, Z.; Hong, K. Deep sea actinomycetes and their secondary metabolites. Front. Microbiol. 2017, 8, 760. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T.; Goodfellow, M. Dark, rare and inspirational microbial matter in the extremobiosphere: 16 000 m of bioprospecting campaigns. Microbiology 2019, 165, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Pathom-Aree, W.; Stach, J.E.; Ward, A.C.; Horikoshi, K.; Bull, A.T.; Goodfellow, M. Divesity of actinomycetes isolated from Challenger Deep sediment (10,898 m) from the Mariana Trench. Extremophiles 2006, 10, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mageed, W.M.; Milne, B.F.; Wagner, M.; Schumacher, M.; Sandor, P.; Pathom-aree, W.; Goodfellow, M.; Bull, A.T.; Horikoshi, K.; Ebel, R.; et al. Dermacozines, a new phenazine family from deep-sea dermacocci isolated from a Mariana Trench sediment. Org. Biomol. Chem. 2010, 8, 2352–2362. [Google Scholar] [CrossRef] [PubMed]
- Ghanta, V.R.; Pasula, A.; Soma, L.; Raman, B. Synthetic studies on dermanozines: First synthesis of dermacozines A, B and C. ChemistrySelect 2016, 6, 1296–1299. [Google Scholar] [CrossRef]
- Pathom-aree, W.; Nogi, Y.; Sutcliffe, I.C.; Ward, A.C.; Horikoshi, K.; Bull, A.T.; Goodfellow, M. Dermacoccus abyssi sp. nov., a piezotolerant actinomycete isolated from the Mariana Trench. Int. J. Syst. Evol. Microbiol. 2006, 56, 1233–1237. [Google Scholar] [CrossRef]
- Undabarrena, A.; Ugalde, J.A.; Seeger, M.; Camara, R. Genomic data mining of the marine actinobacteria Streptomyces sp. H-KF8 unveils insights into multi-stress related genes and metabolic pathways involved in antimicrobial synthesis. PeerJ 2017, 5, e2912. [Google Scholar] [CrossRef]
- Pierson, L.S., 3rd; Pierson, E.A. Metabolism and function of phenazines in bacteria: Impacts on the behavior of bacteria in the environment and biotechnological processes. Appl. Microbiol. Biotechnol. 2010, 86, 1659–1670. [Google Scholar] [CrossRef]
- Guttenberger, N.; Blankenfeldt, W.; Breinbauer, R. Recent developments in the isolation, biological function, biosynthesis, and synthesis of phenazine natural products. Bioorg. Med. Chem. 2017, 25, 6149–6166. [Google Scholar] [CrossRef] [PubMed]
- Laursen, J.B.; Nielsen, J. Phenazine natural products: biosynthesis, synthetic analogues, and biological activity. Chem. Rev. 2004, 104, 1663–1686. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Abdel-Mageed, W.M.; Ebel, R.; Bull, A.T.; Goodfellow, M.; Fiedler, H.-P.; Jaspars, M. Dermacozines H–J Isolated from a deep-sea strain of Dermacoccus abyssi from Mariana Trench sediments. J. Nat. Prod. 2014, 77, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Suite, 5th ed.; 2018-4; Schrödinger, LCC: New York, NY, USA, 2018.
- Jacquemin, D.; Laurent, A.D.; Perpète, E.A.; André, J.-M. An Ab initio simulation of the UV/visible spectra of N-benzylideneaniline dyes. Int. J. Quantum Chem. 2009, 9, 3506–3515. [Google Scholar] [CrossRef]
- Bruhn, T.; Schaumlöffel, A.; Hemberger, Y.; Bringmann, G. SpecDis: Quantifying the comparison of calculated and experimental electronic circular dichroism spectra. Chirality 2013, 25, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Angiuoli, S.V.; Gussman, A.; Klimke, W.; Cochrane, G.; Field, D.; Garrity, G.; Kodira, C.D.; Kyrpides, N.; Madupu, R.; Markowitz, V.; et al. Toward an online repository of Standard Operating Procedures (SOPs) for (meta) genomic annotation. OMICS 2008, 12, 137–141. [Google Scholar] [CrossRef]
- Tatusova, T.; Ciufo, S.; Fedorov, B.; O’Neill, K.; Tolstoy, I. RefSeq microbial genomes database: New representation and annotation strategy. Nucleic Acids Res. 2014, 42, D553–D559. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Rui, Z.; Ye, M.; Wang, S.; Fujikawa, K.; Akerele, B.; Aung, M.; Floss, H.G.; Zhang, W.; Yu, T.W. Insights into a divergent phenazine biosynthetic pathway governed by a plasmid-born esmeraldin gene cluster. Chem. Biol. 2012, 19, 1116–1125. [Google Scholar] [CrossRef][Green Version]
- Parsons, J.F.; Greenhagen, B.T.; Shi, K.; Calabrese, K.; Robinson, H.; Ladner, J.E. Structural and functional analysis of the pyocyanin biosynthetic protein PhzM from Pseudomonas aeruginosa. Biochemistry 2007, 46, 1821–1828. [Google Scholar] [CrossRef]
- Coates, R.C.; Bowen, B.P.; Oberortner, E.; Thomashow, L.; Hadjithomas, M.; Zhao, Z.; Ke, J.; Silva, L.; Louie, K.; Wang, G.; et al. An integrated workflow for phenazine-modifying enzyme characterization. J. Ind. Microbiol. Biotechnol. 2018, 45, 567–577. [Google Scholar] [CrossRef]
- Fujii, S.; Nakasone, K.; Horikoshi, K. Cloning of two cold shock genes, cspA and cspG, from the deep-sea psychrophilic bacterium Shewanella violacea strain DSS12. FEMS Microbiol. Lett. 1999, 178, 123–128. [Google Scholar] [CrossRef]
- Li, S.; Xiao, X.; Sun, P.; Wang, F. Screening of genes regulated by cold shock in Shewanella piezotolerans WP3 and time course expression of cold-regulated genes. Arch. Microbiol. 2008, 189, 549–556. [Google Scholar] [CrossRef]
- Gumley, A.W.; Inniss, W.E. Cold shock proteins and cold acclimation proteins in the psychrotrophic bacterium Pseudomonas putida Q5 and its transconjugant. Can. J. Microbiol. 1996, 42, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Redder, P.; Hausmann, S.; Khemici, V.; Yasrebi, H.; Linder, P. Bacterial versatility requires DEAD-box RNA helicases. FEMS Microbiol. Rev. 2015, 39, 392–412. [Google Scholar] [CrossRef] [PubMed]
- Johansson, I.; Karlsson, M.; Johanson, U.; Larsson, C.; Kjellbom, P. The role of aquaporins in cellular and whole plant water balance. Biochim. Biophys. Acta 2000, 1465, 324–342. [Google Scholar] [CrossRef]
- Bartlett, D.H. Microbial adaptations to the psychrosphere/piezosphere. J. Mol. Microbiol. Biotechnol. 1999, 1, 93–100. [Google Scholar]
- Campanaro, S.; Treu, L.; Valle, G. Protein evolution in deep sea bacteria: An analysis of amino acids substitution rates. BMC Evol. Biol. 2008, 8, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Goordial, J.; Raymond-Bouchard, I.; Zolotarov, Y.; de Bethencourt, L.; Ronholm, J.; Shapiro, N.; Woyke, T.; Stromvik, M.; Greer, C.; Bakermans, C.; et al. Cold adaptive traits revealed by comparative genomic analysis of the eurypsychrophile Rhodococcus sp. JG3 isolated from high elevation McMurdo Dry Valley permafrost, Antarctica. FEMS Microbiol. Ecol. 2016, 92, fiv154. [Google Scholar] [PubMed]
- Yano, Y.; Nakayama, A.; Ishihara, K.; Saito, H. Adaptive changes in membrane lipids of barophilic bacteria in response to changes in growth pressure. Appl. Environ. Microbiol. 1998, 64, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Unden, G.; Bongaerts, J. Alternative respiratory pathways of Escherichia coli: Energetics and transcriptional regulation in response to electron acceptors. Biochim. Biophys. Acta 1997, 1320, 217–234. [Google Scholar] [CrossRef]
- Nichols, D.S.; Nichols, P.D.; Russell, N.J.; Davies, N.W.; McMeekin, T.A. Polyunsaturated fatty acids in the psychrophilic bacterium Shewanella gelidimarina ACAM 456T: Molecular species analysis of major phospholipids and biosynthesis of eicosapentaenoic acid. Biochim. Biophys. Acta 1997, 1347, 164–176. [Google Scholar] [CrossRef]
- Usui, K.; Hiraki, T.; Kawamoto, J.; Kurihara, T.; Nogi, Y.; Kato, C.; Abe, F. Eicosapentaenoic acid plays a role in stabilizing dynamic membrane structure in the deep-sea piezophile Shewanella violacea: a study employing high-pressure time-resolved fluorescence anisotropy measurement. Biochim. Biophys. Acta 2012, 1818, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Gorshkova, N.M.; Bowman, J.P.; Lysenko, A.M.; Zhukova, N.V.; Sergeev, A.F.; Mikhailov, V.V.; Nicolau, D.V. Shewanella pacifica sp. nov., a polyunsaturated fatty acid-producing bacterium isolated from sea water. Int. J. Syst. Evol. Microbiol. 2004, 54, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Shang, Y.; Huang, W.; Wang, C. Glycerol-3-phosphate acyltransferase contributes to triacylglycerol biosynthesis, lipid droplet formation, and host invasion in Metarhizium robertsii. Appl. Environ. Microbiol. 2013, 79, 7646–7653. [Google Scholar] [CrossRef]
- Cannon, G.C.; Heinhorst, S.; Kerfeld, C.A. Carboxysomal carbonic anhydrases: Structure and role in microbial CO2 fixation. Biochim. Biophys. Acta 2010, 1804, 382–392. [Google Scholar] [CrossRef]
- Piontek, J.; Händel, N.; De Bodt, C.; Harlay, J.; Chou, L.; Engel, A. The utilization of polysaccharides by heterotrophic bacterioplankton in the Bay of Biscay (North Atlantic Ocean). J. Plankton Res. 2011, 33, 1719–1735. [Google Scholar] [CrossRef]
- Aono, E.; Baba, T.; Ara, T.; Nishi, T.; Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Hasegawa, M.; Takai, Y.; Okumura, Y.; et al. Complete genome sequence and comparative analysis of Shewanella violacea, a psychrophilic and piezophilic bacterium from deep sea floor sediments. Mol. Biosyst. 2010, 6, 1216–1226. [Google Scholar] [CrossRef]
- Kupchan, S.M.; Briton, R.W.; Ziegler, M.F.; Siegel, C.W. Bruceantin, a new potent antileukemic simaroubolide from Brucea antidysenterica. J. Org. Chem. 1973, 38, 178–179. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology; John Wiley and Sons, Inc.: New York, NY, USA, 1994. [Google Scholar]

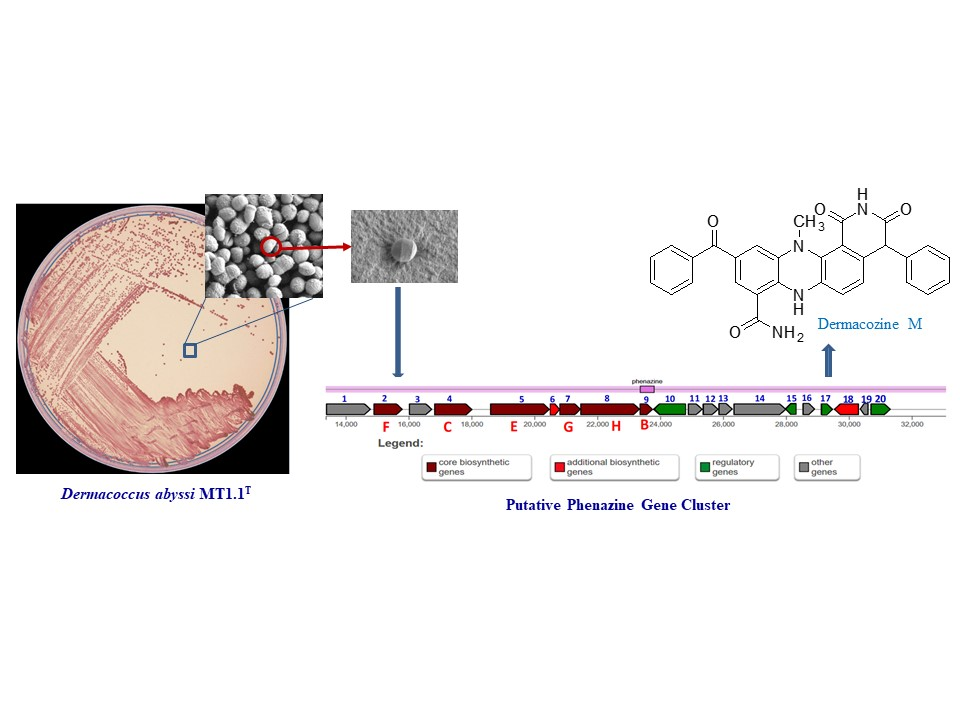
 ), NOESY (
), NOESY ( ), and HMBC (H
), and HMBC (H C) correlations.
C) correlations.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| no. | 11 | |
|---|---|---|
| δC, mult. | δH, mult (J in Hz) | |
| 1 | 131.0, C | - |
| 2 | 127.2, CH | 8.21, d (1.8) |
| 3 | 134.9†, C | - |
| 4 | 119.8, CH | 8.13, d (1.8) |
| 4a | 134.1, C | - |
| 5a | 139.2, C | - |
| 6 | 100.8, C | - |
| 7 | 139.4, C | - |
| 8 | 130.1, CH | 7.25, d (9.7) |
| 9 | 134.8, CH | 7.35, d (9.7) |
| 9a | 152.3, C | - |
| 10a | 137.4, C | - |
| 11 | 165.5, C | - |
| 12 | a. 7.99, brs b. 8.93, brs | |
| 13 | 161.9 †, C | - |
| 14 | 11.34, brs | |
| 15 | 161.7, C | - |
| 16 | 123.4, C | - |
| 17 | 133.9, C | - |
| 18 | 131.1, CH | 7.33, dd (7.5, 1.3) |
| 19 | 127.9, CH | 7.49, t (7.5) |
| 20 | 127.7, CH | 7.44, td (7.5, 1.3) |
| 21 | 127.9, CH | 7.49, t (7.5) |
| 22 | 131.1, CH | 7.33, dd (7.5, 1.3) |
| 23 | 45.6, CH3 | 3.69, s |
| 24 | 194.4, C | - |
| 25 | 136.5, C | - |
| 26 | 129.6, CH | 7.85, dd (7.5, 1.5) |
| 27 | 128.6, CH | 7.64, td (7.5, 1.5) |
| 28 | 133.2, CH | 7.76, m |
| 29 | 128.6, CH | 7.64, td (7.5, 1.5) |
| 30 | 129.6, CH | 7.85, dd (7.5, 1.5) |
| Features | D. abyssi (MT1.1) |
|---|---|
| Assembly size, bp | 3,160,906 |
| No. of contigs | 55 |
| G + C | 68.1 |
| Fold coverage | 122.46× |
| Percentage of bases | 99.7% |
| N50 | 159983 |
| L50 | 7 |
| Genes | 3,335 |
| CDs | 2,959 |
| Pseudogenes | 163 |
| Pseudogenes (frameshifted) | 82 |
| Protein encoding genes | 2810 |
| rRNA | 6 |
| tRNA | 49 |
| ncRNAs | 3 |
| Accession no. | NZ_QWLM00000000 |
| No. | Locus Tag | Nucleotide (nt) | Protein | Accession Number | Function | Homologue | Accession Number | Identity | Coverage Percentage | E Value |
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | D1832_RS13610 | 846 | PhzF family phenazine biosynthesis isomerase | WP_118914810 | DHHA isomerase | PhzF family phenazine biosynthesis isomerase [Streptomyces purpureus] | WP_019884027 | 76.95% | 99% | 6e-150 |
| 3 | D1832_RS15170 | 669 | Hypothetical protein | WP_147362718 | Unknown activity | Hypothetical protein D1832_13610 [Dermacoccus abyssi] | RHW43994 | 99.28% | 62% | 1e-98 |
| 4 | D1832_RS13620 | 1125 | 3-deoxy-7-phosphoheptulonate synthase (PhzC) | WP_118914924 | DAHP synthase | 3-deoxy-7-phosphoheptulonate synthase [Streptomyces sp. uw30] | WP_147995366 | 67.65% | 99% | 6e-176 |
| 5 | D1832_RS13625 | 1896 | Phenazine-specific anthranilate synthase (PhzE) | WP_118914814 | ADIC synthase | Phenazine-specific anthranilate synthase component I [Streptomyces purpureus] | WP_019884038 | 69.54% | 98% | 0.0 |
| 6 | D1832_RS13630 | 306 | Hypothetical protein (biosynthetic-additional thio_amide) | WP_118914815 | Oxidoreductase | YdhR family protein [Streptomyces sp. uw30] | WP_147993498 | 62.11% | 93% | 9e-33 |
| 7 | D1832_RS13635 | 621 | Pyridoxamine 5’-phosphate oxidase (PhzG) | WP_118914817 | Phenazine-1,6-dicarboxylic acid (PDC) formation | Pyridoxal 5’-phosphate synthase [Streptomyces sp. uw30] | WP_147993497 | 60.68% | 100% | 3e-78 |
| 8 | D1832_RS13640 | 1842 | Asparagine synthase (PhzH) | WP_118914818 | Asparagine synthase (glutamine-hydrolyzing) activity | Asparagine synthase (Glutamine-hydrolyzing) [Streptomyces thermocarboxydus] | WP_137209574 | 79.13% | 99% | 0.0 |
| 9 | D1832_RS13645 | 444 | Phenazine biosynthesis protein: phzB | WP_118914868 | 6-amino-5-oxocyclohex-2-ene-1-carboxylic acid (AOCHC) dimerization | phenazine biosynthesis protein [Streptomyces thermocarboxydus] | WP_137209661 | 83.67% | 100% | 3e-91 |
| 10 | D1832_RS13650 | 993 | WYL domain-containing protein | WP_118914820 | Ligand-binding regulatory domain | WYL domain-containing protein [Streptomyces glaucescens] | WP_043504928 | 69.78% | 97% | 2e-145 |
| 11 | D1832_RS13655 | 402 | RidA family protein | WP_118914821 | Enamine/imine deaminase activity | RidA family protein [Streptomyces exfoliatus] | WP_037644077 | 80.30% | 99% | 2e-71 |
| 12 | D1832_RS13660 | 492 | DUF488 domain-containing protein | WP_118914870 | Unknown | DUF488 domain-containing protein [Streptosporangiaceae bacterium YIM 75507] | WP_119930667 | 68.12% | 98% | 5e-70 |
| 13 | D1832_RS13665 | 393 | DUF4186 domain-containing protein | WP_118914823 | Unknown | DUF4186 domain-containing protein [Nocardia suismassiliense] | WP_107655452 | 81.15% | 93% | 2e-66 |
| 14 | D1832_RS13670 | 1617 | Hypothetical protein | WP_118914825 | Asparaginase activity | Asparaginase [Blastococcus sp. DSM 44205] | SDE71699 | 61.57% | 100% | 0.0 |
| 15 | D1832_RS13675 | 344 | TraR/DksA family transcriptional regulator | WP_118913125 | Transcriptional regulation | DNA-binding protein [Micropruina glycogenica] | WP_105185941 | 62.92% | 77% | 2e-24 |
| 16 | D1832_RS13680 | 360 | VOC family protein | WP_118912933 | Dioxygenase and glyoxalase | VOC family protein [Propionibacterium freudenreichii] | WP_013161949 | 92.50% | 100% | 2e-76 |
| 17 | D1832_RS13685 | 309 | TetR/AcrR family transcriptional regulator | WP_118914637 | Transcriptional regulation | TetR/AcrR family transcriptional regulator [Streptomyces mirabilis] | WP_075033331 | 73.53% | 99% | 1e-41 |
| 18 | D1832_RS13690 | 702 | Alpha/beta hydrolase | WP_118914826 | Carboxylic ester hydrolase activity (esterase) | Alpha/beta hydrolase [Streptacidiphilus jeojiense] | WP_030267495 | 47.84% | 98% | 2e-64 |
| 19 | D1832_RS13695 | 195 | Hypothetical protein | WP_147362719 | Unknown | Hypothetical protein D1832_13690 [Dermacoccus abyssi] | RHW44004 | 98.44% | 100% | 2e-36 |
| 20 | D1832_RS13700 | 579 | TetR/AcrR family transcriptional regulator | WP_118914830 | Transcriptional regulator | TetR family transcriptional regulator [Actinotalea sp. HO-Ch2] | WP_149205434 | 58.19% | 91% | 1e-54 |
| Protein | AA | Accession Number | Function |
|---|---|---|---|
| Shikimate kinase | 213 | WP_118912523 | Shikimic acid biosynthesis |
| Dehydroquinate synthase | 354 | WP_118912522 | Shikimic acid biosynthesis |
| Chorismate synthase | 407 | WP_118912574 | Shikimic acid biosynthesis |
| Enoyl-CoA hydratase | 265 | WP_118912167 | β-hydroxyphenyl propionyl-CoA biosynthesis |
| 3-hydroxyacyl-CoA dehydrogenase | 704 | WP_118913237 | β-ketophenylpropionyl-CoA biosynthesis |
| Thiolase family protein | 405 | WP_047311533 | Benzoyl-CoA biosynthesis |
| Pyridoxal phosphate-dependent aminotransferase | 405 | WP_047311611 | Transamination |
| Acyltransferase family protein | 667 | WP_118912475 | Acyl condensation |
| Class I SAM-dependent DNA methyltransferase | 912 | WP_118914833 | Methylation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Mageed, W.M.; Juhasz, B.; Lehri, B.; Alqahtani, A.S.; Nouioui, I.; Pech-Puch, D.; Tabudravu, J.N.; Goodfellow, M.; Rodríguez, J.; Jaspars, M.; et al. Whole Genome Sequence of Dermacoccus abyssi MT1.1 Isolated from the Challenger Deep of the Mariana Trench Reveals Phenazine Biosynthesis Locus and Environmental Adaptation Factors. Mar. Drugs 2020, 18, 131. https://doi.org/10.3390/md18030131
Abdel-Mageed WM, Juhasz B, Lehri B, Alqahtani AS, Nouioui I, Pech-Puch D, Tabudravu JN, Goodfellow M, Rodríguez J, Jaspars M, et al. Whole Genome Sequence of Dermacoccus abyssi MT1.1 Isolated from the Challenger Deep of the Mariana Trench Reveals Phenazine Biosynthesis Locus and Environmental Adaptation Factors. Marine Drugs. 2020; 18(3):131. https://doi.org/10.3390/md18030131
Chicago/Turabian StyleAbdel-Mageed, Wael M., Bertalan Juhasz, Burhan Lehri, Ali S. Alqahtani, Imen Nouioui, Dawrin Pech-Puch, Jioji N. Tabudravu, Michael Goodfellow, Jaime Rodríguez, Marcel Jaspars, and et al. 2020. "Whole Genome Sequence of Dermacoccus abyssi MT1.1 Isolated from the Challenger Deep of the Mariana Trench Reveals Phenazine Biosynthesis Locus and Environmental Adaptation Factors" Marine Drugs 18, no. 3: 131. https://doi.org/10.3390/md18030131
APA StyleAbdel-Mageed, W. M., Juhasz, B., Lehri, B., Alqahtani, A. S., Nouioui, I., Pech-Puch, D., Tabudravu, J. N., Goodfellow, M., Rodríguez, J., Jaspars, M., & Karlyshev, A. V. (2020). Whole Genome Sequence of Dermacoccus abyssi MT1.1 Isolated from the Challenger Deep of the Mariana Trench Reveals Phenazine Biosynthesis Locus and Environmental Adaptation Factors. Marine Drugs, 18(3), 131. https://doi.org/10.3390/md18030131