Bioactive Metabolites of Marine Origin Have Unusual Effects on Model Membrane Systems

 , , , , , and
, , , , , and
Abstract

1. Introduction
2. Results
3. Discussion
4. Material and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chellat, M.F.; Raguž, L.; Riedl, R. Targeting Antibiotic Resistance. Angew. Chem.-Int. Ed. 2016, 55, 6600–6626. [Google Scholar] [CrossRef]
- Zahreddine, H.; Borden, K.L.B. Mechanisms and insights into drug resistance in cancer. Front. Pharmacol. 2013, 4, 28. [Google Scholar] [CrossRef]
- Vila, J.; Moreno-Morales, J. Ballesté-Delpierre, C. Current landscape in the discovery of novel antibacterial agents. Clin. Microbiol. Infect. 2019. [Google Scholar] [CrossRef]
- Kållberg, C.; Salvesen Blix, H.; Laxminarayan, R. Challenges in Antibiotic R&D Calling for a Global Strategy Considering Both Short- And Long-Term Solutions. ACS Infect. Dis. 2019, 5, 1265–1268. [Google Scholar] [CrossRef]
- Van Blitterswijk, W.J.; Verheij, M. Anticancer mechanisms and clinical application of alkylphospholipids. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2013, 1831, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2009, 26, 170–244. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Ercolano, G.; De Cicco, P.; Ianaro, A. New drugs from the sea: Pro-apoptotic activity of sponges and algae derived compounds. Mar. Drugs 2019, 17, 31. [Google Scholar] [CrossRef]
- Shinde, P.; Banerjee, A.; Mandhare, P. Marine natural products as source of new drugs: A patent review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef]
- Gundersen, L.-L. Synthesis and biological activities of marine terpene-adenine hybrids and synthetic analogs. Phytochem. Rev. 2013, 12, 467–486. [Google Scholar] [CrossRef]
- Sun, Z.H.; Cai, Y.H.; Fan, C.Q.; Tang, G.H.; Bin Luo, H.; Yin, S. Six new tetraprenylated alkaloids from the South China Sea gorgonian Echinogorgia pseudossapo. Mar. Drugs 2014, 12, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, J.; Chen, M.; Wang, K.; Tenney, H.; Crews, P. Bioactive secondary metabolites from the marine sponge genus Agelas. Mar. Drugs 2017, 15, 351. [Google Scholar] [CrossRef] [PubMed]
- García, P.A.; Valles, D.; Díez, E.; Castro, M.Á. Marine Alkylpurines: A promising group of bioactive marine natural products. Mar. Drugs 2018, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Vik, A.; Proszenyák, Á.; Vermeersch, M.; Cos, P.; Maes, L.; Gundersen, L.L. Screening of agelasine D and analogs for inhibitory activity against pathogenic protozoa; identification of hits for visceral leishmaniasis and Chagas disease. Molecules 2009, 14, 279–288. [Google Scholar] [CrossRef]
- Vik, A.; Hedner, C.; Charnock, E.; Tangen, L.W.; Samuelsen, R.; Larsson, Ø.; Bohlin, L.; Gundersen, L.L. Antimicrobial and cytotoxic activity of agelasine and agelasimine analogs. Bioorganic Med. Chem. 2007, 15, 4016–4037. [Google Scholar] [CrossRef]
- Vik, A.; Hedner, C.; Charnock, E.; Samuelsen, R.; Larsson, Ø.; Gundersen, L.L.; Bohlin, L. (+)-Agelasine D: Improved synthesis and evaluation of antibacterial and cytotoxic activities. J. Nat. Prod. 2006, 69, 381–386. [Google Scholar] [CrossRef]
- Roggen, H.; Charnock, R.; Burman, C.; Felth, R.; Larsson, J.; Bohlin, L.; Gundersen, L.L. Antimicrobial and antineoplastic activities of agelasine analogs modified in the purine 2-position. Arch. Pharm. 2011, 344, 50–55. [Google Scholar] [CrossRef]
- Proszenyák, A.; Charnock, C.; Hedner, E.; Larsson, R.; Bohlin, L.; Gundersen, L.L. Synthesis, antimicrobial and antineoplastic activities for agelasine and agelasimine analogs with a β-cyclocitral derived substituent. Arch. Pharm. 2007, 340, 625–634. [Google Scholar] [CrossRef]
- Bakkestuen, A.K.; Gundersen, L.L.; Petersen, D.; Utenova, B.T.; Vik, A. Synthesis and antimycobacterial activity of agelasine E. and analogs. Org. Biomol. Chem. 2005, 3, 1025–1033. [Google Scholar] [CrossRef]
- Chamgordani, E.J.; Paulsen, J.; Gundersen, L.L. Selective N-7 alkylation of 3-methylhypoxanthine; the first synthesis of malonganenone J. Tetrahedron Lett. 2016, 57, 4926–4929. [Google Scholar] [CrossRef]
- Arai, M.; Yamano, Y.; Setiawan, A.; Kobayashi, M. Identification of the target protein of agelasine D, a marine sponge diterpene alkaloid, as an anti-dormant mycobacterial substance. ChemBioChem 2014, 15, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Wu, H.; Ohizumi, Y.; Hirata, Y. Agelasine-A, -B, -C and -D, novel bicyclic diterpenoids with a 9-methyladeninium unit possessing inhibitory effects on na,K-atpase from the okinawa sea sponge Agelas sp.1). Tetrahedron Lett. 1984, 25, 2989–2992. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nakamura, H.; Wu, H.; Kobayashi, J.; Ohizumi, Y. Mode of inhibition of brain Na+,K+-ATPase by agelasidines and agelasines from a sea sponge. Arch. Biochem. Biophys. 1987, 259, 179–184. [Google Scholar] [CrossRef]
- Fischer, W.; Urban, N.; Immig, K.; Franke, H.; Schaefer, M. Natural compounds with P2X7 receptor-modulating properties. Purinergic Signal. 2014, 10, 313–326. [Google Scholar] [CrossRef]
- Pimentel, A.A.; Felibertt, P.; Sojo, F.; Colman, L.; Mayora, A.; Silva, M.L.; Rojas, H.; Dipolo, R.; Suarez, A.I.; Compagnone, R.S.; et al. The marine sponge toxin agelasine B increases the intracellular Ca2+ concentration and induces apoptosis in human breast cancer cells (MCF-7). Cancer Chemother. Pharmacol. 2012, 69, 71–83. [Google Scholar] [CrossRef]
- Zhang, J.R.; Li, P.L.; Tang, X.L.; Qi, X.; Li, G.Q. Cytotoxic tetraprenylated alkaloids from the South China sea gorgonian Euplexaura robusta. Chem. Biodivers. 2012, 9, 2218–2224. [Google Scholar] [CrossRef]
- Cockburn, I.L.; Pesce, E.R.; Pryzborski, J.M.; Davies-Coleman, M.T.; Clark, P.G.K.; Keyzers, R.A.; Stephens, L.L.; Blatch, G.L. Screening for small molecule modulators of Hsp70 chaperone activity using protein aggregation suppression assays: Inhibition of the plasmodial chaperone PfHsp70-1. Biol. Chem. 2011, 2, 431–438. [Google Scholar] [CrossRef]
- Furse, S.; Brooks, N.J.; Woscholski, R.; Gaffney, P.R.J.; Templer, R.H. Pressure-dependent inverse bicontinuous cubic phase formation in a phosphatidylinositol 4-phosphate/phosphatidylcholine system. Chem. Data Collect. 2016, 4, 15–20. [Google Scholar] [CrossRef]
- Escribá, P.V. Membrane-lipid therapy: A historical perspective of membrane-targeted therapies-From lipid bilayer structure to the pathophysiological regulation of cells. Biochim. Biophys. Acta-Biomembr. 2017, 1859, 1493–1506. [Google Scholar] [CrossRef]
- Bakrač, B.; Gutiérrez-Aguirre, I.; Podlesek, Z.; Sonnen, A.F.P.; Gilbert, R.J.C.; Maček, P.; Lakey, J.H.; Anderluh, G. Molecular determinants of sphingomyelin specificity of a eukaryotic pore-forming toxin. J. Biol. Chem. 2008, 283, 18665–18677. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Dalla Serra, M.; Ferreras, M.; Lanio, M.E.; Menestrina, G. Mechanism of membrane permeabilization by sticholysin I, a cytolysin isolated from the venom of the sea anemone Stichodactyla helianthus. Biochemistry 1996, 35, 14947–14957. [Google Scholar] [CrossRef] [PubMed]
- Valcarcel, C.A.; Dalla Serra, M.; Potrich, C.; Bernhart, I.; Tejuca, M.; Martinez, D.; Pazos, F.; Lanio, M.E.; Menestrina, G. Effects of Lipid Composition on Membrane Permeabilization by Sticholysin I and II, Two Cytolysins of the Sea Anemone Stichodactyla helianthus. Biophys. J. 2001, 80, 2761–2774. [Google Scholar] [CrossRef]
- Wen, H.; Glomm, W.R.; Halskau, Ø. Cytotoxicity of bovine α-lactalbumin: Oleic acid complexes correlates with the disruption of lipid membranes. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 2691–2699. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nielsen, S.B.; Wilhelm, K.; Vad, B.; Schleucher, J.; Morozova-Roche, L.A.; Otzen, D. The interaction of equine lysozyme: Oleic acid complexes with lipid membranes suggests a cargo off-loading mechanism. J. Mol. Biol. 2010, 8, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Mulet, X.; Templer, R.H.; Woscholski, R.; Ces, O. Evidence that phosphatidylinositol promotes curved membrane interfaces. Langmuir 2008, 24, 8443–8447. [Google Scholar] [CrossRef] [PubMed]
- Furse, S.; Jakubec, M.; Rise, F.; Williams, H.E.; Rees, C.E.D.; Halskau, O. Evidence that Listeria innocua modulates its membrane’s stored curvature elastic stress, but not fluidity, through the cell cycle. Sci. Rep. 2017, 7, 8012. [Google Scholar] [CrossRef]
- Furse, S.; Brooks, N.J.; Seddon, A.M.; Woscholski, R.; Templer, R.H.; Tate, E.W.; Gaffney, P.R.J.; Ces, O. Lipid membrane curvature induced by distearoyl phosphatidylinositol 4-phosphate. Soft Matter 2012, 8, 3090–3093. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the phosphatidylcholines. Biochim. Biophys. Acta-Rev. Biomembr. 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the hydrated phosphatidylethanolamines. Chem. Phys. Lipids 1994, 69, 1–34. [Google Scholar] [CrossRef]
- Totland, C.; Nerdal, W.; Steinkopf, S. Effects and Location of Coplanar and Noncoplanar PCB in a Lipid Bilayer: A Solid-State NMR Study. Environ. Sci. Technol. 2016, 50, 8290–8295. [Google Scholar] [CrossRef] [PubMed]
- Dave, P.C.; Tiburu, E.K.; Damodaran, K.; Lorigan, G.A. Investigating Structural Changes in the Lipid Bilayer upon Insertion of the Transmembrane Domain of the Membrane-Bound Protein Phospholamban Utilizing 31P and 2H Solid-State NMR Spectroscopy. Biophys. J. 2004, 86, 1564–1573. [Google Scholar] [CrossRef][Green Version]
- Sampaio, J.L.; Gerl, M.J.; Klose, C.; Ejsing, C.S.; Beug, H.; Simons, K.; Shevchenko, A. Membrane lipidome of an epithelial cell line. Proc. Natl. Acad. Sci. USA 2011, 108, 1903–1907. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef]
- Furse, S.; Liddell, S.; Ortori, C.A.; Williams, H.; Neylon, D.C.; Scott, D.J.; Barrett, D.A.; Gray, D.A. The lipidome and proteome of oil bodies from Helianthus annuus (common sunflower). J. Chem. Biol. 2013, 6, 63–76. [Google Scholar] [CrossRef]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef]
- Furse, S.; Wienk, H.; Boelens, R.; De Kroon, A.I.P.M.; Killian, J.A. E. coli MG1655 modulates its phospholipid composition through the cell cycle. FEBS Lett. 2015, 589, 2726–2730. [Google Scholar] [CrossRef]
- Hui, S.W.; Stewart, T.P.; Yeagle, P.L.; Albert, A.D. Bilayer to non-bilayer transition in mixtures of phosphatidylethanolamine and phosphatidylcholine: Implications for membrane properties. Arch. Biochem. Biophys. 1981, 207, 227–240. [Google Scholar] [CrossRef]
- Chapman, D.; Owens, N.F.; Walker, D.A. Physical studies of phospholipids II. Monolayer studies of some synthetic 2,3-diacyl-dl-phosphatidylethanolamines and phosphatidylcholines containing trans double bonds. BBA-Biophys. Incl. Photosynth. 1966, 120, 148–155. [Google Scholar] [CrossRef]
- Setiawan, I.; Blanchard, G.J. Ethanol-induced perturbations to planar lipid bilayer structures. J. Phys. Chem. B 2014, 118, 537–546. [Google Scholar] [CrossRef]
- Komatsu, H.; Okada, S. Ethanol-enhanced permeation of phosphatidylcholine/phosphatidylethanolamine mixed liposomal membranes due to ethanol-induced lateral phase separation. Biochim. Biophys. Acta-Biomembr. 1996, 1283, 73–79. [Google Scholar] [CrossRef]
- Dubovskii, P.V.; Lesovoy, D.M.; Dubinnyi, M.A.; Konshina, A.G.; Utkin, Y.N.; Efremov, R.G.; Arseniev, A.S. Interaction of three-finger toxins with phospholipid membranes: Comparison of S- and P-type cytotoxins. Biochem. J. 2005, 387, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Mcintyre, S.K.; Alam, T.M. Distinguishing Individual Lipid Headgroup Mobility and Phase Transitions in Raft-Forming Lipid Mixtures with 31 P. MAS NMR. Biophys. J. 2006, 90, 4248–4260. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, C.R.; Thiel, S.; Otzen, D.E. Protein-fatty acid complexes: Biochemistry, biophysics and function. FEBS J. 2013, 280, 1733–1749. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Strømland, Ø.; Halskau, Ø.; Stromland, O.; Halskau, O. Alpha-Lactalbumin:Oleic Acid Complex Spontaneously Delivers Oleic Acid to Artificial and Erythrocyte Membranes. J. Mol. Biol. 2015, 427, 3177–3187. [Google Scholar] [CrossRef] [PubMed]
- Chehimi, S.; Pons, A.-M.; Sablé, S.; Hajlaoui, M.-R.; Limam, F. Mode of action of thuricin S, a new class IId bacteriocin from Bacillus thuringiensis. Can. J. Microbiol. 2010, 56, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Drusano, G.L. Antimicrobial pharmacodynamics: Critical interactions of “bug and drug”. Nat. Rev. Microbiol. 2004, 2, 289–300. [Google Scholar] [CrossRef]
- Yang, Y.; Yao, H.; Hong, M. Distinguishing bicontinuous lipid cubic phases from isotropic membrane morphologies using 31P solid-state NMR spectroscopy. J. Phys. Chem. B. 2015, 119, 4993–5001. [Google Scholar] [CrossRef]
- Paulsen, B.; Fredriksen, K.A.; Petersen, D.; Maes, L.; Matheeussen, A.; Naemi, A.O.; Scheie, A.A.; Simm, R.; Ma, R.; Wan, B.; et al. Synthesis and antimicrobial activities of N 6 -hydroxyagelasine analogs and revision of the structure of ageloximes. Bioorganic Med. Chem. 2019, 27, 620–629. [Google Scholar] [CrossRef]




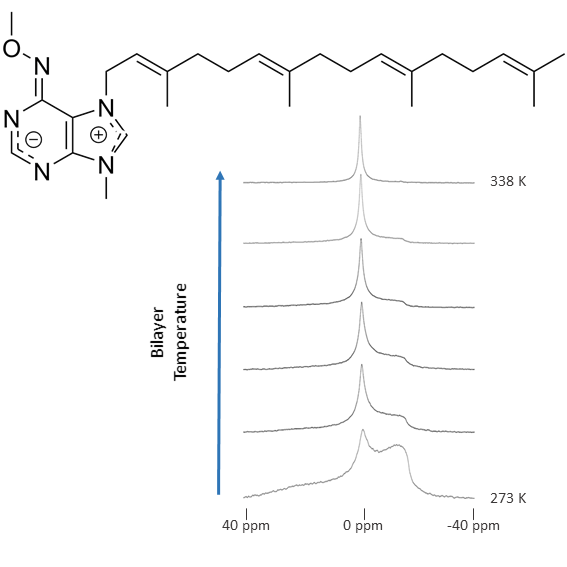{kind=link}
{kind=link}
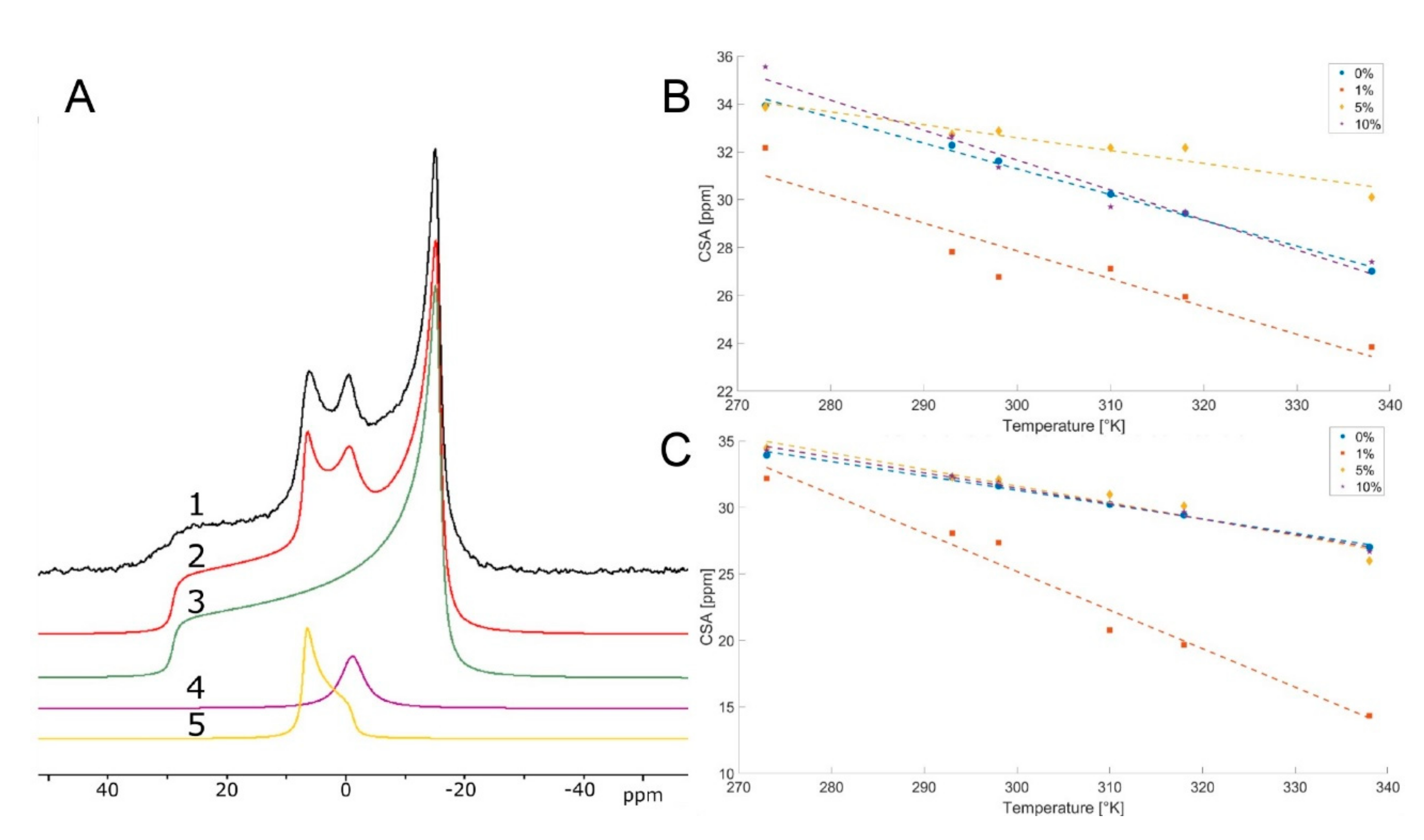{kind=link}
{kind=link}
| Compound | Bacteria MIC (µg/mL) | Fungi MIC (µg/mL) | Cancer Cells IC50 (µg/mL) | Protozoa IC50 (µg/mL) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Staphylococcus aureus | Streptococcus pyogenes | Enterococcus faecalis | Escherichia coli | Pseudomonas aeruginosa | Bacteroides fragilis | Bacteroides thetaiotaomicron | Mycobacterium tuberculosis | Candida krusei | U-937GTB | RPMI8226/s | CEM7S | ACHN | Plasmodium falciparum | Leishmania infantum | Trypanosoma cruzi | Trypanosoma bruceii | |
| 1 | 1 c | 2 c | 8 c | 8 c | 16 c | 16 c | 8–16 c | >6.25 c 92% at 6.25 µg/mL | – | 2.6 c | 1.8 c | 2.3 c | 11.8 c | 0.3 a | 1.5 a | 4.5 a | 0.9 a |
| 2 | 2 c | 2 c | 8 c | 8 c | 32 c | 4–8 c | 4–8 c | >6.25 c 96% at 6.25 µg/mL | – | 0.5 c | 0.1 c | 1.1 c | 3.7 c | 0.3 a | 0.6 a | 0.5 a | 0.3 a |
| 3 | 32 b | – | – | >32 d | – | – | – | >6.25 b 38% at 6.25 µg/mL | >16 b | 1.5 b | 1.3 b | 1.5 b | 9.6 b | – | – | – | – |
| 4 | 4 b | 4 b | 8 b | 16 b | >32 b | 8 b | 8 b | 3.13 b | 2.0 b | 0.7 b | 0.5 b | 0.9 b | 3.6 b | 0.1 a | 0.1 a | 0.1 a | 0.2 a |
| 5 | 32 d | – | – | >32d | – | – | – | – | – | – | – | – | – | – | 4.6 d | 0.9 d | 4.5 d |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubec, M.; Totland, C.; Rise, F.; Chamgordani, E.J.; Paulsen, B.; Maes, L.; Matheeussen, A.; Gundersen, L.-L.; Halskau, Ø. Bioactive Metabolites of Marine Origin Have Unusual Effects on Model Membrane Systems. Mar. Drugs 2020, 18, 125. https://doi.org/10.3390/md18020125
Jakubec M, Totland C, Rise F, Chamgordani EJ, Paulsen B, Maes L, Matheeussen A, Gundersen L-L, Halskau Ø. Bioactive Metabolites of Marine Origin Have Unusual Effects on Model Membrane Systems. Marine Drugs. 2020; 18(2):125. https://doi.org/10.3390/md18020125
Chicago/Turabian StyleJakubec, Martin, Christian Totland, Frode Rise, Elahe Jafari Chamgordani, Britt Paulsen, Louis Maes, An Matheeussen, Lise-Lotte Gundersen, and Øyvind Halskau. 2020. "Bioactive Metabolites of Marine Origin Have Unusual Effects on Model Membrane Systems" Marine Drugs 18, no. 2: 125. https://doi.org/10.3390/md18020125
APA StyleJakubec, M., Totland, C., Rise, F., Chamgordani, E. J., Paulsen, B., Maes, L., Matheeussen, A., Gundersen, L.-L., & Halskau, Ø. (2020). Bioactive Metabolites of Marine Origin Have Unusual Effects on Model Membrane Systems. Marine Drugs, 18(2), 125. https://doi.org/10.3390/md18020125