Chitooligosaccharides Modulate Glucose-Lipid Metabolism by Suppressing SMYD3 Pathways and Regulating Gut Microflora

 ,
,
Abstract
1. Introduction
2. Results
2.1. COS Can Ameliorate Hyperglycemia and Dyslipidemia Induced by T2DM
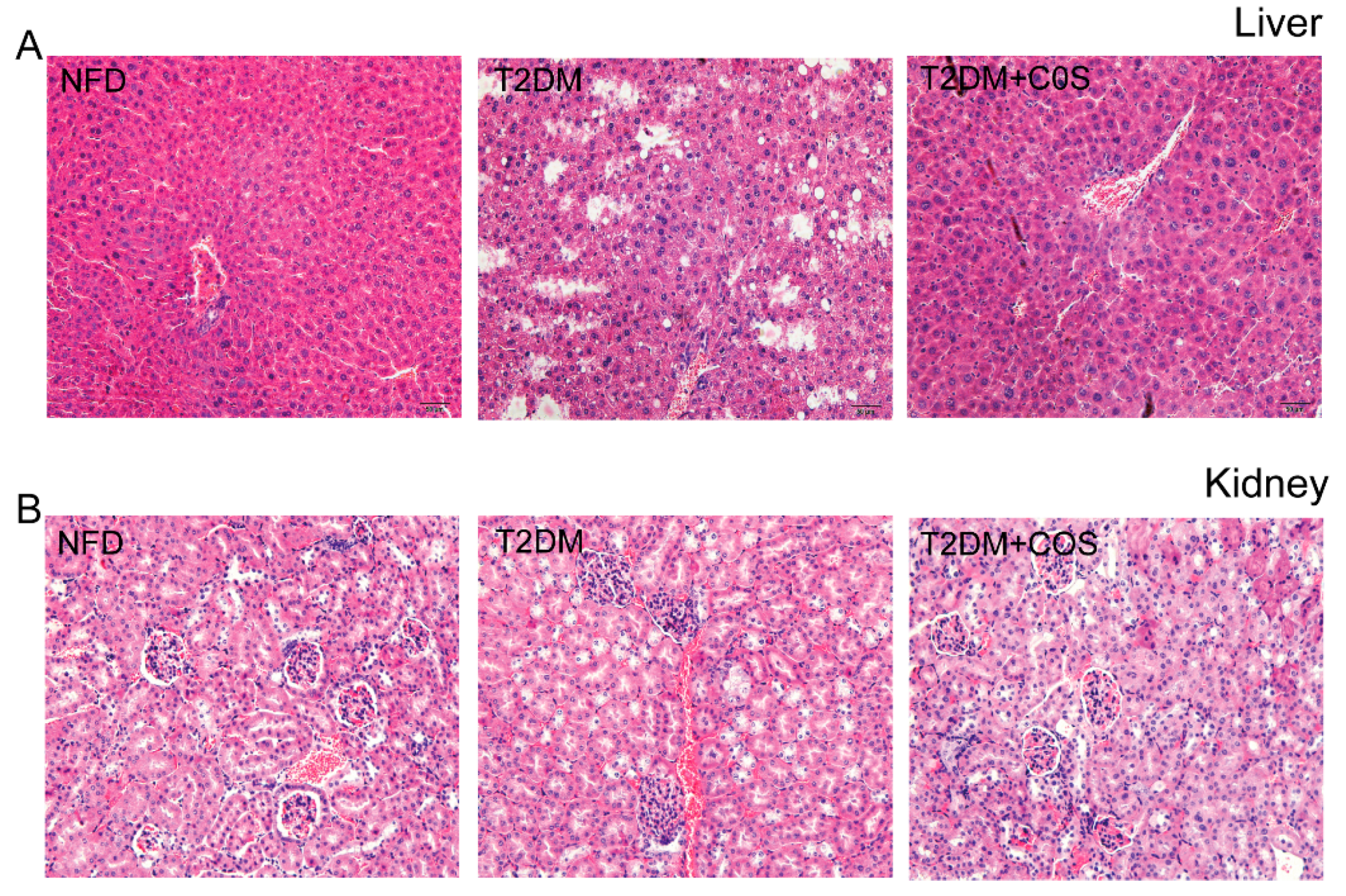
2.2. COS Has Potential Protection Effects on Liver and Renal Damages of Type 2 Diabetic Mice
2.3. COS Altered the T2DM-Induced Gut Microflora Dysbiosis
2.4. COS Regulated Glucose-Lipid Metabolism in Liver and Renal Pathology
2.5. COS-Regulated Lipid Metabolism in the HepG2 Steatosis Model
2.6. SMYD3 Activates HMGCR Transcription via SBE Elements in the Promoter
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Establishment of High-Fat Cell Model
4.2. Cell Transfection
4.3. Animals and Treatment
4.4. Extraction of Total RNA and RT-qPCR
4.5. Western Blotting
4.6. Luciferase Reporter Assay
4.7. Chromatin Immunoprecipitation (CHIP)
4.8. Oil Red O Staining
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xu, N.; Shi, D.Z.; Peng, Q.J.; Wu, B.; Huang, G.S.; Yang, L.C.; Huang, Y. Clinical study on acupuncture treatment of dyslipidemia and hyperglycemia. Chin. J. Clin. 2013, 41, 26–28. [Google Scholar]
- Shen, Z.H.; Lu, Y.; Fang, N.Y. New progress in the study of metabolic syndrome. Chin. J. Cardiovasc. Res. 2008, 6, 466–467. [Google Scholar]
- Gilbert, E.R.; Fu, Z.; Liu, D. Development of a nongenetic mouse model of type 2 diabetes. Exp. Diabetes Res. 2011, 2011, 416254. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Rajapakse, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydr. Polym. 2005, 62, 357–368. [Google Scholar] [CrossRef]
- Easton, J.D.; Saver, J.L.; Albers, G.W.; Alberts, M.J.; Chaturvedi, S.; Feldmann, E.; Hatsukami, T.S.; Higashida, R.T.; Johnston, S.C.; Kidwell, C.S.; et al. Definition and evaluation of transient ischemic attack: A scientific statement for healthcare professionals from the American heart association/American stroke association stroke council; council on cardiovascular surgery and anesthesia; council on cardio. Stroke 2009, 40, 2276–2293. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Benjamin, I.J.; Burke, G.L.; Chait, A.; Eckel, R.H.; Howard, B.V.; Mitch, W.; Smith, S.C.; Sowers, J.R. Diabetes and cardiovascular disease: A statement for healthcare professionals from the american heart association. Circulation 1999, 100, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Lovett, J.K.; Dennis, M.S.; Sandercock, P.A.G.; Bamford, J.; Warlow, C.P.; Rothwell, P.M. Very Early Risk of Stroke After a First Transient Ischemic Attack. Stroke 2003, 34, e138–e140. [Google Scholar] [CrossRef]
- Kleindorfer, D.; Panagos, P.; Pancioli, A.; Khoury, J.; Kissela, B.; Woo, D.; Schneider, A.; Alwell, K.; Jauch, E.; Miller, R.; et al. Incidence and short-term prognosis of transient ischemic attack in a population-based study. Stroke 2005, 36, 720–723. [Google Scholar] [CrossRef]
- Du, Y.Z.; Wang, L.; Yuan, H.; Wei, X.H.; Hu, F.Q. Preparation and characteristics of linoleic acid-grafted chitosan oligosaccharide micelles as a carrier for doxorubicin. Colloids Surf. B Biointerfaces 2009, 69, 257–263. [Google Scholar] [CrossRef]
- Berger, J.; Reist, M.; Mayer, J.M.; Felt, O.; Gurny, R. Structure and interactions in chitosan hydrogels formed by complexation or aggregation for biomedical applications. Eur. J. Pharm. Biopharm. 2004, 57, 35–52. [Google Scholar] [CrossRef]
- Crini, G. Recent developments in polysaccharide-based materials used as adsorbents in wastewater treatment. Prog. Polym. Sci. 2005, 30, 38–70. [Google Scholar] [CrossRef]
- Sumiyoshi, M.; Kimura, Y. Low molecular weight chitosan inhibits obesity induced by feeding a high-fat diet long-term in mice. J. Pharm. Pharmacol. 2006, 58, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Ma, P.; Yu, W.; Tan, C.; Liu, H.; Xiong, C.; Qiao, Y.; Du, Y. Chitooligosaccharides protect human embryonic hepatocytes against oxidative stress induced by hydrogen peroxide. Mar. Biotechnol. 2010, 12, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Bahar, B.; O’Doherty, J.V.; O’Doherty, A.M.; Sweeney, T. Chito-Oligosaccharide Inhibits the De-Methylation of a “CpG” Island within the Leptin (LEP) Promoter during Adipogenesis of 3T3-L1 Cells. PLoS ONE 2013, 8, e60011. [Google Scholar] [CrossRef]
- Jiang, Y.; Fu, C.; Liu, G.; Guo, J.; Su, Z. Cholesterol-lowering effects and potential mechanisms of chitooligosaccharide capsules in hyperlipidemic rats. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Mejía-León, M.E.; Petrosino, J.F.; Ajami, N.J.; Domínguez-Bello, M.G.; De La Barca, A.M.C. Fecal microbiota imbalance in Mexican children with type 1 diabetes. Sci. Rep. 2014, 4, 3814. [Google Scholar] [CrossRef]
- Van Aller, G.S.; Reynoird, N.; Barbash, O.; Huddleston, M.; Liu, S.; Zmoos, A.F.; McDevitt, P.; Sinnamon, R.; Le, B.C.; Mas, G.; et al. Smyd3 regulates cancer cell phenotypes and catalyzes histone H4 lysine 5 methylation. Epigenetics 2012, 7, 340–343. [Google Scholar] [CrossRef]
- Hamamoto, R.; Furukawa, Y.; Morita, M.; Iimura, Y.; Silva, F.P.; Li, M.; Yagyu, R.; Nakamura, Y. SMYD3 encodes a histone methyltransferase involved in the proliferation of cancer cells. Nat. Cell Biol. 2004, 6, 731–740. [Google Scholar] [CrossRef]
- Hamamoto, R.; Silva, F.P.; Tsuge, M.; Nishidate, T.; Katagiri, T.; Nakamura, Y.; Furukawa, Y. Enhanced SMYD3 expression is essential for the growth of breast cancer cells. Cancer Sci. 2006, 97, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Sims, R.J.; Reinberg, D. From chromatin to cancer: A new histone lysine methyltransferase enters the mix. Nat. Cell Biol. 2004, 6, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.D.; Pierce, S.A.; Sims, R.J.; Yamagishi, H.; Weihe, E.K.; Harriss, J.V.; Maika, S.D.; Kuziel, W.A.; King, H.L.; Olson, E.N.; et al. Bop encodes a muscle-restricted protein containing MYND and SET domains and is essential for cardiac differentiation and morphogenesis. Nat. Genet. 2002, 31, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Perez-Burgos, L.; Placek, B.J.; Sengupta, R.; Richter, M.; Dorsey, J.A.; Kubicek, S.; Opravil, S.; Jenuwein, T.; Berger, S.L. Repression of p53 activity by Smyd2-mediated methylation. Nature 2006, 444, 629–632. [Google Scholar] [CrossRef]
- Luo, X.G.; Xi, T.; Guo, S.; Liu, Z.P.; Wang, N.; Jiang, Y.; Zhang, T.C. Effects of SMYD3 overexpression on transformation, serum dependence, and apoptosis sensitivity in NIH3T3 cells. IUBMB Life 2009, 61, 679–684. [Google Scholar] [CrossRef]
- Luo, X.-G.; Ding, Y.; Zhou, Q.-F.; Ye, L.; Wang, S.-Z.; Xi, T. SET and MYND domain-containing protein 3 decreases sensitivity to dexamethasone and stimulates cell adhesion and migration in NIH3T3 cells. J. Biosci. Bioeng. 2009, 103, 582. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, X.; Deng, J.; Pan, Y.; Zhang, L.; Liang, H. SMYD3 overexpression was a risk factor in the biological behavior and prognosis of gastric carcinoma. Tumor Biol. 2015, 36, 2685–2694. [Google Scholar] [CrossRef]
- Liu, Y.; Deng, J.; Luo, X.; Pan, Y.; Zhang, L.; Zhang, R.; Liang, H. Overexpression of SMYD3 was associated with increased STAT3 activation in gastric cancer. Med. Oncol. 2015, 32, 404. [Google Scholar] [CrossRef]
- Du, S.J.; Tan, X.; Zhang, J. SMYD proteins: Key regulators in skeletal and cardiac muscle development and function. Anat. Rec. 2014, 297, 1650–1662. [Google Scholar] [CrossRef]
- Liu, L.; Tang, D.; Zhao, H.; Xin, X.; Aisa, H.A. Hypoglycemic effect of the polyphenols rich extract from Rose rugosa Thunb on high fat diet and STZ induced diabetic rats. J. Ethnopharmacol. 2017, 200, 174–181. [Google Scholar] [CrossRef]
- Rosenblit, P.D. Common medications used by patients with type 2 diabetes mellitus: What are their effects on the lipid profile? Cardiovasc. Diabetol. 2016, 15, 95. [Google Scholar] [CrossRef] [PubMed]
- Yabe, D.; Seino, Y. Two incretin hormones GLP-1 and GIP: Comparison of their actions in insulin secretion and β cell preservation. Prog. Biophys. Mol. Biol. 2011, 107, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Friedrichsen, B.N.; Neubauer, N.; Lee, Y.C.; Gram, V.K.; Blume, N.; Petersen, J.S.; Nielsen, J.H.; Møldrup, A. Stimulation of pancreatic β-cell replication by incretins involves transcriptional induction of cyclin D1 via multiple signalling pathways. J. Endocrinol. 2006, 188, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, M.; Mita, T.; Azuma, K.; Ebato, C.; Goto, H.; Nomiyama, T.; Fujitani, Y.; Hirose, T.; Kawamori, R.; Watada, H. Inhibition of monocyte adhesion to endothelial cells and attenuation of atherosclerotic lesion by a glucagon-like peptide-1 receptor agonist, exendin-4. Diabetes 2010, 59, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, K.; Wang, S.; Jenison, R.D.; Guo, G.L.; Klaassen, C.D.; Wan, Y.J.Y.; Zhong, X.B. Linkage disequilibrium blocks, haplotype structure, and htSNPs of human CYP7A1 gene. BMC Genet. 2006, 7, 29. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yamaguchi, R.; Perkins, G.; Hirota, K. Targeting cholesterol with β-cyclodextrin sensitizes cancer cells for apoptosis. FEBS Lett. 2015, 589, 4097–4105. [Google Scholar] [CrossRef]
- Lin, X.; Li, Z.; Gorfe, A.A. Reversible Effects of Peptide Concentration and Lipid Composition on H-Ras Lipid Anchor Clustering. Biophys. J. 2015, 109, 2467–2470. [Google Scholar] [CrossRef][Green Version]
- Lei, Y.M.K.; Nair, L.; Alegre, M.L. The interplay between the intestinal microbiota and the immune system. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 9–19. [Google Scholar] [CrossRef]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef]
- Mukaida, N. Intestinal microbiota: Unexpected alliance with tumor therapy. Immunotherapy 2014, 6, 231–233. [Google Scholar] [CrossRef] [PubMed]
- Hold, G.L. Gastrointestinal Microbiota and Colon Cancer. Dig. Dis. 2016, 34, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Adolph, T.E. Influence of the human intestinal microbiome on obesity and metabolic dysfunction. Curr. Opin. Pediatr. 2015, 27, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Zacarias, J.L.; Castro-Munozledo, F.; Kuri-Harcuch, W. Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry 1992, 97, 493–497. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| GAPDH | CGAGATCCCTCCAAAATCAA | TTCACACCCATGACGAACAT |
| HMGCR | CTCCTCCTTACTCGATAC | TAGATACACCACGCTCAT |
| SMYD3 | CCCAGTATCTCTTTGCTCAATCAC | TTACGGGTGTTGAAGGT |
| CYP7A1 | CAGAAGCATAGACCCAAGTGAT | TCGGTAGCAGAAGGCATACATC |
| GLP-1 | GATTCAGTCCCAGGCAGCGTAT | CTTTCTTGATCTTGGCGGGTGTT |
| mHMGCR | TTATGTCTTTAGGCTTGGTC | ACTCAGGGTAATCACTTGC |
| mGAPDH | ATTCAACGGCACAGTCAAGG | GCAGAAGGGGCGGAGATGA |
| mSMYD3 | CTGCTTTGAGTGTGACT | CTGGTAGATGTTGATGT |
| Fragment I | CGCTGATTTGGGTCTATG | GTGCGTTCCTTCTGCTCT |
| Fragment II | ATGGGTAAATCTCGGGAA | GGAGCGTGAGGGAAAACG |
| Fragment III | GCAGGCCCTAGTGCTGGG | AAGAGAGGATCGTTCGAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Jiang, Y.; Luo, X.; Wang, C.; Wang, N.; He, H.; Zhang, T.; Chen, L. Chitooligosaccharides Modulate Glucose-Lipid Metabolism by Suppressing SMYD3 Pathways and Regulating Gut Microflora. Mar. Drugs 2020, 18, 69. https://doi.org/10.3390/md18010069
Wang Q, Jiang Y, Luo X, Wang C, Wang N, He H, Zhang T, Chen L. Chitooligosaccharides Modulate Glucose-Lipid Metabolism by Suppressing SMYD3 Pathways and Regulating Gut Microflora. Marine Drugs. 2020; 18(1):69. https://doi.org/10.3390/md18010069
Chicago/Turabian StyleWang, Qiutong, Yajie Jiang, Xuegang Luo, Chang Wang, Nan Wang, Hongpeng He, Tongcun Zhang, and Liehuan Chen. 2020. "Chitooligosaccharides Modulate Glucose-Lipid Metabolism by Suppressing SMYD3 Pathways and Regulating Gut Microflora" Marine Drugs 18, no. 1: 69. https://doi.org/10.3390/md18010069
APA StyleWang, Q., Jiang, Y., Luo, X., Wang, C., Wang, N., He, H., Zhang, T., & Chen, L. (2020). Chitooligosaccharides Modulate Glucose-Lipid Metabolism by Suppressing SMYD3 Pathways and Regulating Gut Microflora. Marine Drugs, 18(1), 69. https://doi.org/10.3390/md18010069