Comparison of Physicochemical Characteristics and Fibril Formation Ability of Collagens Extracted from the Skin of Farmed River Puffer (Takifugu obscurus) and Tiger Puffer (Takifugu rubripes)


Abstract
1. Introduction
2. Results and Discussion
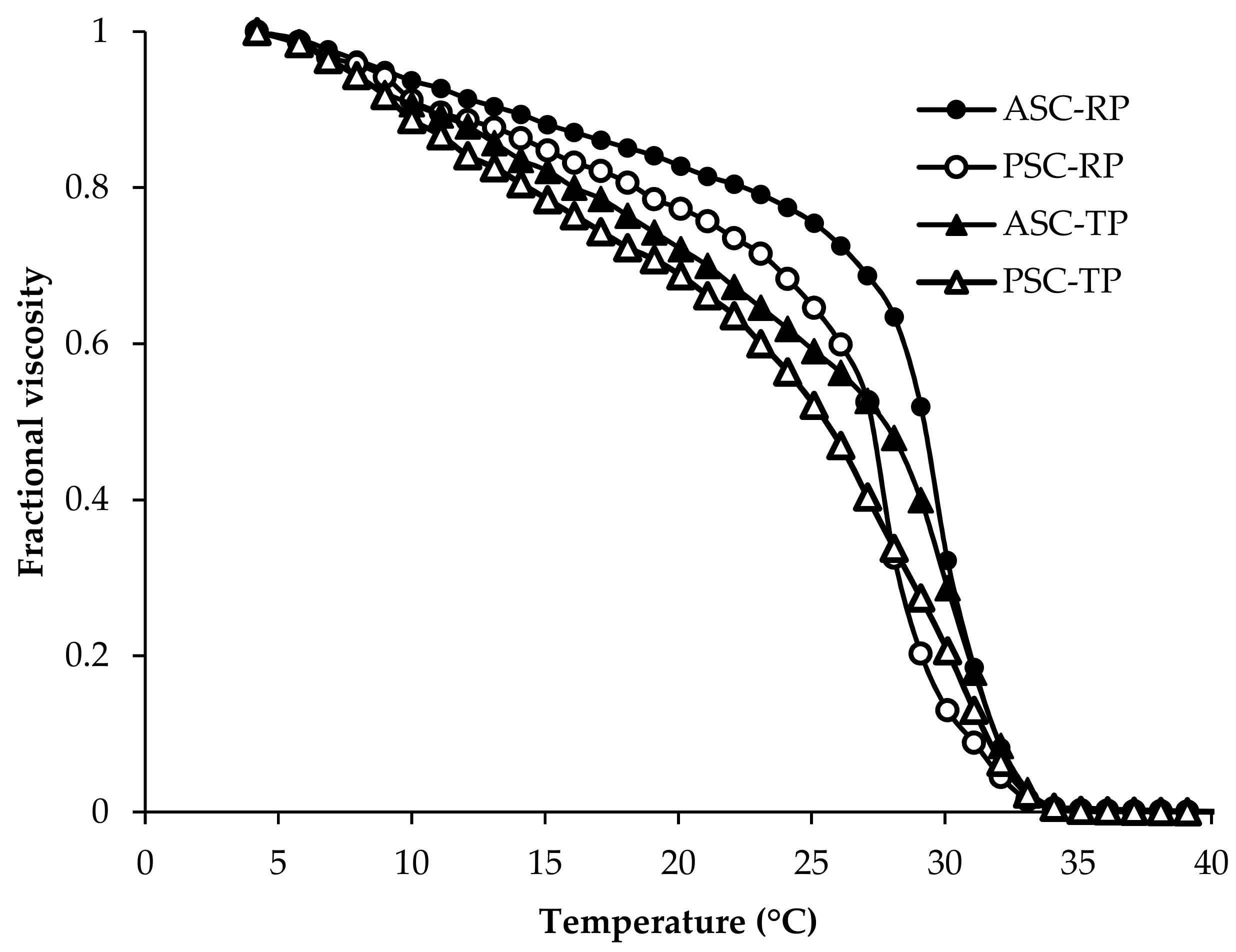
2.1. Amino Acid Composition and Thermal Denaturation Temperature (Td)
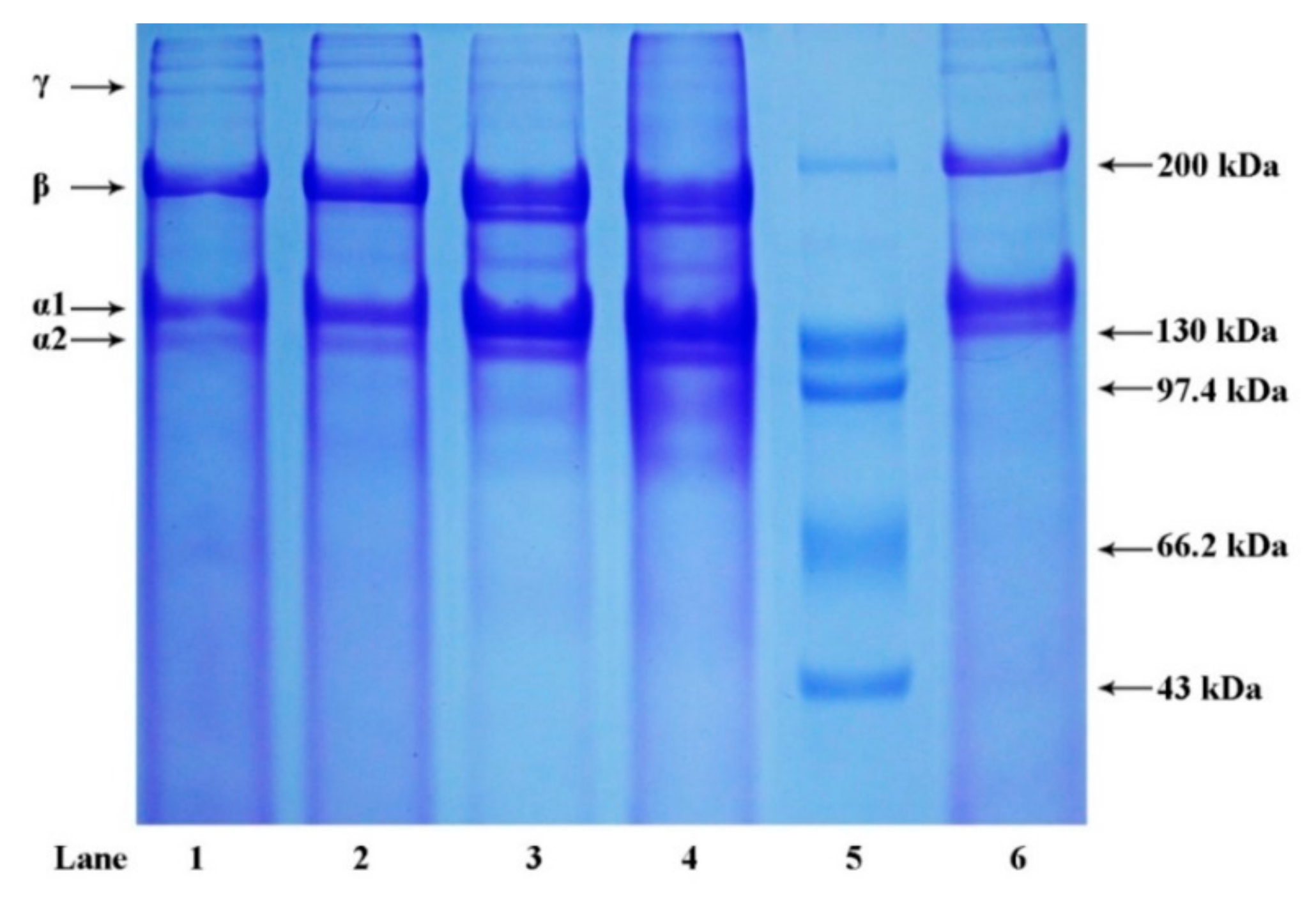
2.2. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
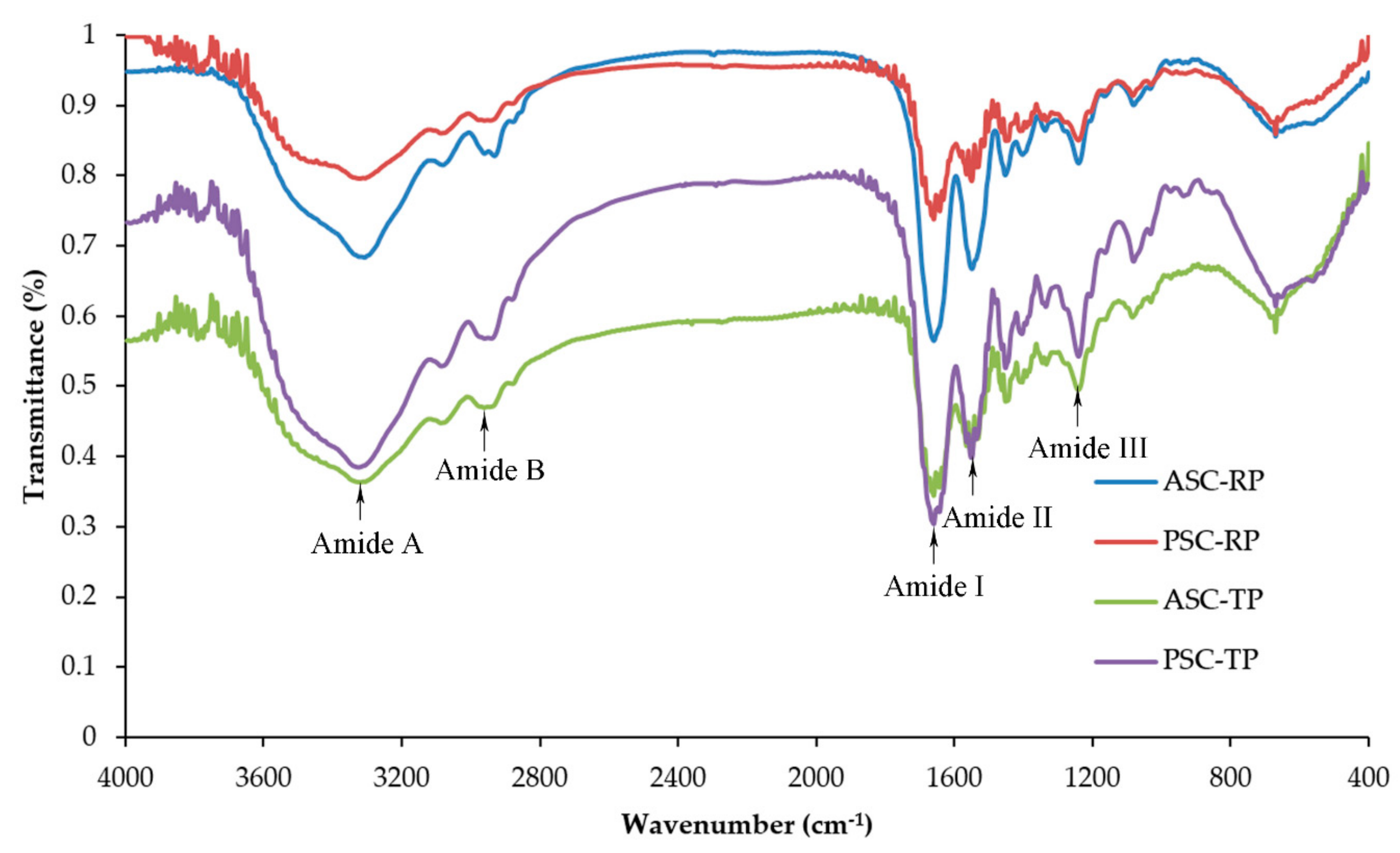
2.3. Fourier Transform Infrared Spectroscopy (FTIR)
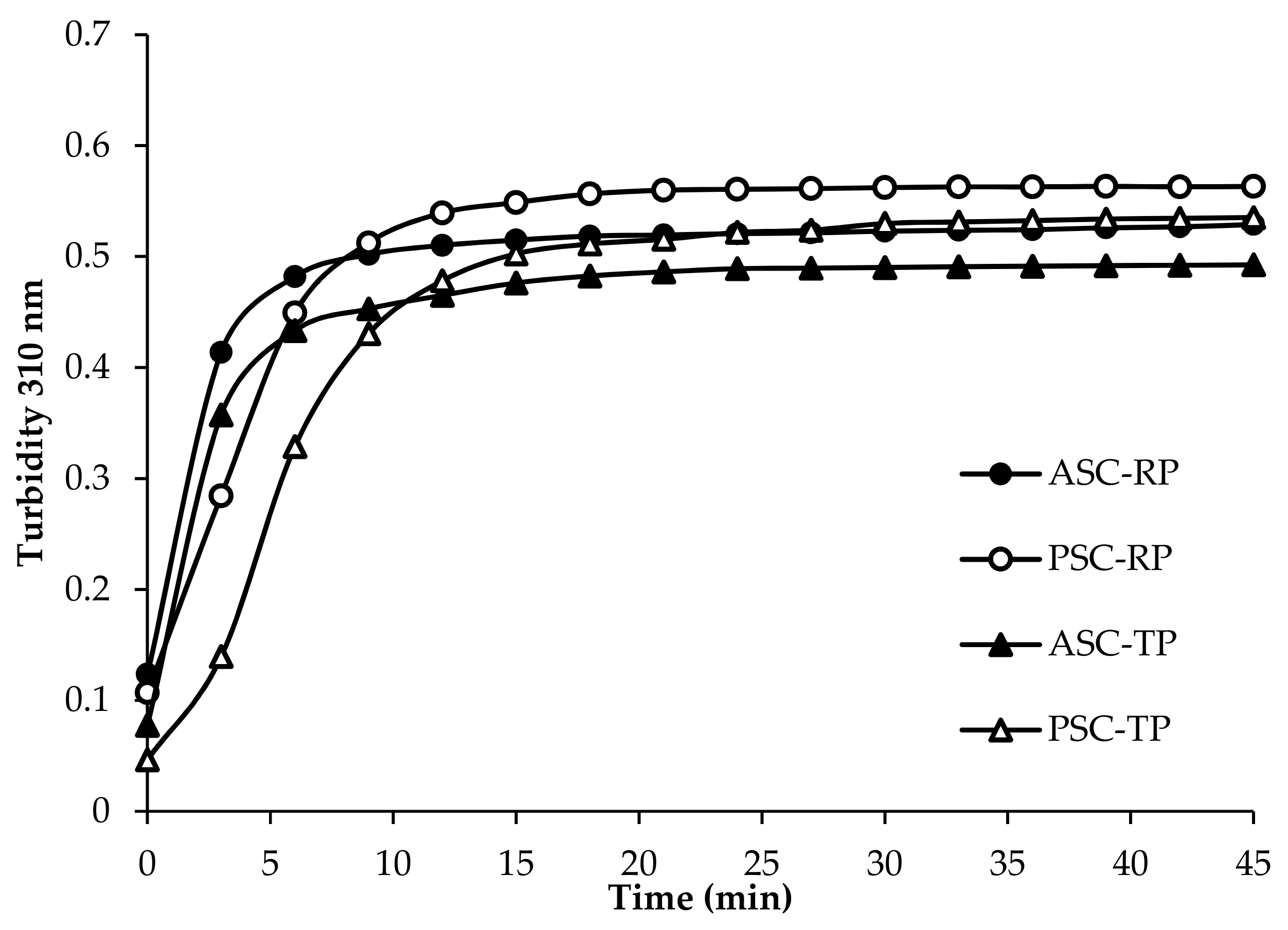
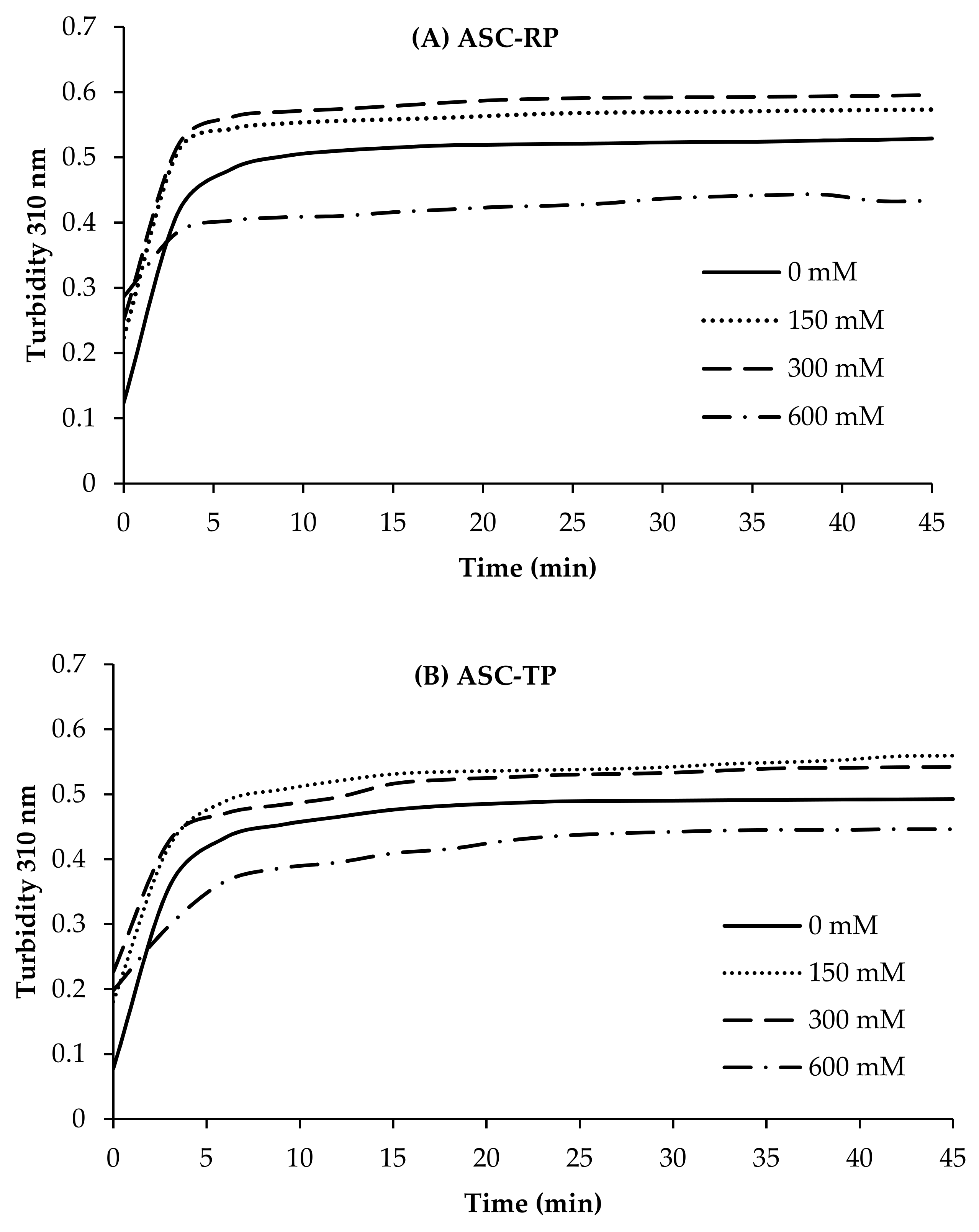
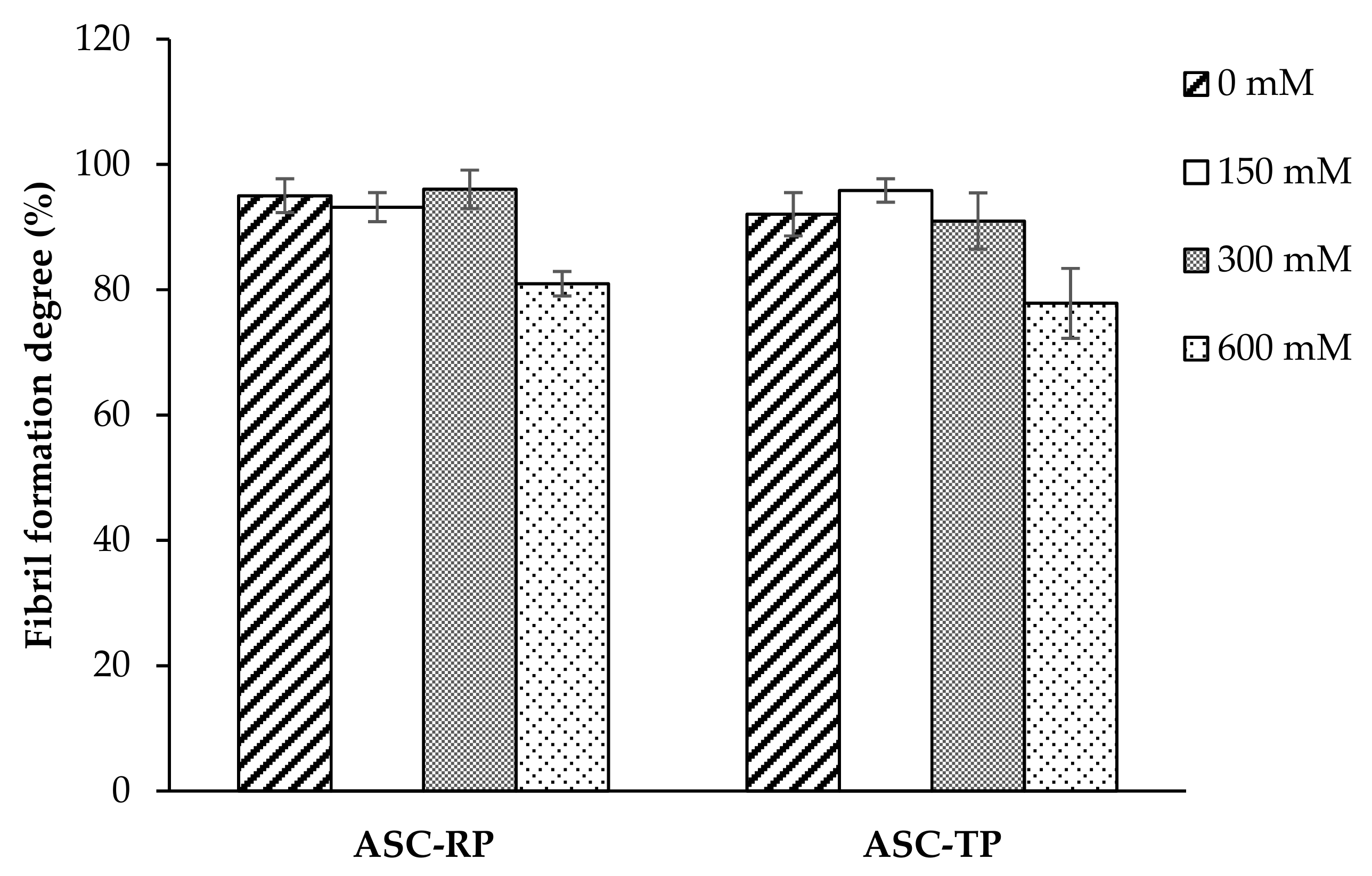
2.4. Fibril Formation Ability
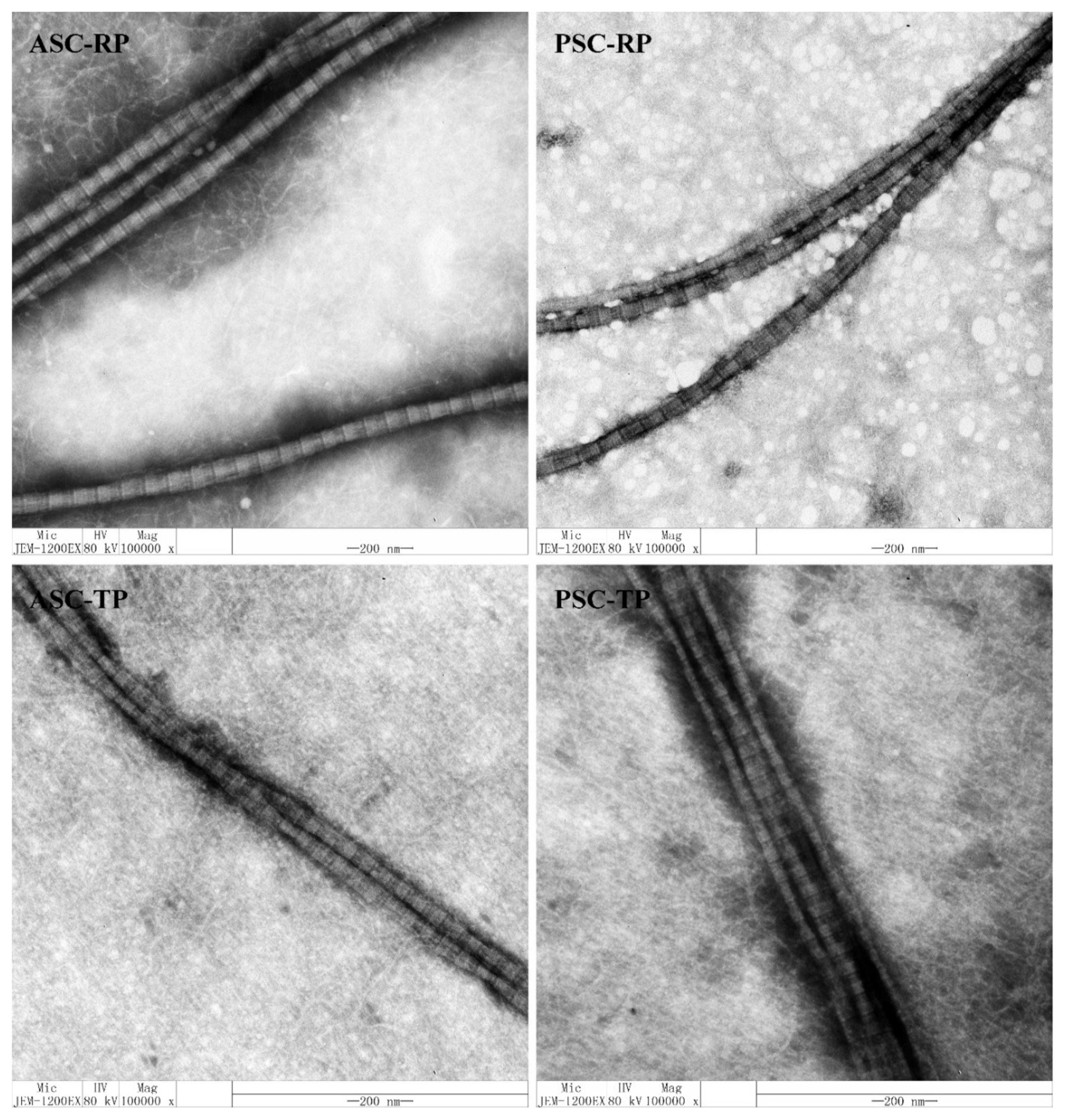
2.5. Transmission Electron Micrographs (TEM) of Collagen Fibrils
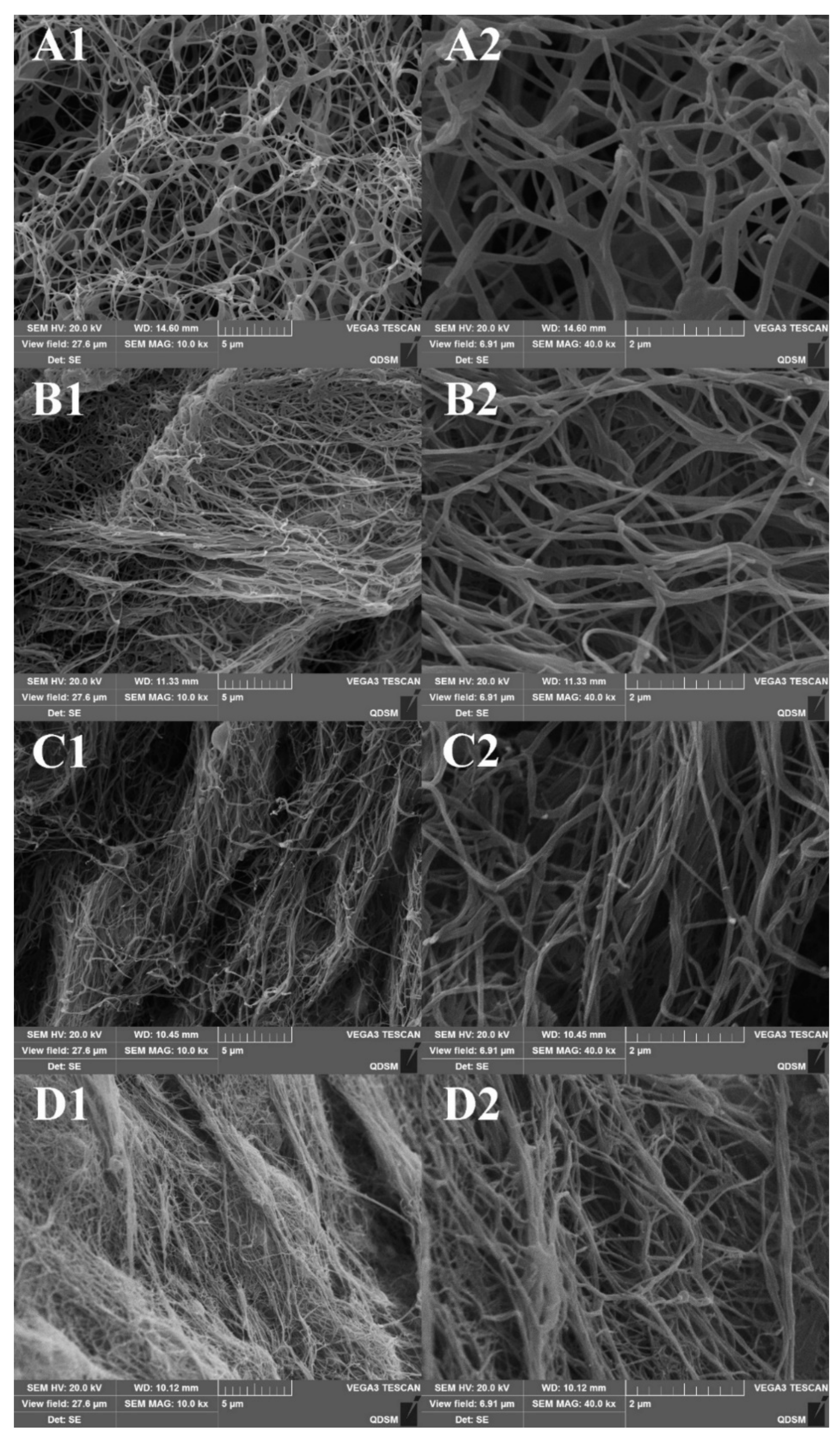
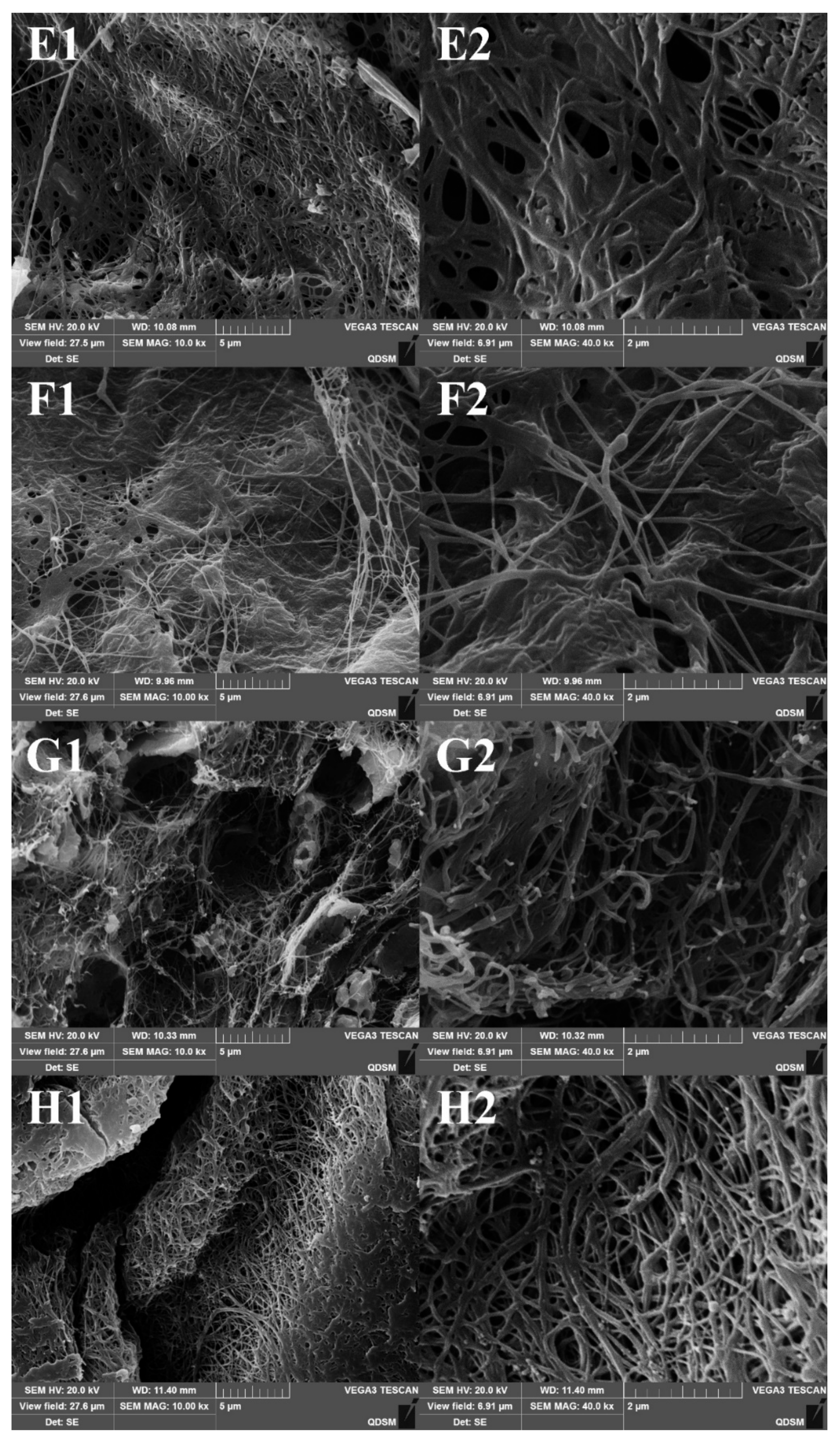
2.6. Scanning Electron Micrographs (SEM) of Collagen Fibrils
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Extraction of Acid-Soluble Collagen (ASC)
3.3. Extraction of Pepsin-Soluble Collagen (PSC)
3.4. Amino Acid Composition
3.5. Determination of Denaturation Temperature (Td)
3.6. SDS-PAGE
3.7. FTIR Spectral Analysis
3.8. Determination of Fibril Formation Ability
3.9. Transmission Electron Microscopy (TEM)
3.10. Scanning Electron Microscopy (SEM)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Noguchi, T.; Arakawa, O.; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 153–157. [Google Scholar]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar]
- Ikeda, K.; Murakami, Y.; Emoto, Y.; Ngy, L.; Taniyama, S.; Yagi, M.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to non-toxic cultured specimens of the pufferfish Takifugu rubripes. Toxicon 2009, 53, 99–103. [Google Scholar]
- Tao, N.P.; Wang, L.Y.; Gong, X.; Liu, Y. Comparison of nutritional composition of farmed pufferfish muscles among Fugu obscurus, Fugu flavidus and Fugu rubripes. J. Food Compos. Anal. 2012, 28, 40–45. [Google Scholar]
- Tao, N.P.; Wu, R.; Zhou, P.G.; Gu, S.Q.; Wu, W. Characterization of odor-active compounds in cooked meat of farmed obscure puffer (Takifugu obscurus) using gas chromatography–mass spectrometry–olfactometry. J. Food Drug Anal. 2014, 22, 431–438. [Google Scholar]
- Zhang, M.X.; Wang, X.C.; Liu, Y.; Xu, X.L.; Zhou, G.H. Isolation and identification of flavour peptides from Puffer fish (Takifugu obscurus) muscle using an electronic tongue and MALDI-TOF/TOF MS/MS. Food Chem. 2012, 135, 1463–1470. [Google Scholar]
- Zhang, X.-L. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2017; p. 22. [Google Scholar]
- Byeon, M.S.; Park, J.Y.; Yoon, S.W.; Kang, H.W. Structure and development of spines over the skin surface of the river puffer Takifugu obscurus (Tetraodontidae, Teleostei) during larval growth. J. Appl. Ichthyol. 2011, 27, 67–72. [Google Scholar] [CrossRef]
- Gelse, K. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar]
- Parenteau-Bareil, R.; Gauvin, R.; Berthod, F. Collagen-based biomaterials for tissue engineering applications. Materials 2010, 3, 1863–1887. [Google Scholar]
- Ferreira, A.M.; Gentile, P.; Chiono, V.; Ciardelli, G.; Ferreira-Duarte, A. Collagen for bone tissue regeneration. Acta Biomater. 2012, 8, 3191–3200. [Google Scholar]
- Chieruzzi, M.; Pagano, S.; Moretti, S.; Pinna, R.; Milia, E.; Torre, L.; Eramo, S. Nanomaterials for tissue engineering in dentistry. Nanomaterials 2016, 6, 134. [Google Scholar]
- Fang, M.; Goldstein, E.L.; Matich, E.K.; Orr, B.G.; Holl, M.M.B. Type I collagen self-assembly: The roles of substrate and concentration. Langmuir 2013, 29, 2330–2338. [Google Scholar]
- Shen, L.; Tian, Z.; Liu, W.; Li, G. Influence on the physicochemical properties of fish collagen gels using self-assembly and simultaneous cross-linking with the N-hydroxysuccinimide adipic acid derivative. Connect. Tissue Res. 2015, 56, 244–252. [Google Scholar]
- Petersen, S.B.; Jonson, V.; Fojan, P.; Wimmer, R.; Pedersen, S. Sorbitol prevents the self-aggregation of unfolded lysozyme leading to an up to 13 °C stabilisation of the folded form. J. Biotechnol. 2004, 114, 269–278. [Google Scholar]
- Grinnell, F.; Petroll, W.M. Cell Motility and mechanics in three-dimensional collagen matrices. Annu. Rev. Cell Dev. Biol. 2010, 26, 335–361. [Google Scholar]
- Sang, L.; Wang, X.; Chen, Z.; Lu, J.; Gu, Z.; Li, X. Assembly of collagen fibrillar networks in the presence of alginate. Carbohydr. Polym. 2010, 82, 1264–1270. [Google Scholar]
- Cozza, N.; Bonani, W.; Motta, A.; Migliaresi, C. Evaluation of alternative sources of collagen fractions from Loligo vulgaris squid mantle. Int. J. Biol. Macromol. 2016, 87, 504–513. [Google Scholar]
- Sun, L.; Hou, H.; Li, B.; Zhang, Y. Characterization of acid- and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2017, 99, 8–14. [Google Scholar]
- Tan, Y.; Chang, S.K. Isolation and characterization of collagen extracted from channel catfish (Ictalurus punctatus) skin. Food Chem. 2018, 242, 147–155. [Google Scholar]
- Blanco, M.; Vázquez, J.A.; Pérez-Martín, R.I.; Sotelo, C.G. Collagen extraction optimization from the skin of the small-spotted catshark (S. canicula) by response surface methodology. Mar. Drugs 2019, 17, 40. [Google Scholar]
- Weng, W.; Wang, Y. Partial characterization and primary structure of collagen subunits from golden pompano skins. Process. Biochem. 2018, 70, 98–103. [Google Scholar]
- Zhang, J.; Duan, R.; Huang, L.; Song, Y.; Regenstein, J.M. Characterisation of acid-soluble and pepsin-solubilised collagen from jellyfish (Cyanea nozakii Kishinouye). Food Chem. 2014, 150, 22–26. [Google Scholar]
- Rahman, A.M. Collagen of extracellular matrix from marine invertebrates and its medical applications. Mar. Drugs 2019, 17, 118. [Google Scholar] [CrossRef]
- Song, W.-K.; Liu, D.; Sun, L.-L.; Li, B.-F.; Hou, H. Physicochemical and biocompatibility properties of type I collagen from the skin of Nile Tilapia (Oreochromis niloticus) for biomedical applications. Mar. Drugs 2019, 17, 137. [Google Scholar]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and characterisation of collagen extracted from the skin of striped catfish (Pangasianodon hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar]
- Zhang, X.; Ookawa, M.; Tan, Y.; Ura, K.; Adachi, S.; Takagi, Y. Biochemical characterisation and assessment of fibril-forming ability of collagens extracted from Bester sturgeon Huso huso×Acipenser ruthenus. Food Chem. 2014, 160, 305–312. [Google Scholar]
- Tang, Y.; Jin, S.; Li, X.; Li, X.; Hu, X.; Chen, Y.; Huang, F.; Yang, Z.; Yu, F.; Ding, G. Physicochemical properties and biocompatibility evaluation of collagen from the skin of Giant Croaker (Nibea japonica). Mar. Drugs 2018, 16, 222. [Google Scholar]
- Wang, L.; An, X.; Yang, F.; Xin, Z.; Zhao, L.; Hu, Q. Isolation and characterisation of collagens from the skin, scale and bone of deep-sea redfish (Sebastes mentella). Food Chem. 2008, 108, 616–623. [Google Scholar]
- Sun, L.; Li, B.; Song, W.; Si, L.; Hou, H. Characterization of Pacific cod (Gadus macrocephalus) skin collagen and fabrication of collagen sponge as a good biocompatible biomedical material. Process. Biochem. 2017, 63, 229–235. [Google Scholar]
- El-Rashidy, A.A.; Gad, A.; Abu-Hussein, A.E.-H.G.; Habib, S.I.; Badr, N.A.; Hashem, A.A. Chemical and biological evaluation of Egyptian Nile Tilapia (Oreochromis niloticas) fish scale collagen. Int. J. Biol. Macromol. 2015, 79, 618–626. [Google Scholar]
- Zhang, J.; Duan, R. Characterisation of acid-soluble and pepsin-solubilised collagen from frog (Rana nigromaculata) skin. Int. J. Biol. Macromol. 2017, 101, 638–642. [Google Scholar]
- Yousefi, M.; Ariffin, F.; Huda, N. An alternative source of type I collagen based on by-product with higher thermal stability. Food Hydrocoll. 2017, 63, 372–382. [Google Scholar]
- Li, L.Y.; Zhao, Y.Q.; He, Y.; Chi, C.F.; Wang, B. Physicochemical and antioxidant properties of acid- and pepsin-soluble collagens from the scales of Miiuy Croaker (Miichthys Miiuy). Mar. Drugs 2018, 16, 394. [Google Scholar]
- Li, Z.R.; Wang, B.; Chi, C.F.; Zhang, Q.H.; Gong, Y.D.; Tang, J.J.; Luo, H.Y.; Ding, G.F. Isolation and characterization of acid soluble collagens and pepsin soluble collagens from the skin and bone of Spanish mackerel (Scomberomorous niphonius). Food Hydrocoll. 2013, 31, 103–113. [Google Scholar]
- Doyle, B.B.; Bendit, E.G.; Blout, E.R. Infrared spectroscopy of collagen and collagen-like polypeptides. Biopolymers 1975, 14, 937–957. [Google Scholar]
- Abe, Y.; Krimm, S. Normal vibrations of crystalline polyglycine I. Biopolymers 1972, 11, 1817–1839. [Google Scholar]
- Payne, K.J.; Veis, A. Fourier transform ir spectroscopy of collagen and gelatin solutions: Deconvolution of the amide I band for conformational studies. Biopolymers 1988, 27, 1749–1760. [Google Scholar]
- Zhao, W.H.; Chi, C.F.; Zhao, Y.Q.; Wang, B. Preparation, physicochemical and antioxidant properties of acid- and pepsin-soluble collagens from the swim bladders of Miiuy Croaker (Miichthys miiuy). Mar. Drugs 2018, 16, 161. [Google Scholar]
- Zhu, J.; Kaufman, L.J. Collagen I self-assembly: Revealing the developing structures that generate turbidity. Biophys. J. 2014, 106, 1822–1831. [Google Scholar]
- Yan, M.; Qin, S.; Li, J. Study on the self-assembly property of type I collagen prepared from tilapia (Oreochromis niloticus) skin by different extraction methods. Int. J. Food Sci. Technol. 2015, 50, 2088–2096. [Google Scholar]
- Liu, Y.; Ma, D.; Wang, Y.; Qin, W. A comparative study of the properties and self-aggregation behavior of collagens from the scales and skin of grass carp (Ctenopharyngodon idella). Int. J. Biol. Macromol. 2018, 106, 516–522. [Google Scholar]
- Li, Y.; Douglas, E.P. Effects of various salts on structural polymorphism of reconstituted type I collagen fibrils. Colloids Surf. B Biointerfaces 2013, 112, 42–50. [Google Scholar]
- Haworth, R.A.; Chapman, J.A. A study of the growth of normal and iodinated collagen fibrilsin vitro using electron microscope autoradiography. Biopolymers 1977, 16, 1895–1906. [Google Scholar]
- Doyle, B.B.; Hukins, D.W.; Hulmes, D.J.; Miller, A.; Woodhead-Galloway, J. Collagen polymorphism: Its origins in the amino acid sequence. J. Mol. Biol. 1975, 91, 79–99. [Google Scholar]
- Shen, L.; Bu, H.; Yang, H.; Liu, W.; Li, G. Investigation on the behavior of collagen self-assembly in vitro via adding sodium silicate. Int. J. Biol. Macromol. 2018, 115, 635–642. [Google Scholar]
- Liao, W.; Guanghua, X.; Li, Y.; Shen, X.R.; Li, C. Comparison of characteristics and fibril-forming ability of skin collagen from barramundi (Lates calcarifer) and tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2018, 107, 549–559. [Google Scholar]
- Freudenberg, U.; Behrens, S.H.; Welzel, P.B.; Müller, M.; Grimmer, M.; Salchert, K.; Taeger, T.; Schmidt, K.; Pompe, W.; Werner, C. Electrostatic interactions modulate the conformation of collagen, I. Biophys. J. 2007, 92, 2108–2119. [Google Scholar]
- Li, Y.; Asadi, A.; Monroe, M.R.; Douglas, E.P. pH effects on collagen fibrillogenesis in vitro: Electrostatic interactions and phosphate binding. Mater. Sci. Eng. C 2009, 29, 1643–1649. [Google Scholar]
- Pal, G.K.; Nidheesh, T.; Suresh, P. Comparative study on characteristics and in vitro fibril formation ability of acid and pepsin soluble collagen from the skin of catla (Catla catla) and rohu (Labeo rohita). Food Res. Int. 2015, 76, 804–812. [Google Scholar]
- Yunoki, S.; Nagai, N.; Suzuki, T.; Munekata, M. Novel biomaterial from reinforced salmon collagen gel prepared by fibril formation and cross-linking. J. Biosci. Bioeng. 2004, 98, 40–47. [Google Scholar]
- Hoyer, B.; Bernhardt, A.; Lode, A.; Heinemann, S.; Sewing, J.; Klinger, M.; Notbohm, H.; Gelinsky, M. Jellyfish collagen scaffolds for cartilage tissue engineering. Acta Biomater. 2014, 10, 883–892. [Google Scholar]
- Tang, L.; Chen, S.; Su, W.; Weng, W.; Osako, K.; Tanaka, M. Physicochemical properties and film-forming ability of fish skin collagen extracted from different freshwater species. Process. Biochem. 2015, 50, 148–155. [Google Scholar]
- Chen, S.; Ikoma, T.; Ogawa, N.; Migita, S.; Kobayashi, H.; Hanagata, N. In vitro formation and thermal transition of novel hybrid fibrils from type I fish scale collagen and type I porcine collagen. Sci. Technol. Adv. Mater. 2010, 11, 35001. [Google Scholar]
- Nudelman, F.; Bomans, P.H.H.; George, A.; De With, G.; Sommerdijk, N.A.J.M. The role of the amorphous phase on the biomimetic mineralization of collagen. Faraday Discuss. 2012, 159, 357–370. [Google Scholar]
- Gaurav Kumar, P.; Nidheesh, T.; Govindaraju, K.; Jyoti; Suresh, P.V. Enzymatic extraction and characterisation of a thermostable collagen from swim bladder of rohu (Labeo rohita). J. Sci. Food Agr. 2017, 97, 1451–1458. [Google Scholar] [CrossRef]
- Pagano, S.; Chieruzzi, M.; Balloni, S.; Lombardo, G.; Torre, L.; Bodo, M.; Cianetti, S.; Marinucci, L. Biological, thermal and mechanical characterization of modified glass ionomer cements: The role of nanohydroxyapatite, ciprofloxacin and zinc l-carnosine. Mater. Sci. Eng. C 2019, 94, 76–85. [Google Scholar]
- Adibzadeh, N.; Aminzadeh, S.; Jamili, S.; Karkhane, A.A.; Farrokhi, N. Purification and characterization of pepsin-solubilized collagen from skin of sea cucumber Holothuria parva. Appl. Biochem. Biotechnol. 2014, 173, 143–154. [Google Scholar]
- Usha, R.; Ramasami, T. The effects of urea and n-propanol on collagen denaturation: using DSC, circular dicroism and viscosity. Thermochim. Acta 2004, 409, 201–206. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | ASC-RP | PSC-RP | ASC-TP | PSC-TP | Calf Skin Collagen [31] | Porcine Skin Collagen [32] |
|---|---|---|---|---|---|---|
| Aspartic acid (Asp) | 46 | 44 | 45 | 47 | 45 | 44 |
| Threonine (Thr) | 22 | 23 | 22 | 21 | 18 | 16 |
| Serine (Ser) | 39 | 38 | 40 | 38 | 33 | 33 |
| Glutamic acid (Glu) | 72 | 69 | 73 | 74 | 75 | 72 |
| Glycine (Gly) | 326 | 331 | 332 | 339 | 330 | 341 |
| Alanine (Ala) | 126 | 124 | 127 | 122 | 119 | 115 |
| Cysteine (Cys) | 2 | 1 | 2 | 1 | 0 | 0 |
| Valine (Val) | 28 | 28 | 27 | 24 | 21 | 22 |
| Methionine (Met) | 9 | 11 | 10 | 14 | 6 | 6 |
| Isoleucine (Ile) | 10 | 9 | 9 | 9 | 11 | 10 |
| Leucine (Leu) | 18 | 17 | 17 | 18 | 23 | 22 |
| Tyrosine (Tyr) | 4 | 3 | 3 | 2 | 3 | 1 |
| Phenylalanine (Phe) | 10 | 15 | 10 | 13 | 3 | 12 |
| Histidine (His) | 6 | 7 | 5 | 7 | 5 | 5 |
| Lysine (Lys) | 29 | 28 | 30 | 27 | 26 | 27 |
| Arginine (Arg) | 57 | 53 | 58 | 52 | 50 | 48 |
| Proline (Pro) | 116 | 116 | 114 | 115 | 121 | 123 |
| Hydroxyproline (Hyp) | 80 | 83 | 76 | 77 | 94 | 97 |
| Imino acid (Pro + Hyp) | 196 | 199 | 190 | 192 | 215 | 220 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.-S.; Yu, Y.; Sun, Y.; Liu, N.; Zhou, D.-Q. Comparison of Physicochemical Characteristics and Fibril Formation Ability of Collagens Extracted from the Skin of Farmed River Puffer (Takifugu obscurus) and Tiger Puffer (Takifugu rubripes). Mar. Drugs 2019, 17, 462. https://doi.org/10.3390/md17080462
Wang S-S, Yu Y, Sun Y, Liu N, Zhou D-Q. Comparison of Physicochemical Characteristics and Fibril Formation Ability of Collagens Extracted from the Skin of Farmed River Puffer (Takifugu obscurus) and Tiger Puffer (Takifugu rubripes). Marine Drugs. 2019; 17(8):462. https://doi.org/10.3390/md17080462
Chicago/Turabian StyleWang, Shan-Shan, Ying Yu, Yong Sun, Nan Liu, and De-Qing Zhou. 2019. "Comparison of Physicochemical Characteristics and Fibril Formation Ability of Collagens Extracted from the Skin of Farmed River Puffer (Takifugu obscurus) and Tiger Puffer (Takifugu rubripes)" Marine Drugs 17, no. 8: 462. https://doi.org/10.3390/md17080462
APA StyleWang, S.-S., Yu, Y., Sun, Y., Liu, N., & Zhou, D.-Q. (2019). Comparison of Physicochemical Characteristics and Fibril Formation Ability of Collagens Extracted from the Skin of Farmed River Puffer (Takifugu obscurus) and Tiger Puffer (Takifugu rubripes). Marine Drugs, 17(8), 462. https://doi.org/10.3390/md17080462