In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass


 ,
, 
 , ,
, ,  ,
,  ,
, 

Abstract
1. Introduction
2. Results and Discussion
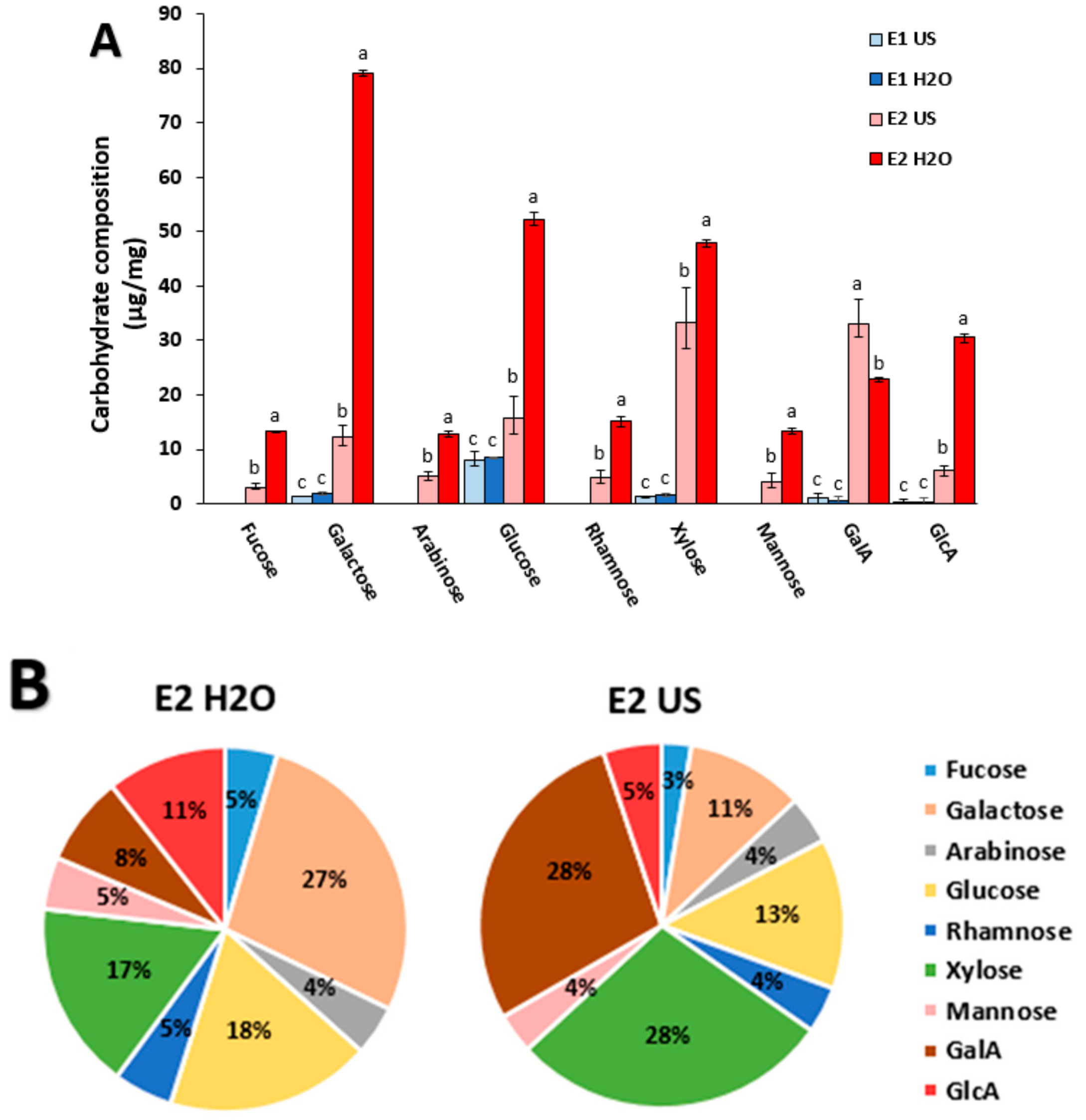
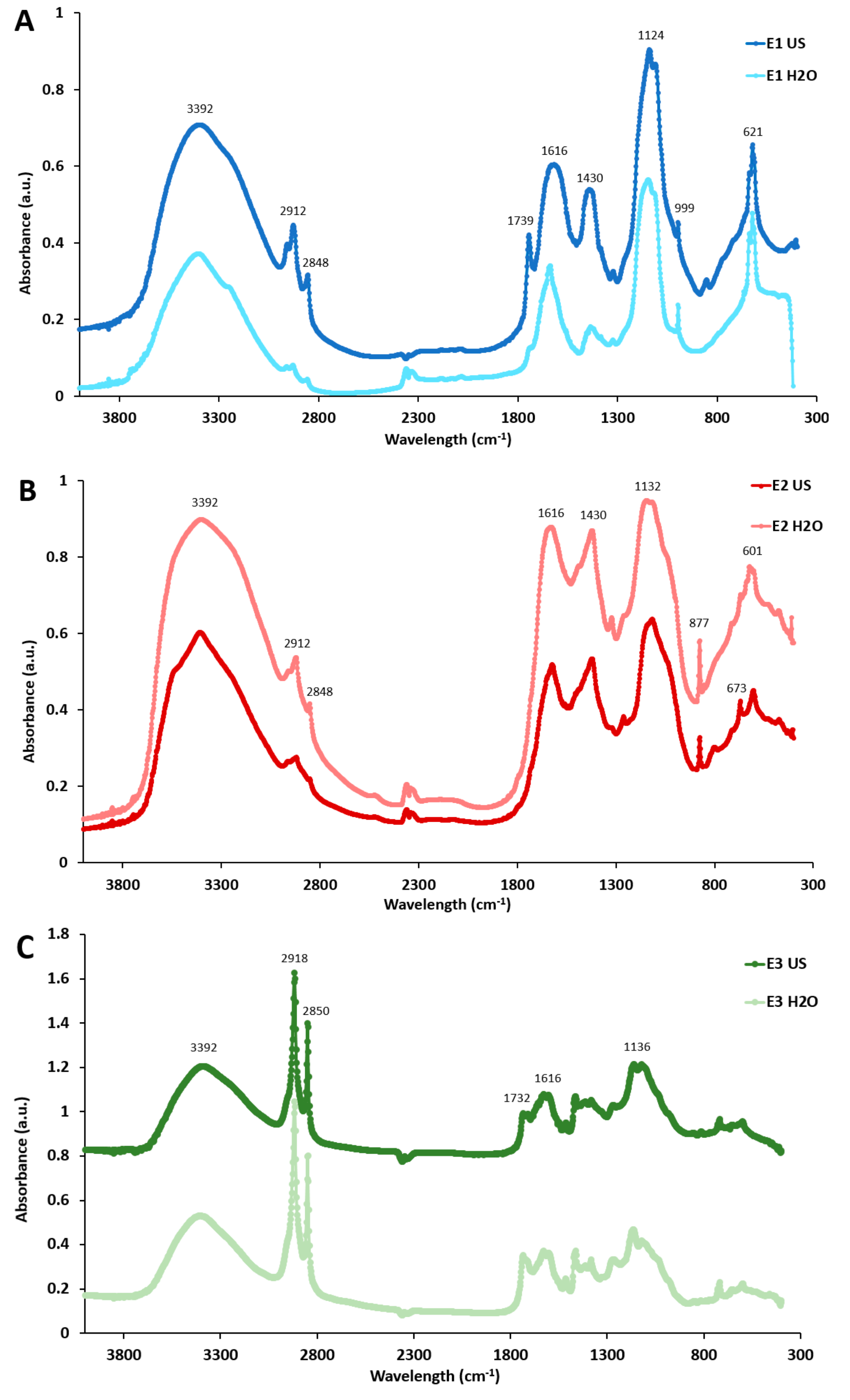
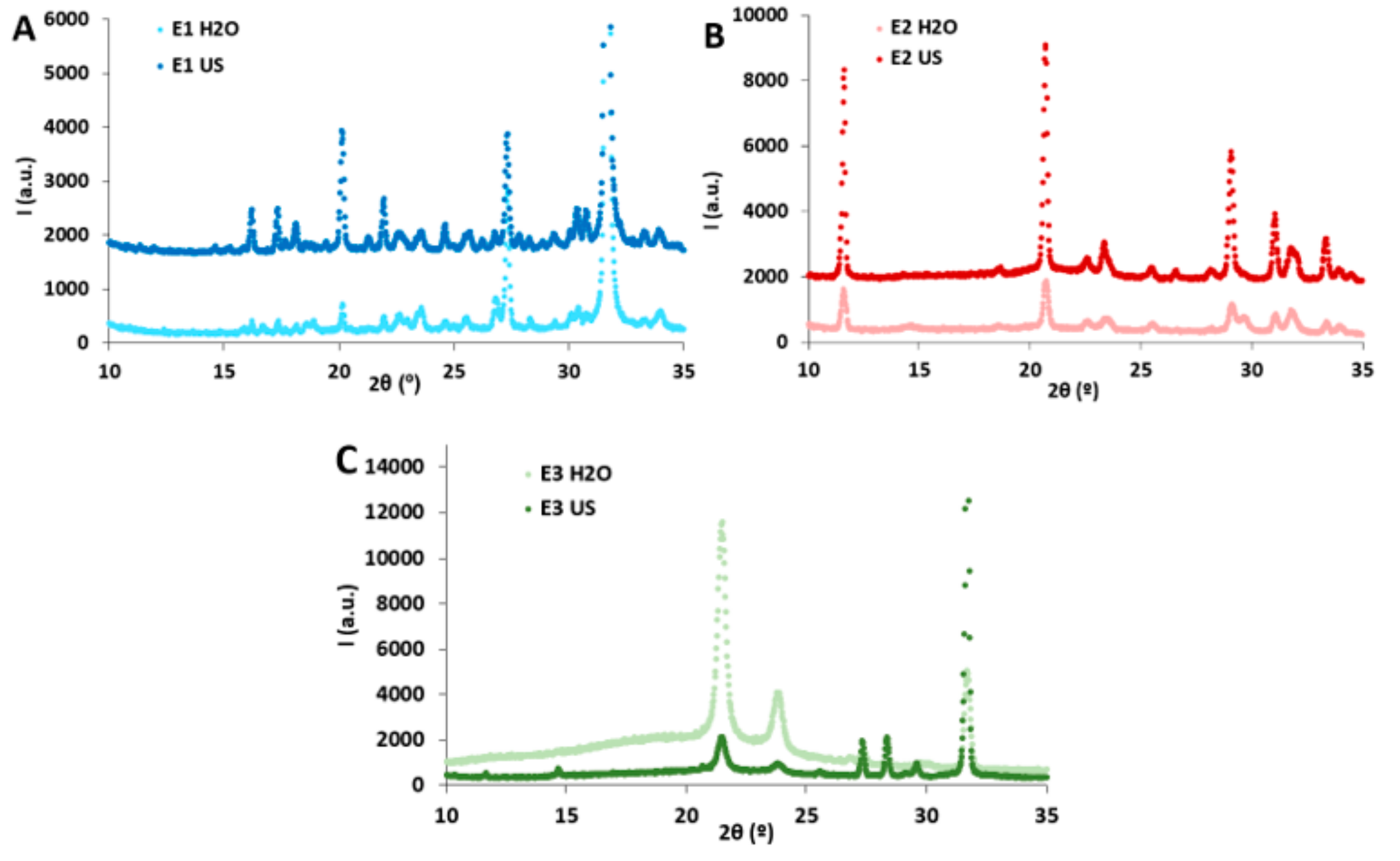
2.1. Composition and Structural Characterization of the P. oceanica Extracts
2.2. Evaluation of the Bioactive Properties from the P. oceanica Extracts
2.2.1. Antioxidant Capacity
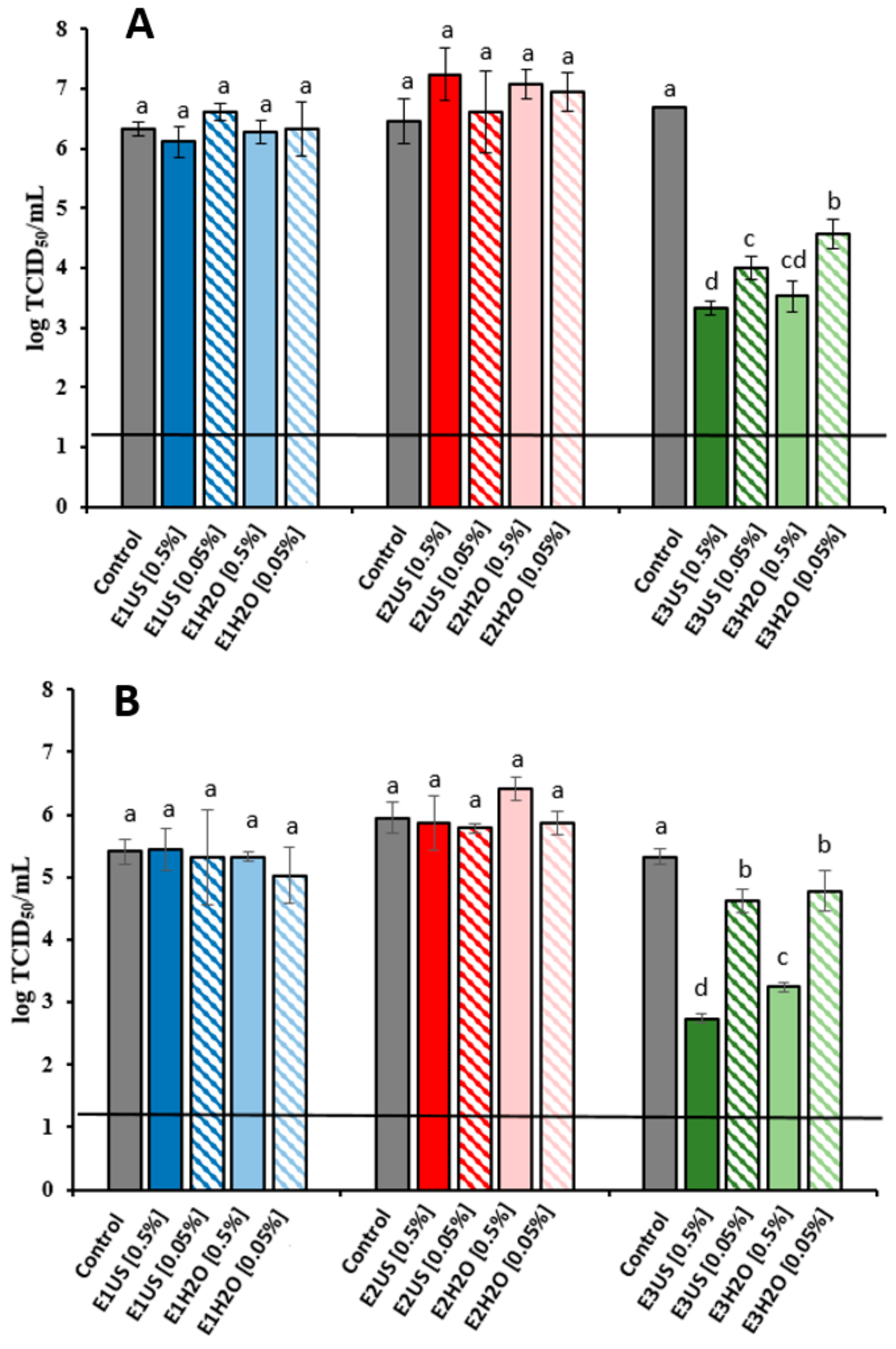
2.2.2. Antifungal Activity Assays
2.2.3. Antiviral Activity Assays
3. Materials and Methods
3.1. Materials and Reagents
3.2. Production of P. oceanica Extracts
3.2.1. Water-Based Extractions
3.2.2. Organic Solvent-Based Extractions
3.3. Chemical Composition Analysis
3.3.1. Total Phenolic Content
3.3.2. Protein Content
3.3.3. Lipid Content
3.3.4. Ash Content
3.4. Carbohydrate Composition
3.5. Fourier-Transform Infrared Spectroscopy (FT-IR)
3.6. X-ray Diffraction (XRD)
3.7. Thermogravimetric Analyses (TGA)
3.8. ABTS·+ Radical Cation Scavenging Activity
3.9. β-carotene–Linoleic Acid Assay
3.10. Antifungal Activity Assays
3.11. Antiviral Activity Assays
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lorenzo, J.; Agregán, R.; Munekata, P.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Munekata, P.E.; Gomez, B.; Barba, F.J.; Mora, L.; Perez-Santaescolastica, C.; Toldra, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Benito-González, I.; López-Rubio, A.; Martínez-Sanz, M. Potential of lignocellulosic fractions from Posidonia oceanica to improve barrier and mechanical properties of bio-based packaging materials. Int. J. Biol. Macromol. 2018, 118, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sanz, M.; Erboz, E.; Fontes, C.; López-Rubio, A. Valorization of Arundo donax for the production of high performance lignocellulosic films. Carbohydr. Polym. 2018, 199, 276–285. [Google Scholar] [CrossRef]
- Benito-González, I.; López-Rubio, A.; Martínez-Sanz, M. High-performance starch biocomposites with celullose from waste biomass: Film properties and retrogradation behaviour. Carbohydr. Polym. 2019, 216, 180–188. [Google Scholar] [CrossRef]
- Aguilar-Briseño, J.A.; Cruz-Suarez, L.E.; Sassi, J.-F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated polysaccharides from Ulva clathrata and Cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion, in NDV infection. Mar. Drugs 2015, 13, 697–712. [Google Scholar] [CrossRef]
- Piva, G.; Fracassetti, D.; Tirelli, A.; Mascheroni, E.; Musatti, A.; Inglese, P.; Piergiovanni, L.; Rollini, M. Evaluation of the antioxidant/antimicrobial performance of Posidonia oceanica in comparison with three commercial natural extracts and as a treatment on fresh-cut peaches (Prunus persica Batsch). Postharvest Biol. Technol. 2017, 124, 54–61. [Google Scholar] [CrossRef]
- Gokce, G.; Haznedaroglu, M.Z. Evaluation of antidiabetic, antioxidant and vasoprotective effects of Posidonia oceanica extract. J. Ethnopharmacol. 2008, 115, 122–130. [Google Scholar] [CrossRef]
- Kim, I.-S.; Yang, M.; Lee, O.-H.; Kang, S.-N. The antioxidant activity and the bioactive compound content of Stevia rebaudiana water extracts. Food Sci. Technol. 2011, 44, 1328–1332. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.; Escobedo-Avellaneda, Z.; Iqbal, H.; Welti-Chanes, J. State-of-the-art extraction methodologies for bioactive compounds from algal biome to meet bio-economy challenges and opportunities. Molecules 2018, 23, 2953. [Google Scholar] [CrossRef] [PubMed]
- Cikoš, A.-M.; Jokić, S.; Šubarić, D.; Jerković, I. Overview on the application of modern methods for the extraction of bioactive compounds from marine macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, Y.; Abdelhamid, A.; Mkadmini Hammi, K.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Microwave-assisted extraction and pharmacological evaluation of polysaccharides from Posidonia oceanica. Biosci. Biotechnol. Biochem. 2017, 81, 1917–1925. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yin, C.; Fan, X.; Fan, Z.; Shi, D.; Gao, H. Optimization of enzymes-microwave-ultrasound assisted extraction of Lentinus edodes polysaccharides and determination of its antioxidant activity. Int. J. Biol. Macromol. 2018, 111, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Surveswaran, S.; Cai, Y.-Z.; Corke, H.; Sun, M. Systematic evaluation of natural phenolic antioxidants from 133 Indian medicinal plants. Food Chem. 2007, 102, 938–953. [Google Scholar] [CrossRef]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Machu, L.; Misurcova, L.; Vavra Ambrozova, J.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Gómez-Ordóñez, E.; Rupérez, P. Infrared characterisation, monosaccharide profile and antioxidant activity of chemical fractionated polysaccharides from the edible seaweed sugar Kombu (Saccharina latissima). Int. J. Food Sci. Technol. 2015, 50, 340–346. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Kesraoui, O.; Marzouki, M.N.; Maugard, T.; Limam, F. In vitro evaluation of antioxidant activities of free and bound phenolic compounds from Posidonia oceanica (l.) Delile leaves. Afri. J. Biotechnol. 2011, 10, 3176. [Google Scholar]
- Martínez-Sanz, M.; Gómez-Mascaraque, L.G.; Ballester, A.R.; Martínez-Abad, A.; Brodkorb, A.; López-Rubio, A. Production of unpurified agar-based extracts from red seaweed Gelidium sesquipedale by means of simplified extraction protocols. Algal Res. 2019, 38, 101420. [Google Scholar] [CrossRef]
- Chu, Y.-F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef] [PubMed]
- Grignon-Dubois, M.; Rezzonico, B. Phenolic fingerprint of the seagrass Posidonia oceanica from four locations in the Mediterranean Sea: First evidence for the large predominance of chicoric acid. Bot. Mar. 2015, 58, 379–391. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Martínez-Abad, A.; Tan, H.-T.; Bulone, V.; Vilaplana, F. Sequential fractionation of feruloylated hemicelluloses and oligosaccharides from wheat bran using subcritical water and xylanolytic enzymes. Green Chem. 2017, 19, 1919–1931. [Google Scholar] [CrossRef]
- Martínez-Abad, A.; Giummarella, N.; Lawoko, M.; Vilaplana, F. Differences in extractability under subcritical water reveal interconnected hemicellulose and lignin recalcitrance in birch hardwoods. Green Chem. 2018, 20, 2534–2546. [Google Scholar] [CrossRef]
- Kaal, J.; Serrano, O.; José, C.; Rencoret, J. Radically different lignin composition in Posidonia species may link to differences in organic carbon sequestration capacity. Org. Geochem. 2018, 124, 247–256. [Google Scholar] [CrossRef]
- Pena, M.J.; Kulkarni, A.R.; Backe, J.; Boyd, M.; O’Neill, M.A.; York, W.S. Structural diversity of xylans in the cell walls of monocots. Planta 2016, 244, 589–606. [Google Scholar] [CrossRef]
- Izquierdo, M.; Marzal, P.; Gabaldón, C.; Silvetti, M.; Castaldi, P. Study of the interaction mechanism in the biosorption of copper (ii) ions onto Posidonia oceanica and peat. CLEAN 2012, 40, 428–437. [Google Scholar] [CrossRef]
- Dlapa, P.; Bodí, M.B.; Mataix-Solera, J.; Cerdà, A.; Doerr, S.H. FT-IR spectroscopy reveals that ash water repellency is highly dependent on ash chemical composition. Catena 2013, 108, 35–43. [Google Scholar] [CrossRef]
- Dick, D.P.; Knicker, H.; Ávila, L.G.; Inda Jr, A.V.; Giasson, E.; Bissani, C.A. Organic matter in constructed soils from a coal mining area in southern Brazil. Org. Geochem. 2006, 37, 1537–1545. [Google Scholar] [CrossRef]
- Shaltout, A.A.; Allam, M.A.; Moharram, M.A. FTIR spectroscopic, thermal and XRD characterization of hydroxyapatite from new natural sources. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2011, 83, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, M.; Qin, C.; Lv, B.; Mao, Q.; Liu, Z. Structural characterization and evaluation of the antioxidant activities of polysaccharides extracted from Qingzhuan brick tea. Int. J. Biol. Macromol. 2017, 101, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Posidonia oceanica (L.) Delile ethanolic extract modulates cell activities with skin health applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Jiang, X.; Han, X.; Wang, H. Fusion characteristic study on seaweed biomass ash. Energy Fuels 2008, 22, 2229–2235. [Google Scholar] [CrossRef]
- Nanda, S.; Mohanty, P.; Pant, K.K.; Naik, S.; Kozinski, J.A.; Dalai, A.K. Characterization of North American lignocellulosic biomass and biochars in terms of their candidacy for alternate renewable fuels. BioEnergy Res. 2013, 6, 663–677. [Google Scholar] [CrossRef]
- Vassilev, S.V.; Braekman-Danheux, C.; Laurent, P. Characterization of refuse-derived char from municipal solid waste: 1. Phase-mineral and chemical composition. Fuel Process. Technol. 1999, 59, 95–134. [Google Scholar] [CrossRef]
- Lutz, R.; Aserin, A.; Wicker, L.; Garti, N. Structure and physical properties of pectins with block-wise distribution of carboxylic acid groups. Food Hydrocol. 2009, 23, 786–794. [Google Scholar] [CrossRef]
- Carà, P.D.; Pagliaro, M.; Elmekawy, A.; Brown, D.R.; Verschuren, P.; Shiju, N.R.; Rothenberg, G. Hemicellulose hydrolysis catalysed by solid acids. Catal. Sci. Technol. 2013, 3, 2057–2061. [Google Scholar] [CrossRef]
- Ueno, S.; Miyazaki, A.; Yano, J.; Furukawa, Y.; Suzuki, M.; Sato, K. Polymorphism of linoleic acid (cis-9, cis-12-Octadecadienoic acid) and α-linolenic acid (cis-9, cis-12, cis-15-Octadecatrienoic acid). Chem. Phys. Lipids 2000, 107, 169–178. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Akbar, J.; Hussain, M.A.; Saghir, S.; Sher, M. Evaluation of hot-water extracted arabinoxylans from ispaghula seeds as drug carriers. Carbohydr. Polym. 2011, 83, 1218–1225. [Google Scholar] [CrossRef]
- Aleksandrova, G.; Lesnichaya, M.; Myachin, Y.A.; Sukhov, B.; Trofimov, B. Effect of silver nanoparticles on the thermal characteristics of nanocomposites of galactose-containing polysaccharides. Dokl. Chem. 2011, 439, 187. [Google Scholar] [CrossRef]
- Jayadas, N.; Nair, K.P. Coconut oil as base oil for industrial lubricants—evaluation and modification of thermal, oxidative and low temperature properties. Tribol. Int. 2006, 39, 873–878. [Google Scholar] [CrossRef]
- De Oliveira, J.P.; Bruni, G.P.; Fabra, M.J.; Da Rosa Zavareze, E.; López-Rubio, A.; Martínez-Sanz, M. Development of food packaging bioactive aerogels through the valorization of Gelidium sesquipedale seaweed. Food Hydrocoll. 2019, 89, 337–350. [Google Scholar] [CrossRef]
- Tupec, M.; Hýsková, V.; Bělonožníková, K.; Hraníček, J.; Červený, V.; Ryšlavá, H. Characterization of some potential medicinal plants from Central Europe by their antioxidant capacity and the presence of metal elements. Food Biosci. 2017, 20, 43–50. [Google Scholar] [CrossRef]
- Hamid, H.A.; Mutazah, R.; Yusoff, M.M.; Karim, N.A.A.; Razis, A.F.A. Comparative analysis of antioxidant and antiproliferative activities of Rhodomyrtus tomentosa extracts prepared with various solvents. Food Chem. Toxicol. 2016, 108, 451–457. [Google Scholar] [CrossRef]
- Jeddou, K.B.; Chaari, F.; Maktouf, S.; Nouri-Ellouz, O.; Helbert, C.B.; Ghorbel, R.E. Structural, functional, and antioxidant properties of water-soluble polysaccharides from potatoes peels. Food Chem. 2016, 205, 97–105. [Google Scholar] [CrossRef]
- Bouaziz, F.; Koubaa, M.; Ghorbel, R.E.; Chaabouni, S.E. Biological properties of water-soluble polysaccharides and hemicelluloses from almond gum. Int. J. Biol. Macromol. 2017, 95, 667–674. [Google Scholar] [CrossRef]
- Wilson, C.; Solar, J.; El Ghaouth, A.; Wisniewski, M. Rapid evaluation of plant extracts and essential oils for antifungal activity against Botrytis cinerea. Plant Dis. 1997, 81, 204–210. [Google Scholar] [CrossRef]
- Afolayan, A. Extracts from the shoots of Arctotis arctotoides inhibit the growth of bacteria and fungi. Pharm. Biol. 2003, 41, 22–25. [Google Scholar] [CrossRef]
- Randazzo, W.; Fabra, M.J.; Falcó, I.; López-Rubio, A.; Sánchez, G. Polymers and biopolymers with antiviral activity: Potential applications for improving food safety. Compr. Rev. Food Sci Food Saf. 2018, 17, 754–768. [Google Scholar] [CrossRef]
- Oh, E.-G.; Kim, K.-L.; Shin, S.B.; Son, K.-T.; Lee, H.-J.; Kim, T.H.; Kim, Y.-M.; Cho, E.-J.; Kim, D.-K.; Lee, E.-W. Antiviral activity of green tea catechins against feline calicivirus as a surrogate for norovirus. Food Sci. Biotechnol. 2013, 22, 593–598. [Google Scholar] [CrossRef]
- Su, X.; D’Souza, D.H. Grape seed extract for the control of human enteric viruses. App. Environ. Microbiol. 2011, 77, 3982–3987. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Frings, C.S.; Dunn, R.T. A colorimetric method for determination of total serum lipids based on the sulfo-phospho-vanillin reaction. Am. J. Clin. Pathol. 1970, 53, 89–91. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Miller, H. A simplified method for the evaluation of antioxidants. J. Am. Oil Chem. Soc. 1971, 48, 91. [Google Scholar] [CrossRef]
- Falcó, I.; Randazzo, W.; Gómez-Mascaraque, L.G.; Aznar, R.; López-Rubio, A.; Sánchez, G. Fostering the antiviral activity of green tea extract for sanitizing purposes through controlled storage conditions. Food Control 2018, 84, 485–492. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Carbohydrate Content (mg/g Extract) | Protein Content (mg BSA/g Extract) | Lipid Content (mg/g Extract) | Total Phenolics (mg GAE/g Extract) | Ash Content (mg/g Extract) | TEAC (µmol TE/g Extract) * | % β-Carotene Bleaching Inhibition † |
|---|---|---|---|---|---|---|---|
| E1 US | 12.2 ± 2.9 c | 58.4 ± 3.2 d | n.d. | 7.6 ± 2.2 c | 751.5 ± 1.4 a | 275.9 ± 10.2 c,d | 57.0 ± 2.3 c |
| E1 H2O | 12.9 ± 2.4 c | 70.9 ± 1.4 c | n.d. | 37.9 ± 8.0 b | 723.1 ± 6.6 b | 236.5 ± 16.5 e | 45.0 ± 1.8 d |
| E2 US | 117.7 ± 18.5 b | 269.3 ± 5.5 b | n.d. | 48.7 ± 5.5 b | 413.3 ± 8.8 c | 346.7 ± 5.6 b | 68.7 ± 4.2 b |
| E2 H2O | 287.1 ± 2.5 a | 363.0 ± 1.9 a | n.d. | 90.2 ± 17.6 a | 339.1 ± 9.5 d | 730.0 ± 12.2 a | 65.5 ± 5.3 b |
| E3 US | - | - | 785.3 ± 73.5 a | 79.6 ± 23.1 a | 180.7 ± 3.6 e | 211.2 ± 51.2 d,e | 70.8 ± 2.6 b |
| E3 H2O | - | - | 560.3 ± 85.3 b | 76.2 ± 3.3 a | 347.5 ± 22.9 d | 443.8 ± 150.2 b,c | 82.8 ± 3.5 a |
| Microorganism | E1 US | E1 H2O | E2 US | E2 H2O | E3 US | E3 H2O |
| 3 dpi. | ||||||
| Penicillium digitatum | - | - | 37 ± 9 | 12 ± 3 | 46 ± 2 | 14 ± 4 |
| Penicillium italicum | - | - | - | - | 40 ± 2 | 24 ± 2 |
| Penicillium expansum | - | - | - | - | −(42 ± 3) a | −(27 ± 3) a |
| Botrytis cinerea | - | - | 41 ± 12 | 19 ± 8 | 100 | 100 |
| Geotrichum candidum | - | - | - | - | - | - |
| Aspergillus niger | - | - | - | - | −(100) a | −(90 ± 4) a |
| 6–7 dpi | ||||||
| Penicillium digitatum | - | - | 24 ± 8 | 12 ± 4 | - | - |
| Penicillium italicum | - | - | - | - | 15 ± 3 b | − b |
| Penicillium expansum | - | - | - | - | −(22 ± 3) a | −(19 ± 2) a |
| Botrytis cinerea | - | - | - | - | 61 ± 4 | 69 ± 5 |
| Geotrichum candidum | - | - | - | - | - | - |
| Aspergillus niger | - | - | - | - | −(100) a | −(70 ± 11) a |
| Sample Code | Pre-Treatment | Extraction Method | Solvent | Fraction (Ethanol Precipitation) |
|---|---|---|---|---|
| E1 US | - | Ultrasound | Water | Supernatant |
| E1 H2O | - | Heating | Water | Supernatant |
| E2 US | - | Ultrasound | Water | Precipitate |
| E2 H2O | - | Heating | Water | Precipitate |
| E3 US | Ultrasound | Soxhlet | Toluene:ethanol | - |
| E3 H2O | Heating | Soxhlet | Toluene:ethanol | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito-González, I.; López-Rubio, A.; Martínez-Abad, A.; Ballester, A.-R.; Falcó, I.; González-Candelas, L.; Sánchez, G.; Lozano-Sánchez, J.; Borrás-Linares, I.; Segura-Carretero, A.; et al. In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass. Mar. Drugs 2019, 17, 409. https://doi.org/10.3390/md17070409
Benito-González I, López-Rubio A, Martínez-Abad A, Ballester A-R, Falcó I, González-Candelas L, Sánchez G, Lozano-Sánchez J, Borrás-Linares I, Segura-Carretero A, et al. In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass. Marine Drugs. 2019; 17(7):409. https://doi.org/10.3390/md17070409
Chicago/Turabian StyleBenito-González, Isaac, Amparo López-Rubio, Antonio Martínez-Abad, Ana-Rosa Ballester, Irene Falcó, Luis González-Candelas, Gloria Sánchez, Jesús Lozano-Sánchez, Isabel Borrás-Linares, Antonio Segura-Carretero, and et al. 2019. "In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass" Marine Drugs 17, no. 7: 409. https://doi.org/10.3390/md17070409
APA StyleBenito-González, I., López-Rubio, A., Martínez-Abad, A., Ballester, A.-R., Falcó, I., González-Candelas, L., Sánchez, G., Lozano-Sánchez, J., Borrás-Linares, I., Segura-Carretero, A., & Martínez-Sanz, M. (2019). In-Depth Characterization of Bioactive Extracts from Posidonia oceanica Waste Biomass. Marine Drugs, 17(7), 409. https://doi.org/10.3390/md17070409