Characterizing Eckol as a Therapeutic Aid: A Systematic Review



Abstract
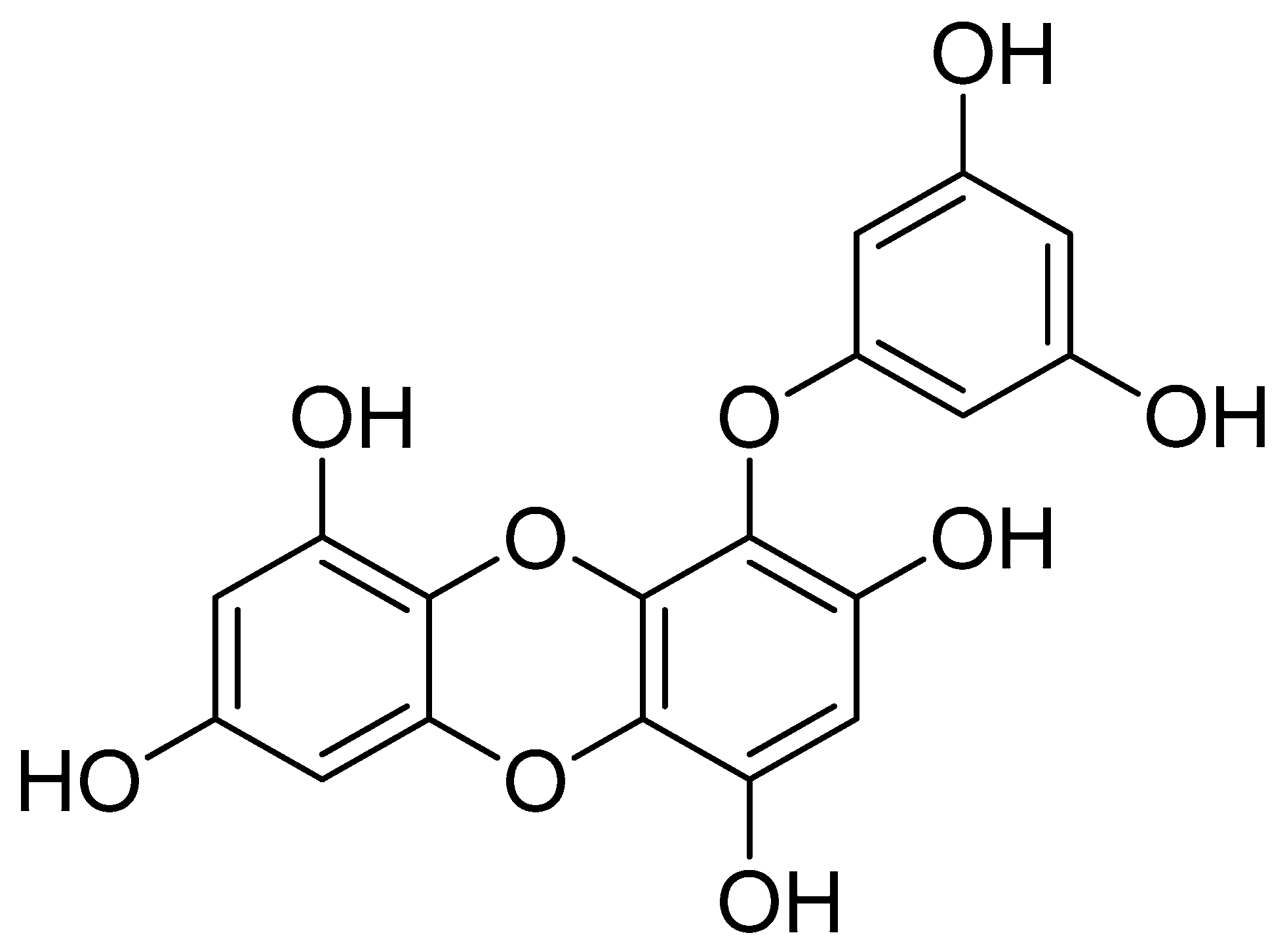
1. Introduction
2. Biological Activities of Eckol
2.1. Anti-Oxidant Activity
2.2. Anti-Diabetic Activity
2.3. Hepatoprotective Activity
2.4. Neuroprotective Activity
2.5. Radioprotective Activity
2.6. Anti-Photo Aging Effects
2.7. Anti-Hypertensive Activity
2.8. Anticoagulative Activity
2.9. Antibacterial and Antiviral Activity
2.10. Anti-Obesity Activity
2.11. Anticancer and Cytotoxic Activity
2.12. Anti-Histaminic Activity
2.13. Anti-Inflammatory Activity
2.14. Anti-Hyperlipidemic Activity
2.15. Anti-Tyrosinase Activity
2.16. Miscellaneous
3. Discussion
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Muhammad, S.A.; Muhammad, J.; Muhammad, S.; Muhammad, K.P.; Shaista, H.; Viqar, U.A. Metabolites of marine algae collected from Karachi-coasts of Arabian Sea. Nat. Prod. Sci. 2000, 6, 61–65. [Google Scholar]
- Chen, J.H.; Lim, J.D.; Sohn, E.H.; Choi, Y.S.; Han, E.T. Growth-inhibitory effect of a fucoidan from brown seaweed Undaria pinnatifida on Plasmodium parasites. Parasitol. Res. 2009, 104, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Taskin, E.; Ozturk, M.; Kurt, O. Antibacterial activities of some marine algae from the Aegean Sea (Turkey). Afr. J. Biotechnol. 2007, 6, 2746–2751. [Google Scholar]
- Dhargalkar, V.; Pereira, N. Seaweed: Promising plant of the millennium. Sci. Cult. 2005, 71, 60–66. [Google Scholar]
- Zou, Y.; Qian, Z.J.; Li, Y.; Kim, M.M.; Lee, S.H.; Kim, S.K. Antioxidant effects of phlorotannins isolated from Ishige okamurae in free radical mediated oxidative systems. J. Agric. Food Chem. 2008, 56, 7001–7009. [Google Scholar] [CrossRef]
- Airanthi, M.W.A.; Hosokawa, M.; Miyashita, K. Comparative antioxidant activity of edible Japanese brown seaweeds. J. Food Sci. 2011, 76, C104–C111. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef]
- Singh, I.P.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Kang, S.M.; Cha, S.H.; Ko, J.Y.; Kang, M.C.; Kim, D.; Heo, S.J.; Kim, J.S.; Heu, M.S.; Kim, Y.T.; Jung, W.K.; et al. Neuroprotective effects of phlorotannins isolated from a brown alga, Ecklonia cava, against H2O2-induced oxidative stress in murine hippocampal HT22 cells. Environ. Toxicol. Pharmacol. 2012, 34, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Seok, J.K.; Boo, Y.C. Ecklonia cava extract and dieckol attenuate cellular lipid peroxidation in keratinocytes exposed to PM10. Evid. Based Complement. Alternat. Med. 2018, 2018, 8248323. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.S.; Chung, H.Y.; Kim, J.Y.; Son, B.W.; Jung, H.A.; Choi, J.S. Inhibitory phlorotannins from the edible brown alga Ecklonia stolonifera on total reactive oxygen species (ROS) generation. Arch. Pharm. Res. 2004, 27, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Shin, T.; Utsuki, T.; Choi, J.S.; Byun, D.S.; Kim, H.R. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and hepatoprotective properties in tacrine-treated HepG2 cells. J. Agric. Food Chem. 2012, 60, 5340–5349. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Kitajima, C.; Ito, H.; Miyazaki, T.; Baba, M.; Okuyama, T.; Okada, Y. Antidiabetic effect of polyphenols from brown alga Ecklonia kurome in genetically diabetic KK-Ay mice. Pharm. Biol. 2012, 50, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Takahashi, M.; Fukuyama, Y.; Kinzyo, Z. An anti-plasmin inhibitor, eckol, isolated from the brown alga Ecklonia kurome Okamura. Agric. Biol. Chem. 1989, 53, 3025–3030. [Google Scholar]
- Rengasamy, K.R.; Aderogba, M.A.; Amoo, S.O.; Stirk, W.A.; Van Staden, J. Potential antiradical and alpha-glucosidase inhibitors from Ecklonia maxima (Osbeck) Papenfuss. Food Chem. 2013, 141, 1412–1415. [Google Scholar] [CrossRef]
- Nakamura, T.; Nagayama, K.; Uchida, K.; Tanaka, R. Antioxidant activity of phlorotannins isolated from the brown alga Eisenia bicyclis. Fish. Sci. 1996, 62, 923–926. [Google Scholar] [CrossRef]
- Sugiura, Y.; Matsuda, K.; Yamada, Y.; Nishikawa, M.; Shioya, K.; Katsuzaki, H.; Imai, K.; Amano, H. Anti-allergic phlorotannins from the edible brown alga, Eisenia arborea. Food Sci. Technol. Res. 2007, 13, 54–60. [Google Scholar] [CrossRef]
- Joung, E.J.; Lee, M.S.; Choi, J.W.; Kim, J.S.; Shin, T.; Jung, B.M.; Yoon, N.Y.; Lim, C.W.; Kim, J.I.; Kim, H.R. Anti-inflammatory effect of ethanolic extract from Myagropsis myagroides on murine macrophages and mouse ear edema. BMC Complement. Altern. Med. 2012, 12, 171. [Google Scholar] [CrossRef]
- Kang, M.C.; Cha, S.H.; Wijesinghe, W.; Kang, S.M.; Lee, S.H.; Kim, E.A.; Song, C.B.; Jeon, Y.J. Protective effect of marine algae phlorotannins against AAPH-induced oxidative stress in zebrafish embryo. Food Chem. 2013, 138, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.J.; Lee, M.; Shin, T.; Yoon, N.; Kim, J.H.; Kim, H.R. Eckol enhances heme oxygenase-1 expression through activation of Nrf2/JNK pathway in HepG2 cells. Molecules 2014, 19, 15638–15652. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Tanaka, R.; Katsuzaki, H.; Imai, K.; Matsushita, T. The anti-inflammatory effects of phlorotannins from Eisenia arborea on mouse ear edema by inflammatory inducers. J. Funct. Foods 2013, 5, 2019–2023. [Google Scholar] [CrossRef]
- Kim, T.H.; Ku, S.K.; Lee, T.; Bae, J.S. Vascular barrier protective effects of phlorotannins on HMGB1-mediated proinflammatory responses in vitro and in vivo. Food Chem. Toxicol. 2012, 50, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Kim, J.I.; Choung, S.Y.; Choi, J.S. Protective effect of the edible brown alga Ecklonia stolonifera on doxorubicin-induced hepatotoxicity in primary rat hepatocytes. J. Pharm. Pharmacol. 2014, 66, 1180–1188. [Google Scholar]
- Kannan, R.R.; Aderogba, M.A.; Ndhlala, A.R.; Stirk, W.A.; Van Staden, J. Acetylcholinesterase inhibitory activity of phlorotannins isolated from the brown alga, Ecklonia maxima (Osbeck) Papenfuss. Food Res. Int. 2013, 54, 1250–1254. [Google Scholar] [CrossRef]
- Kim, I.H.; Nam, T.J. Enzyme-treated Ecklonia cava extract inhibits adipogenesis through the downregulation of C/EBPα in 3T3-L1 adipocytes. Int. J. Mol. Med. 2017, 39, 636–644. [Google Scholar] [CrossRef]
- Jung, H.A.; Hyun, S.K.; Kim, H.R.; Choi, J.S. Angiotensin-converting enzyme I inhibitory activity of phlorotannins from Ecklonia stolonifera. Fish. Sci. 2006, 72, 1292–1299. [Google Scholar] [CrossRef]
- Choi, J.G.; Kang, O.H.; Brice, O.O.; Lee, Y.S.; Chae, H.S.; Oh, Y.C.; Sohn, D.H.; Park, H.; Choi, H.G.; Kim, S.G. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods. 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Shanmugam, M.; Mody, K. Heparinoid-active sulphated polysaccharides from marine algae as potential blood anticoagulant agents. Curr. Sci. 2000, 79, 1672–1683. [Google Scholar]
- Abdul, Q.A.; Choi, R.J.; Jung, H.A.; Choi, J.S. Health benefit of fucosterol from marine algae: A review. J. Sci. Food Agric. 2016, 96, 1856–1866. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Koirala, P.; Jung, H.A.; Choi, J.S. Recent advances in pharmacological research on Ecklonia species: A review. Arch. Pharm. Res. 2017, 40, 981–1005. [Google Scholar] [CrossRef]
- Yu, K.X.; Jantan, I.; Ahmad, R.; Wong, C.L. The major bioactive components of seaweeds and their mosquitocidal potential. Parasitol. Res. 2014, 113, 3121–3141. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Abu-Ghannam, N. Recent developments in the application of seaweeds or seaweed extracts as a means for enhancing the safety and quality attributes of foods. Innov. Food Sci. Emerg. Technol. 2011, 12, 600–609. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Jeon, Y.J. Biological activities and potential cosmeceutical applications of bioactive components from brown seaweeds: A review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Jeon, Y.J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.K. Phlorotannins from Ecklonia cava (Phaeophyceae): Biological activities and potential health benefits. Biofactors 2010, 36, 408–414. [Google Scholar] [CrossRef]
- Shin, T.; Ahn, M.; Hyun, J.W.; Kim, S.H.; Moon, C. Antioxidant marine algae phlorotannins and radioprotection: A review of experimental evidence. Acta Histochem. 2014, 116, 669–674. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Z.J.; Ryu, B.; Lee, S.H.; Kim, M.M.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorg. Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.C.; Kang, K.A.; Zhang, R.; Piao, M.J.; Kim, G.Y.; Kang, M.Y.; Lee, S.J.; Lee, N.H.; Surh, Y.J.; Hyun, J.W. Up-regulation of Nrf2-mediated heme oxygenase-1 expression by eckol, a phlorotannin compound, through activation of Erk and PI3K/Akt. Int. J. Biochem. Cell Biol. 2010, 42, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Lee, Y.; Kim, S.Y.; Kim, H.S.; Joo, H.G.; Park, J.W.; et al. Eckol isolated from Ecklonia cava attenuates oxidative stress induced cell damage in lung fibroblast cells. FEBS Lett. 2005, 579, 6295–6304. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.D.; Kang, K.A.; Piao, M.J.; Kim, K.C.; Zheng, J.; Yao, C.W.; Cha, J.W.; Hyun, C.L.; Kang, H.K.; Lee, N.H.; et al. Cytoprotective effect of Eckol against oxidative stress-induced mitochondrial dysfunction: Involvement of the FoxO3a/AMPK pathway. J. Cell. Biochem. 2014, 115, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Tanaka, R.; Miyake, H.; Tamaru, Y.; Ueda, M.; Shibata, T. Evaluation for antioxidative properties of phlorotannins isolated from the brown alga Eisenia bicyclis, by the H-ORAC method. Food Nutr. Sci. 2013, 4, 78. [Google Scholar]
- Jung, H.A.; Yoon, N.Y.; Woo, M.H.; Choi, J.S. Inhibitory activities of extracts from several kinds of seaweeds and phlorotannins from the brown alga Ecklonia stolonifera on glucose-mediated protein damage and rat lens aldose reductase. Fish. Sci. 2008, 74, 1363–1365. [Google Scholar] [CrossRef]
- Kim, Y.C.; An, R.B.; Yoon, N.Y.; Nam, T.J.; Choi, J.S. Hepatoprotective constituents of the edible brown alga Ecklonia stolonifera on tacrine-induced cytotoxicity in HepG2 cells. Arch. Pharm. Res. 2005, 28, 1376. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Zhang, M.; Chen, Y.; Zhu, T.; Wang, J. Protective effect of eckol against acute hepatic injury induced by carbon tetrachloride in mice. Mar. Drugs 2018, 16, 300. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, K.; Jeon, J.S.; Jho, E.H.; Kim, C.Y.; Nho, C.W.; Um, B.H. Isolation of phlorotannins from Eisenia bicyclis and their hepatoprotective effect against oxidative stress induced by tert-butyl hyperoxide. Appl. Biochem. Biotechnol. 2011, 165, 1296–1307. [Google Scholar] [CrossRef]
- Kang, M.C.; Kim, K.N.; Kang, S.M.; Yang, X.; Kim, E.A.; Song, C.B.; Nah, J.W.; Jang, M.K.; Lee, J.S.; Jung, W.K.; et al. Protective effect of dieckol isolated from Ecklonia cava against ethanol caused damage in vitro and in zebrafish model. Environ. Toxicol. Pharmacol. 2013, 36, 1217–1226. [Google Scholar] [CrossRef]
- Jung, H.A.; Oh, S.H.; Choi, J.S. Molecular docking studies of phlorotannins from Eisenia bicyclis with BACE1 inhibitory activity. Bioorg. Med. Chem. Lett. 2010, 20, 3211–3215. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.Y.; Chung, H.Y.; Kim, H.R.; Choi, J.S. Acetyl-and butyrylcholinesterase inhibitory activities of sterols and phlorotannins from Ecklonia stolonifera. Fish. Sci. 2008, 74, 200. [Google Scholar] [CrossRef]
- Jung, H.A.; Roy, A.; Jung, J.H.; Choi, J.S. Evaluation of the inhibitory effects of eckol and dieckol isolated from edible brown alga Eisenia bicyclis on human monoamine oxidases A and B. Arch. Pharm. Res. 2017, 40, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.R.; Moon, H.E.; Kim, H.R.; Jung, H.A.; Choi, J.S. Neuroprotective effect of edible brown alga Eisenia bicyclis on amyloid beta peptide-induced toxicity in PC12 cells. Arch. Pharm. Res. 2012, 35, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Yang, H.; Jeon, Y.J.; Lee, C.J.; Jin, Y.H.; Baek, N.I.; Kim, D.; Kang, S.M.; Yoon, M.; Yong, H.; et al. Phlorotannins of the edible brown seaweed Ecklonia cava Kjellman induce sleep via positive allosteric modulation of gamma-aminobutyric acid type A–benzodiazepine receptor: A novel neurological activity of seaweed polyphenols. Food Chem. 2012, 132, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Paudel, P.; Seong, S.H.; Wu, S.; Park, S.; Jung, H.A.; Choi, J.S. Eckol as a potential therapeutic against neurodegenerative diseases targeting dopamine D3/D4 receptors. Mar. Drugs 2019, 17, 108. [Google Scholar] [CrossRef] [PubMed]
- Joe, M.J.; Kim, S.N.; Choi, H.Y.; Shin, W.S.; Park, G.M.; Kang, D.W.; Kim, Y.K. The inhibitory effects of eckol and dieckol from Ecklonia stolonifera on the expression of matrix metalloproteinase-1 in human dermal fibroblasts. Biol. Pharm. Bull. 2006, 29, 1735–1739. [Google Scholar] [CrossRef]
- Piao, M.J.; Lee, N.H.; Chae, S.; Hyun, J.W. Eckol inhibits ultraviolet B-induced cell damage in human keratinocytes via a decrease in oxidative stress. Biol. Pharm. Bull. 2012, 35, 873–880. [Google Scholar] [CrossRef][Green Version]
- Hwang, H.; Chen, T.; Nines, R.G.; Shin, H.C.; Stoner, G.D. Photochemoprevention of UVB-induced skin carcinogenesis in SKH-1 mice by brown algae polyphenols. Int. J. Cancer 2006, 119, 2742–2749. [Google Scholar] [CrossRef]
- Heo, S.J.; Ko, S.C.; Cha, S.H.; Kang, D.H.; Park, H.S.; Choi, Y.U.; Kim, D.; Jung, W.K.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on melanogenesis and their protective effect against photo-oxidative stress induced by UV-B radiation. Toxicol. In Vitro 2009, 23, 1123–1130. [Google Scholar] [CrossRef]
- Cha, S.H.; Ko, C.I.; Kim, D.; Jeon, Y.J. Protective effects of phlorotannins against ultraviolet B radiation in zebrafish (Danio rerio). Vet. Dermatol. 2012, 23, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Lee, N.H.; Joo, H.G.; Jee, Y. Modulation of apoptosis of eckol against ionizing radiation in mice. Biochem. Biophys. Res. Commun. 2008, 372, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.; Kim, S.H.; Kim, J.C.; Hyun, J.W.; Lee, N.H.; Park, J.W.; Shin, T. Protective effect of phlorotannin components phloroglucinol and eckol on radiation-induced intestinal injury in mice. Phytother. Res. 2008, 22, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Ahn, G.N.; Lee, N.H.; Kim, J.M.; Yun, J.S.; Hyun, J.W.; Jeon, Y.J.; Wie, M.B.; Lee, Y.J.; Park, J.W. Radioprotective properties of eckol against ionizing radiation in mice. FEBS Lett. 2008, 582, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Ko, D.O.; Wang, Z.H.; Lee, I.K.; Kim, B.J.; Jeong, I.Y.; Shin, T.; Park, J.W.; et al. Eckol protects V79-4 lung fibroblast cells against γ-ray radiation-induced apoptosis via the scavenging of reactive oxygen species and inhibiting of the c-Jun NH2-terminal kinase pathway. Eur. J. Pharmacol. 2008, 591, 114–123. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Yoon, S.Y.; Park, J.Y.; Kim, Y.M.; Park, S.J.; Rho, M.C.; Kim, S.J.; Lee, W.S. Influenza virus neuraminidase inhibitory activity of phlorotannins from the edible brown alga Ecklonia cava. J. Agric. Food Chem. 2011, 59, 6467–6473. [Google Scholar] [CrossRef]
- Yang, H.K.; Jung, M.H.; Avunje, S.; Nikapitiya, C.; Kang, S.Y.; Ryu, Y.B.; Lee, W.S.; Jung, S.J. Efficacy of algal Ecklonia cava extract against viral hemorrhagic septicemia virus (VHSV). Fish. Shellfish Immunol. 2018, 72, 273–281. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, J.H.; Kwon, J.M.; Kwon, H.J.; Jeong, H.J.; Kim, Y.M.; Kim, D.; Lee, W.S.; Ryu, Y.B. Dieckol, a SARS-CoV 3CL (pro) inhibitor, isolated from the edible brown algae Ecklonia cava. Bioorg. Med. Chem. 2013, 21, 3730–3737. [Google Scholar] [CrossRef]
- Kim, T.H.; Lee, T.; Ku, S.K.; Bae, J.S. Vascular barrier protective effects of eckol and its derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 3710–3712. [Google Scholar]
- Sugiura, Y.; Usui, M.; Katsuzaki, H.; Imai, K.; Kakinuma, M.; Amano, H.; Miyata, M. Orally administered phlorotannins from Eisenia arborea suppress chemical mediator release and cyclooxygenase-2 signaling to alleviate mouse ear swelling. Mar. Drugs 2018, 16, 267. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, E.H.; Park, K.; Kwon, J.Y.; Kim, P.H.; Jung, W.K.; Kim, Y.M. Eckol from Eisenia bicyclis inhibits inflammation through the Akt/NF-κB signaling in Propionibacterium acnes-induced human keratinocyte HaCaT cells. J. Food Biochem. 2017, 41, e12312. [Google Scholar] [CrossRef]
- Shibata, T.; Nagayama, K.; Tanaka, R.; Yamaguchi, K.; Nakamura, T. Inhibitory effects of brown algal phlorotannins on secretory phospholipase A2s, lipoxygenases and cyclooxygenases. J. Appl. Phycol. 2003, 15, 61–66. [Google Scholar] [CrossRef]
- Li, Y.; Lee, S.H.; Le, Q.T.; Kim, M.M.; Kim, S.K. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and FcεRI. J. Agric. Food Chem. 2008, 56, 12073–12080. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Fujimoto, K.; Nagayama, K.; Yamaguchi, K.; Nakamura, T. Inhibitory activity of brown algal phlorotannins against hyaluronidase. Int. J. Food Sci. Technol. 2002, 37, 703–709. [Google Scholar] [CrossRef]
- Hyun, K.H.; Yoon, C.H.; Kim, R.K.; Lim, E.J.; An, S.; Park, M.J.; Hyun, J.W.; Suh, Y.; Kim, M.J.; Lee, S.J. Eckol suppresses maintenance of stemness and malignancies in glioma stem-like cells. Toxicol. Appl. Pharmacol. 2011, 254, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.M.; Wilson, M.N.; Martin, O.O.; Ntevheleni, T.; Wilfred, T.M. Phlorotannins and a sterol isolated from a brown alga Ecklonia maxima, and their cytotoxic activity against selected cancer cell lines HeLa, H157 and MCF7. Interdiscip. J. Chem. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Kim, T.H.; Ku, S.K.; Bae, J.S. Antithrombotic and profibrinolytic activities of eckol and dieckol. J. Cell. Biochem. 2012, 113, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- Park, E.Y.; Kim, E.H.; Kim, M.H.; Seo, Y.W.; Lee, J.I.; Jun, H.S. Polyphenol-rich fraction of brown alga Ecklonia cava collected from Gijang, Korea, reduces obesity and glucose levels in high-fat diet-induced obese mice. Evid. Based Complement. Alternat. Med. 2012, 2012, 418912. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Lee, M.S.; Lee, E.W.; Kim, Y.M.; Kim, T.H. Pancreatic lipase inhibitory activity of phlorotannins isolated from Eisenia bicyclis. Phytother. Res. 2013, 27, 148–151. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Kim, H.R.; Chung, H.Y.; Choi, J.S. Anti-hyperlipidemic effect of an edible brown algae, Ecklonia stolonifera, and its constituents on poloxamer 407-induced hyperlipidemic and cholesterol-fed rats. Arch. Pharm. Res. 2008, 31, 1564–1571. [Google Scholar] [CrossRef]
- Moon, H.E.; Ahn, B.R.; Jung, H.A.; Choi, J.S. Inhibitory activity of Ecklonia stolonifera and its isolated phlorotannins against Cu 2+-induced low-density lipoprotein oxidation. Fish. Sci. 2012, 78, 927–934. [Google Scholar] [CrossRef]
- Kang, H.S.; Kim, H.R.; Byun, D.S.; Son, B.W.; Nam, T.J.; Choi, J.S. Tyrosinase inhibitors isolated from the edible brown alga Ecklonia stolonifera. Arch. Pharm. Res. 2004, 27, 1226. [Google Scholar] [CrossRef] [PubMed]
- Agostinho, P.; Cunha, R.A.; Oliveira, C. Neuroinflammation, oxidative stress and the pathogenesis of Alzheimer’s disease. Curr. Pharm. Des. 2010, 16, 2766–2778. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Ghosh, K.K. Recent advances in the antioxidant therapies for Alzheimer’s disease: Emphasis on natural antioxidants. In Pathology, Prevention and Therapeutics of Neurodegenerative Disease; Springer: Singapore, 2019; pp. 253–263. [Google Scholar]
- Ceriello, A.; Esposito, K.; Piconi, L.; Ihnat, M.A.; Thorpe, J.E.; Testa, R.; Boemi, M.; Giugliano, D. Oscillating glucose is more deleterious to endothelial function and oxidative stress than mean glucose in normal and type 2 diabetic patients. Diabetes 2008, 57, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Perše, M. Oxidative stress in the pathogenesis of colorectal cancer: Cause or consequence? BioMed Res. Int. 2013, 2013, 725710. [Google Scholar] [CrossRef] [PubMed]
- Trouba, K.J.; Hamadeh, H.K.; Amin, R.P.; Germolec, D.R. Oxidative stress and its role in skin disease. Antioxid. Redox Signal. 2002, 4, 665–673. [Google Scholar] [CrossRef]
- McCall, A.L. Insulin therapy and hypoglycemia. Endocrinol. Metab. Clin. 2012, 41, 57–87. [Google Scholar] [CrossRef]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown alga Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef]
- Moon, H.E.; Islam, M.N.; Ahn, B.R.; Chowdhury, S.S.; Sohn, H.S.; Jung, H.A.; Choi, J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory phlorotannins from edible brown algae, Ecklonia stolonifera and Eisenia bicyclis. Biosci. Biotechnol. Biochem. 2011, 75, 1472–1480. [Google Scholar] [CrossRef]
- Agarwal, S.; Manchanda, P.; Vogelbaum, M.A.; Ohlfest, J.R.; Elmquist, W.F. Function of the blood-brain barrier and restriction of drug delivery to invasive glioma cells: Findings in an orthotopic rat xenograft model of glioma. Drug Metab. Dispos. 2013, 41, 33–39. [Google Scholar] [CrossRef]
- Osseni, R.; Debbasch, C.; Christen, M.O.; Rat, P.; Warnet, J.M. Tacrine-induced reactive oxygen species in a human liver cell line: The role of anethole dithiolethione as a scavenger. Toxicol. In Vitro 1999, 13, 683–688. [Google Scholar] [CrossRef]
- Heemels, M.T. Neurodegenerative diseases. Nature 2016, 539, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.G.; Postuma, R. Premotor and non-motor features of Parkinson’s disease. Curr. Opin. Neurol. 2014, 27, 434. [Google Scholar] [CrossRef] [PubMed]
- Ingelsson, M. Alpha-synuclein oligomers—neurotoxic molecules in Parkinson’s disease and other lewy body disorders. Front. Neurosci. 2016, 10, 408. [Google Scholar] [CrossRef]
- Meletis, C.D.; Zabriskie, N. Natural approaches for optimal sleep. Altern. Complement. Ther. 2008, 14, 181–188. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Wang, S.; Wacker, D.; Levit, A.; Che, T.; Betz, R.M.; McCorvy, J.D.; Venkatakrishnan, A.; Huang, X.P.; Dror, R.O.; Shoichet, B.K. D4 dopamine receptor high-resolution structures enable the discovery of selective agonists. Science 2017, 358, 381–386. [Google Scholar] [CrossRef]
- Chien, E.Y.; Liu, W.; Zhao, Q.; Katritch, V.; Han, G.W.; Hanson, M.A.; Shi, L.; Newman, A.H.; Javitch, J.A.; Cherezov, V. Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 2010, 330, 1091–1095. [Google Scholar] [CrossRef]
- Mashiba, H.; Matsunaga, K.; Gojobori, M. Effect of immunochemotherapy with OK-432 and yeast cell wall on the activities of peritoneal macrophages of mice. Jpn. J. Cancer Res. 1979, 70, 687–692. [Google Scholar]
- Bogo, V.; Jacobs, A.; Weiss, J. Behavioral toxicity and efficacy of WR-2721 as a radioprotectant. Radiat. Res. 1985, 104, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Jagetia, G.C. Radioprotective potential of plants and herbs against the effects of ionizing radiation. J. Clin. Biochem. Nutr. 2007, 40, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.; Morison, W.L.; Sauder, D.N. Photoaging: Mechanisms and repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R. Cutaneous aging: Chronologic versus photoaging. Photodamage 1995, 1995, 123–135. [Google Scholar]
- Gilchrest, B.A. Skin aging and photoaging: An overview. J. Am. Acad. Dermatol. 1989, 21, 610–613. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. 2000, 18, 1135. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef]
- Birkedal-Hansen, H.; Moore, W.; Bodden, M.; Windsor, L.; Birkedal-Hansen, B.; DeCarlo, A.; Engler, J. Matrix metalloproteinases: A review. Crit. Rev. Oral Biol. Med. 1993, 4, 197–250. [Google Scholar] [CrossRef]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Kerkelä, E.; Saarialho-Kere, U. Matrix metalloproteinases in tumor progression: Focus on basal and squamous cell skin cancer. Exp. Dermatol. 2003, 12, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Datta, S.; Wang, Z.; Li, X.Y.; Quan, T.; Chung, J.H.; Kang, S.; Voorhees, J.J. c-Jun–dependent inhibition of cutaneous procollagen transcription following ultraviolet irradiation is reversed by all-trans retinoic acid. J. Clin. Investig. 2000, 106, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, M.P.; White, L.A.; Schroen, D.J.; Benbow, U.; Brinckerhoff, C.E. Regulating expression of the gene for matrix metalloproteinase-1 (collagenase): Mechanisms that control enzyme activity, transcription, and mRNA stability. Crit. Rev. Eukaryot. Gene Expr. 1996, 6, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Scharffetter–Kochanek, K.; Brenneisen, P.; Wenk, J.; Herrmann, G.; Ma, W.; Kuhr, L.; Meewes, C.; Wlaschek, M. Photoaging of the skin from phenotype to mechanisms. Exp. Gerontol. 2000, 35, 307–316. [Google Scholar] [CrossRef]
- Scharffetter-Kochanek, K.; Wlaschek, M.; Brenneisen, P.; Schauen, M.; Blaudschun, R.; Wenk, J. UV-induced reactive oxygen species in photocarcinogenesis and photoaging. Biol. Chem. 1997, 378, 1247–1258. [Google Scholar] [PubMed]
- Kvam, E.; Tyrrell, R.M. Induction of oxidative DNA base damage in human skin cells by UV and near visible radiation. Carcinogenesis 1997, 18, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Robertson, J. Captopril in the treatment of clinical hypertension and cardiac failure. Lancet 1979, 314, 836–839. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Ko, S.C.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on angiotensin I-converting enzyme (ACE) inhibitory activity. Nutr. Res. Pract. 2011, 5, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Kido, M.; Mori, H.; Nakayama, Y.; Takahashi, M. Structure of an anti-plasmin inhibitor, eckol, isolated from the brown alga Ecklonia kurome Okamura and inhibitory activities of its derivatives on plasma plasmin inhibitors. Chem. Pharm. Bull. 1989, 37, 349–353. [Google Scholar] [CrossRef]
- Wise, R.; Hart, T.; Cars, O.; Streulens, M.; Helmuth, R.; Huovinen, P.; Sprenger, M. Antimicrobial resistance. Is a major threat to public health. Br. Med. J. 1998, 317, 609–610. [Google Scholar] [CrossRef]
- Pérez, M.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Lee, D.S.; Kang, Y.M.; Son, K.T.; Jeon, Y.J.; Kim, Y.M. Application of yeast Candida utilis to ferment Eisenia bicyclis for enhanced antibacterial effect. Appl. Biochem. Biotechnol. 2013, 171, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Eom, S.H.; Lee, E.H.; Jung, Y.J.; Kim, H.J.; Jo, M.R.; Son, K.T.; Lee, H.J.; Kim, J.H.; Lee, M.S.; et al. In vitro antibacterial and synergistic effect of phlorotannins isolated from edible brown seaweed Eisenia bicyclis against acne-related bacteria. Algae 2014, 29, 47–55. [Google Scholar] [CrossRef]
- Ahn, M.J.; Yoon, K.D.; Min, S.Y.; Lee, J.S.; Kim, J.H.; Kim, T.G.; Kim, S.H.; Kim, N.G.; Huh, H.; Kim, J. Inhibition of HIV-1 reverse transcriptase and protease by phlorotannins from the brown alga Ecklonia cava. Biol. Pharm. Bull. 2004, 27, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Rossetti, L. Minireview: Nutrient sensing and the regulation of insulin action and energy balance. Endocrinology 2003, 144, 5172–5178. [Google Scholar] [CrossRef]
- Gurevich-Panigrahi, T.; Panigrahi, S.; Wiechec, E.; Los, M. Obesity: Pathophysiology and clinical management. Curr. Med. Chem. 2009, 16, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Vazzana, N.; Santilli, F.; Sestili, S.; Cuccurullo, C.; Davi, G. Determinants of increased cardiovascular disease in obesity and metabolic syndrome. Curr. Med. Chem. 2011, 18, 5267–5280. [Google Scholar] [CrossRef]
- Youssef, W.I.; McCullough, A.J. Steatohepatitis in obese individuals. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 733–747. [Google Scholar] [CrossRef]
- Lowe, M.E. Pancreatic triglyceride lipase and colipase: Insights into dietary fat digestion. Gastroenterology 1994, 107, 1524–1536. [Google Scholar] [CrossRef]
- Croce, C.M. Oncogenes and cancer. N. Engl. J. Med. 2008, 358, 502–511. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer. 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Lagunoff, D.; Martin, T.; Read, G. Agents that release histamine from mast cells. Annu. Rev. Pharmacol. Toxicol. 1983, 23, 331–351. [Google Scholar] [CrossRef] [PubMed]
- Church, M.K.; Levi-Schaffer, F. The human mast cell. J. Allergy Clin. Immunol. 1997, 99, 155–160. [Google Scholar] [CrossRef]
- Xie, Q.W.; Whisnant, R.; Nathan, C. Promoter of the mouse gene encoding calcium-independent nitric oxide synthase confers inducibility by interferon gamma and bacterial lipopolysaccharide. J. Exp. Med. 1993, 177, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- He, P. Leucocyte/endothelium interactions and microvessel permeability: Coupled or uncoupled? Cardiovasc. Res. 2010, 87, 281–290. [Google Scholar] [CrossRef]
- Komarova, Y.A.; Mehta, D.; Malik, A.B. Dual regulation of endothelial junctional permeability. Sci. STKE 2007, 2007, re8. [Google Scholar] [CrossRef] [PubMed]
- Majno, G.; Palade, G. Studies on inflammation: I. The effect of histamine and serotonin on vascular permeability: An electron microscopic study. J. Biophys. Biochem. Cytol. 1961, 11, 571–605. [Google Scholar] [CrossRef]
- Ware, L.B.; Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1334–1349. [Google Scholar] [CrossRef]
- Dhillon, S.S.; Mahadevan, K.; Bandi, V.; Zheng, Z.; Smith, C.W.; Rumbaut, R.E. Neutrophils, nitric oxide, and microvascular permeability in severe sepsis. Chest 2005, 128, 1706–1712. [Google Scholar] [CrossRef]
- Cummings, B.S.; McHowat, J.; Schnellmann, R.G. Phospholipase A2s in cell injury and death. J. Pharmacol. Exp. Ther. 2000, 294, 793–799. [Google Scholar]
- Bomalaski, J.S.; Clark, M.A. Phospholipase A2 and arthritis. Arthritis Rheum. 1993, 36, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Nakatsuji, T.; Zhu, W.; Gallo, R.L.; Huang, C.M. Passive immunoprotection targeting a secreted CAMP factor of Propionibacterium acnes as a novel immunotherapeutic for acne vulgaris. Vaccine 2011, 29, 3230–3238. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Piao, M.S.; Lee, J.B.; Oh, J.S.; Kim, I.G.; Lee, S.C. Propionibacterium acnes stimulates pro-matrix metalloproteinase-2 expression through tumor necrosis factor-α in human dermal fibroblasts. J. Investig. Dermatol. 2008, 128, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Jalian, H.R.; Liu, P.T.; Kanchanapoomi, M.; Phan, J.N.; Legaspi, A.J.; Kim, J. All-trans retinoic acid shifts Propionibacterium acnes-induced matrix degradation expression profile toward matrix preservation in human monocytes. J. Investig. Dermatol. 2008, 128, 2777–2782. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L.H.; Davelaar, E.; Kolb, R.M.; Pennings, E.J.; Smit, N.P. Tyrosine and cysteine are substrates for blackspot synthesis in potato. Phytochemistry 1998, 49, 703–707. [Google Scholar] [CrossRef]
- Ando, H.; Kondoh, H.; Ichihashi, M.; Hearing, V.J. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Investig. Dermatol. 2007, 127, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, S.M.; Sok, C.H.; Hong, J.T.; Oh, J.Y.; Jeon, Y.J. Cellular activities and docking studies of eckol isolated from Ecklonia cava (Laminariales, Phaeophyceae) as potential tyrosinase inhibitor. Algae 2015, 30, 163–170. [Google Scholar]
- Kang, J.I.; Kim, S.C.; Kim, M.K.; Boo, H.J.; Jeon, Y.J.; Koh, Y.S.; Yoo, E.S.; Kang, S.M.; Kang, H.K. Effect of dieckol, a component of Ecklonia cava, on the promotion of hair growth. Int. J. Mol. Sci. 2012, 13, 6407–6423. [Google Scholar] [CrossRef]
- Chang, M.Y.; Han, S.Y.; Shin, H.C.; Byun, J.Y.; Rah, Y.C.; Park, M.K. Protective effect of a purified polyphenolic extract from Ecklonia cava against noise-induced hearing loss: Prevention of temporary threshold shift. Int. J. Pediatr. Otorhinolaryngol. 2016, 87, 178–184. [Google Scholar] [CrossRef]
- Djouad, F.; Guerit, D.; Marie, M.; Toupet, K.; Jorgensen, C.; Noel, D. Mesenchymal stem cells: New insights into bone regenerative applications. J. Biomater. Tissue Eng. 2012, 2, 14–28. [Google Scholar] [CrossRef]
- Gimble, J.M.; Nuttall, M.E. The relationship between adipose tissue and bone metabolism. Clin. Biochem. 2012, 45, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M. Bone marrow mesenchymal stem cell differentiation: Involvement in osteoporosis with obesity and diabetes. J. Bone Marrow Res. 2013, 1, e107. [Google Scholar]
- Karadeniz, F.; Ahn, B.N.; Kim, J.A.; Seo, Y.; Jang, M.S.; Nam, K.H.; Kim, M.; Lee, S.H.; Kong, C.S. Phlorotannins suppress adipogenesis in pre-adipocytes while enhancing osteoblastogenesis in pre-osteoblasts. Arch. Pharm. Res. 2015, 38, 2172–2182. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Scientific Name | Types of Algae | Extracts | References |
|---|---|---|---|
| Ecklonia cava | Brown algae | Methanol/Ethanol | [11,12] |
| Ecklonia stolonifera | Brown algae | Methanol/Ethanol | [13,14] |
| Ecklonia kurome | Brown algae | Methanol/Ethanol | [15,16] |
| Ecklonia maxima | Brown algae | Methanol | [17] |
| Eisenia bicyclis | Brown algae | Methanol | [18] |
| Eisenia arborea | Brown algae | Methanol:Chloroform | [19] |
| Myagropsis myagroides | Brown algae | Ethanol | [20] |
| Pharmacological Effect | Experimental Methods | Pathway/Mode | Application | References |
|---|---|---|---|---|
| Antioxidant | Radical 2,2-diphenyl-1-picrylhydrazyl (DPPH), hydroxyl, peroxyl, and superoxide anion scavenging activity using the electron spin resonance spectrometry (ESR) technique. Cellular reactive oxygen species (ROS) determination using Dichloro-dihydro-fluorescein diacetate (DCFH-DA). Measurement of cellular glutathione (GSH) level. Myeloperoxidase (MPO) assay. | Showed significant radical scavenging activity. Significantly reduced ROS level at 50 μM concentration. Showed moderate inhibitory activity against membrane protein oxidation. Moderately enhanced cellular GSH level. Inhibited MPO activity moderately, inhibited ROS generation in kidney homogenate | In vitro | [41] |
| Heme oxygenase (HO)-1 activity/expression level via immunoprecipitation and Western blotting. | Regulated HO-1 via Nrf2 nuclear translocation and extracellular related kinase (Erk), Phosphoinositide 3-kinase (PI3K)/protein kinase B (PKB, also known as Akt) modulation. | In vitro | [42] | |
| DPPH radical scavenging assay, measurement of hydrogen peroxide, catalase activity, and lipid peroxidation assay. Western blotting, electrophoretic mobility shift assay (EMSA), and transient transfection and NF-κB luciferase assay. | Exhibited significant scavenging of DPPH radical, hydrogen peroxide (H2O2), hydroxy radical, intracellular ROS, prevention of lipid peroxidation. Phosphorylated extracellular signal-regulated kinase and activity of NF-κB. | In vitro | [43] | |
| Measurement of ROS and Ca2+ levels using dihydrorhodamine (DHR) 123 and Rhod-2 acetoxymethyl-ester (AM) dye. Mitochondrial succinate dehydrogenase activity. Western blot analysis, reverse transcriptase polymerase chain reaction (RT-PCR), and measurement of manganese superoxide dismutase (MnSOD) activity. | Decreased H2O2-induced mitochondrial ROS levels and attenuated ROS-generated mitochondrial Ca2+ levels. Prevented dysfunction of mitochondria following ROS and Ca2+ induction. Induction of MnSOD following activation of AMP-activated protein kinase/forkhead box O3a (AMPK/FoxO3a). | In vitro | [44] | |
| Radical scavenging activity using dichloro-dihydro-fluorescein (DCF) method, lipid peroxidation assay, and image analysis. Measurement of intracellular Ca2+ level using Fluo-3 AM in HT22 cells. | Scavenged intracellular ROS and repressed accumulation of ROS, prevented of lipid peroxidation, inhibited H2O2-induced cell death, and Ca2+ release in HT22 cells. | In vitro | [11] | |
| Radical scavenging activity using DCF method, lipid peroxidation assay, and image analysis. Cell death evaluation using acridine orange. | Scavenged intracellular ROS, prevented of lipid peroxidation. Decreased cell death in zebrafish embryos induced by2,2’-Azobis(2-amidinopropane) dihydrochloride (AAPH). | In vitro | [21] | |
| Cellular lipid peroxidation assay qRT-PCR analysis, enzyme-linked immunosorbent assay (ELISA). | Attenuated cellular lipid peroxidation, expression level of inflammatory cytokines (tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and IL-8). | In vitro | [12] | |
| Weight gain test using methyl α-linolenate. | Showed weight gain indicating its antioxidant potential. | In vitro | [18] | |
| Hydrophilic oxygen radical absorbance capacity (H-ORAC) assay. | Scavenged peroxyl radicals induced by AAPH. | In vitro | [45] | |
| Anti-diabetic | Protein tyrosine phosphatase (PTP1B) and α-glucosidase inhibitory assay. | Inhibited α-glucosidase enzyme activity indicating its potential to delay the postprandial increase of blood glucose level. | In vitro | [17] |
| Advanced glycation end products (AGEs), α-amylase, rat lens aldose reductase (RLAR) inhibitory assay. | Inhibited AGEs formation, α-amylase, rat lens aldose reductase activity. | In vitro | [46] | |
| Spectrophotometric assay on carbohydrate-hydrolyzing enzymes. Starch and sucrose tolerance test. | Inhibited α-amylase and α-glucosidase enzyme activity. Enhanced glucose tolerance and reduced fasting blood glucose, insulin, fructosamine, and glycoalbumin level. | In vitro/ In vivo | [15] | |
| Hepatoprotective | 2-(4-Iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2 tetrazolium (WST-1) colorimetric assay. | Scavenged free radicals; inhibited trichloromethyl free radicals (CCl3·), and trichloromethyl peroxy radicals (CCl3OO·) through cytochrome P450 systems-related CCl4 metabolism | In vitro | [25] |
| Tacrine-induced cytotoxicity assay. | Altered intracellular glutathione concentrations leading to inhibition of ROS generation and lipid peroxidation. | In vitro | [47] | |
| Western immunoblot, confocal microscopic analysis. | Diminished the expression of Fas-mediated cell-death proteins (tBid, caspase-3, and poly (ADP-ribose) polymerase). Decreased the release of cytochrome c from mitochondria to cytosol. | In vitro | [14] | |
| Enzymatic colorimetric method. Terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling (TUNEL) staining. ELISA. Bicinchoninic acid assay (BCA). | Increased alanine transaminase (ALT) and aspartate aminotransferase (AST) levels in serum and ameliorated morphological liver injury in rat. Protected rat hepatocytes from CCl4-mediated apoptosis. Reduced malondialdehyde (MDA) formations, TNF-α, in IL-1β, IL-6 and IL-10. Enhanced superoxide dismutase (SOD), glutathione peroxidase (GSH-Px) activities. Increased GSH content. | In vivo | [48] | |
| Online HPLC- 2,2’-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid (ABTS) radical scavenging assay. | Scavenged ABTS radical and decreased oxidative stress in t-BHP induced cell death in HepG2 cells. | In vitro | [49] | |
| 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Nuclear staining with Hoechst 33342. | Reduced the formation of apoptotic bodies in ethanol induced damage in Chang liver cells | In vitro | [50] | |
| Neuroprotective | β-secretase 1 (BACE1) inhibition assay. | Potently inhibited BACE1 activity | In vitro | [51] |
| Cholinesterase enzyme inhibition assay. | Inhibited acetyl and butyl cholinesterase enzyme activity. | In vitro | [52] | |
| Monoamine oxidase (MAO) enzyme inhibition assay. | Exhibited good inhibition in MAOs-A and-B enzyme activity. | In vitro | [53] | |
| MTT assay in PC12 cells. Determination of ROS level in Aβ-treated PC12 cells. | Protected PC12 cells against Aβ-induced cytotoxicity. Inhibited intracellular ROS and Ca2+ generation. | In vitro | [54] | |
| GABAA–Benzodiazepine (BZD) receptor-binding assay and pentobarbital-induced sleep test. | Showed allosteric modulation of the GABAA–BZD receptor. | In vitro/ In vivo | [55] | |
| G-protein coupled receptors (GPCRs) cell based functional assays. | Exhibited dual human dopamine D3/D4 agonist activity. | In vitro | [56] | |
| Anti-photo aging | Luciferase reporter gene assay, Western blot analysis and RT-PCR. | Inhibition on NF-κB and AP-1 reporter activity. Inhibited the expression of matrix metalloproteinase 1 (MMP-1) in TNF-α-induced human dermal fibroblasts and collagen degradation by MMP-1. | In vitro | [57] |
| ROS level determination using DCF method in ultraviolet (UV) B radiated HaCaT cells, Lipid peroxidation assay, single-cell gel electrophoresis (Comet Assay), and protein carbonyl formation. Mitochondrial membrane potential (Δψm) analysis. | Decreased UVB-induced intracellular ROS level. Inhibited membrane lipid peroxidation, protein carbonylation and cellular DNA damage. Protected against UVB-induced apoptosis. | In vitro | [58] | |
| Real-time RT-PCR analysis, Western blot analysis, prostaglandin (PG) E2 enzyme immunoassay, immunohistochemical detection of proliferating nuclear cell antigen. | Suppressed the expression of cyclooxygenase-2 (COX-2) leading to decrease in COX-2 protein production in UVB-induced skin carcinogenesis. Suppressed the gene expression of PGE2 in UVB-induced skin carcinogenesis. Significantly decreased the proliferation percentage of the cells in the epidermis | In vitro | [59] | |
| DCFH-DA and MTT assay, Comet assays. | Reduction in intracellular ROS accumulation, Reduction in fragmentation and destruction of irradiated cells. | In vitro | [60] | |
| Measurement of ROS level in UVB induced photo damage in zebrafish embryos. Detection of nitric oxide using diaminofluorophore 4-amino-5-methylamino2′,7′difluorofluorescein diacetate (DAF-FM DA). | Reduced reactive oxygen species and nitric oxide levels. | In vivo | [61] | |
| Radio protective | Hematoxylin and eosin staining and immunohistochemistry of jejunal crypt cells. Western blot analysis. | Protected the viability of lymphocytes and intestinal cells from radiation induced apoptosis. Decreased pro-apoptotic p53 and Bax and increased antiapoptotic B-cell lymphoma-2 (Bcl-2) concentration. | In vivo/ In vitro | [62] |
| Jejunal crypt assay. Apoptosis assay (hematoxylin-eosin and an in situ DNA end- labeling (ISEL) technique). | Enhanced the jejunal crypt survival. Protection against apoptosis induced by radiation in the mice jejunal crypts. | In vivo | [63] | |
| Colony-forming units (CFU) assay, alkaline comet assay, 3H-thymidine, incorporation assay, flow cytometry. | Enhances hematopoietic recovery. Reduced DNA damage in lymphocytes. Induced splenocyte proliferation. Increased the populations of T and B cells. | In vivo | [64] | |
| Intracellular reactive oxygen species measurement, single cell gel electrophoresis (Comet assay), 8-hydroxy-2′-deoxyguanosine (8-OHdG) assay and lipid peroxidation assay, mitochondrial membrane potential (Δψ) analysis electrophoretic mobility shift assay transient transfection and activator protein (AP)-1 luciferase assay. | Scavenging effect on reactive oxygen species, inhibition of damage of cellular DNA and peroxidation of membrane lipid in γ-ray irradiated cells. Protected against apoptosis by increasing Bcl-2 expression and decreasing Bax expression in γ-ray irradiated cells. Suppression of the SEK1-JNK-AP-1 pathway induced by γ-ray irradiation. | In vitro | [65] | |
| Anti-bacterial and Anti-viral | Disk-diffusion assay, time–kill assay, checkerboard dilution test, antibacterial activity assay. | Antibacterial activity against methicillin-resistant Staphylococcus aureus, Salmonella species and food-borne pathogenic bacteria with additive effect of eckol and ampicillin. Reduction of CFU. | In vitro | [29] |
| Chemiluminescent neuraminidase inhibition assay. Recombinant influenza virus neuraminidase inhibition assay. | Inhibition of influenza virus neuraminidase binding to free enzyme and/or product-bound enzyme. | In vitro | [66] | |
| Plaque reduction assay (Co-, pre- and post-exposure antiviral activity), qRT-PCR. | Inhibitory activity against viral hemorrhagic septicemia virus (VHSV) in the fathead minnow (FHM) cell line. Significantly induced inflammatory cytokine responses (interleukin (IL)-1β, IL-6, and IFN-γ) and interferon (IFN)-α/β, interferon stimulated gene (ISG) 15 and Mx were significantly activated. | In vitro/ In vivo | [67] | |
| Severe acute respiratory syndrome coronavirus 3C-like proteinase (SARS-CoV 3CLpro) cis-cleavage assay. | Inhibition through SARS-CoV 3CLpro trans/cis-cleavage. | In vitro | [68] | |
| Anti-inflammatory | The flux of albumin in a dual chamber system monitored in lipopolysaccharide (LPS)-mediated human umbilical vein endothelial cells (HUVECs) and in mice. Acetic acid-induced vascular permeability in mice. | Inhibited LPS-mediated barrier disruption and trans endothelial migration of leukocytes to human endothelial cells. Suppressed acetic acid induced-hyper permeability and carboxymethylcellulose-induced leukocytes migration in vivo. | In vitro/ In vivo | [69] |
| Competitive ELISA for high mobility group box 1 (HMGB1), permeability assay in vitro and in vivo, cell–cell adhesion assay, migration assay in vitro and in vivo. | Inhibited HMGB1 release and HMGB1 induced barrier disruption. Suppressed the expressions of cell adhesion molecule (CAM) (VCAM-1, ICAM-1 and E-selectin), inhibited the binding of monocytes to HMGB1-stimulated endothelial cells and inhibited trans-endothelial migration (TEM). | In vitro/ In vivo | [24] | |
| Assays of mouse ear edema induced by arachidonate (AA), 12-O-tetradecanoylphorbol-13-acetate (TPA), and oxazolone (OXA). Cell stimulation, anti-degranulation assay and qRT-PCR | Suppressed mouse ear swelling and the release of histamine, leukotriene B4, prostaglandin E2, and mRNA expression, and/or the activity of COX-2 in rat basophilic leukemia (RBL)-2H3 cells, inhibited delayed-type (type IV) allergic reactions induced by oxazolone | In vivo | [70] | |
| RNA extraction, RT-PCR, Western blot analysis | Inhibited the expression and production of proinflammatory mediators and cytokines in HaCat cells. Inhibited phosphorylation of Akt and activation of NF-κb mediated by Propionibacterium acnes. | In vitro | [71] | |
| Secretory phospholipase A2s, soybean lipoxygenase (LOX), 5-lipoxygenase, and COX-1 and COX-2 inhibitory assay. | Inhibition on secretory phospholipase A2s, LOX and COX activity. | In vitro | [72] | |
| Anti-histamine | Histamine release assay. Rat basophilic leukemia (RBL)-2H3 cell stimulation and assay for β-hexosaminidase release. Flow cytometry analysis. | Inhibition on the degranulation of KU812 cells stimulated by immunoglobulin E (IgE) and anti-IgE antibodies. Inhibition on the degranulation of RBL-2H3 cells mediated by IgE stimulation via a granular enzyme (β-hexosaminidase). Inhibited binding of IgE and FcεRI receptor in KU812 cells. | In vitro | [73] |
| Assay of hyaluronidase. | Showed inhibition in hyaluronidase activity. | In vitro | [74] | |
| Anti-hypertensive | Angiotensin converting enzyme (ACE) inhibitory activity assay. Determination of nitric oxide (NO) production. | ACE inhibitory and peroxynitrite scavenging properties. | In vitro | [28] |
| Anti-cancer | Flow cytometric analysis, soft agar colony formation assay, and invasion assay. PI3K kinase assay, Raf-1 kinase assay, and activated Ras affinity precipitation assay. | Suppressed expression of the glioma stem-like cell markers and the self-renewal-related proteins without cell death. Inhibited PI3K-Akt and Ras-Raf-1-Erk signaling pathways. | In vitro | [75] |
| MTT reduction assay on a limited variety of cancer cell lines. | Exhibited anti-proliferation effect. | In vitro | [76] | |
| Anti-coagulant | Anti-α2 plasmin, anti-α2 macroglobulin, anti-α1 anti thrombin (AT), anti-ATIII, anti-ATIII-heparin complex, anti-CI-inactivator activity assays and assay of inhibition of proteases. Evaluation on effects of fibrinolysis in human plasma induced by urokinase. | Potentiation of thrombolytic activity. Enhanced the effects fibrinolysis in human plasma induced by urokinase. | In vitro | [16] |
| Anticoagulation assay and thrombin activity assay and Factor Xa (FXa) activity assay. ELISA for plasminogen activator inhibitor (PAI)-1 and tissue plasminogen activator (t-PA). | Prolonged activated partial thromboplastin time (aPTT) and prothrombin time (PT) significantly and inhibited the generation of thrombin or FXa in human umbilical vein endothelial cells (HUVECs). Inhibited TNF-α induced PAI-1 production and the ratio between PAI-1 and t-PA (procoagulant and hypofibrinolysis activities). | In vitro/ In vivo | [77] | |
| Anti-obesity | Measurement of fat mass. Plasma analysis and intraperitoneal glucose tolerance test. Analysis of mRNA by quantitative real-time PCR. Quantification of liver triglyceride (TG) content. | Reduction of body weight, adiposity, ALT, and cholesterol Reduction of blood glucose levels. Increased mRNA expression of adipogenic ttyt 777 genes, and mRNA expression of inflammatory cytokines. Decreased macrophage marker gene. Inhibited intrahepatic lipid accumulation and hepatic lipogenic gene mRNA expression. | In vivo | [78] |
| Glucose utilization assay and triglyceride accumulation assay. Western blot analysis. | Suppressed glucose utilization, TG build-up, and downregulation of CCAAT-enhancer-binding proteins (C/EBP)α in 3T3-L1 adipocytes. | In vitro | [27] | |
| Assay of pancreatic lipase activity. | Inhibited pancreatic lipase activity. | In vitro | [79] | |
| Anti-hyperlipidemic | Enzymatic colorimetric methods for biochemical estimations by using commercial kits. | Reduced the level of TG, total cholesterol (TC), and low-density lipoprotein-cholesterol (LDL-C) and increased level of the high-density lipoprotein-cholesterol (HDLC). | In vivo | [80] |
| Thiobarbituric acid reactive substances (TBARS) assay. | Inhibitory activity against Cu2+-induced LDL oxidation and conjugated diene formation (TBARS assay). | In vitro | [81] | |
| Anti-tyrosinase | Measurement of cellular melanin contents and tyrosinase activity in B16F10 melanoma cells. Western blot analysis. | Suppressed tyrosinase activity and melanin synthesis. Decreased the expression of cellular tyrosinase enzyme, tyrosinase-related protein (TRP) 1, and TRP2. | In vitro | [82] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manandhar, B.; Paudel, P.; Seong, S.H.; Jung, H.A.; Choi, J.S. Characterizing Eckol as a Therapeutic Aid: A Systematic Review. Mar. Drugs 2019, 17, 361. https://doi.org/10.3390/md17060361
Manandhar B, Paudel P, Seong SH, Jung HA, Choi JS. Characterizing Eckol as a Therapeutic Aid: A Systematic Review. Marine Drugs. 2019; 17(6):361. https://doi.org/10.3390/md17060361
Chicago/Turabian StyleManandhar, Bandana, Pradeep Paudel, Su Hui Seong, Hyun Ah Jung, and Jae Sue Choi. 2019. "Characterizing Eckol as a Therapeutic Aid: A Systematic Review" Marine Drugs 17, no. 6: 361. https://doi.org/10.3390/md17060361
APA StyleManandhar, B., Paudel, P., Seong, S. H., Jung, H. A., & Choi, J. S. (2019). Characterizing Eckol as a Therapeutic Aid: A Systematic Review. Marine Drugs, 17(6), 361. https://doi.org/10.3390/md17060361