The Ameliorating Effect of Plasma Protein from Tachypleus tridentatus on Cyclophosphamide-Induced Acute Kidney Injury in Mice


Abstract
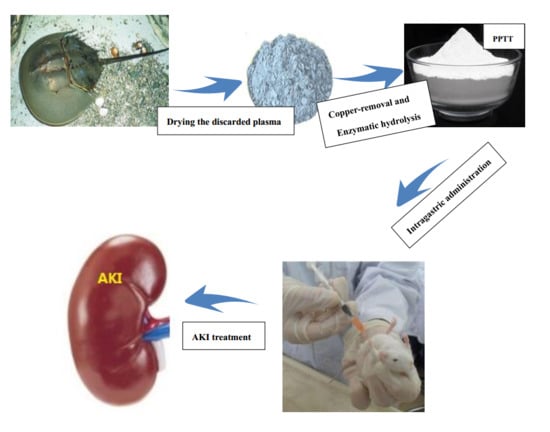
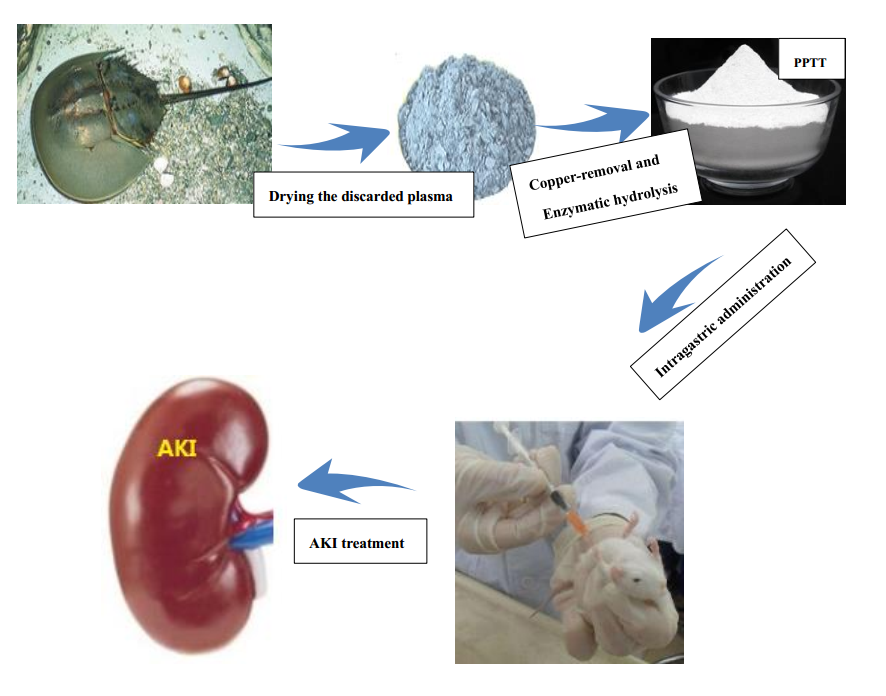
1. Introduction
2. Results
2.1. Attenuation of PPTT on the Negative Effects on Body Mass, Kidney Mass, Kidney Index in CTX-Induced Mice
2.2. PPTT Relieves CTX -Induced Renal Oxidative Stress
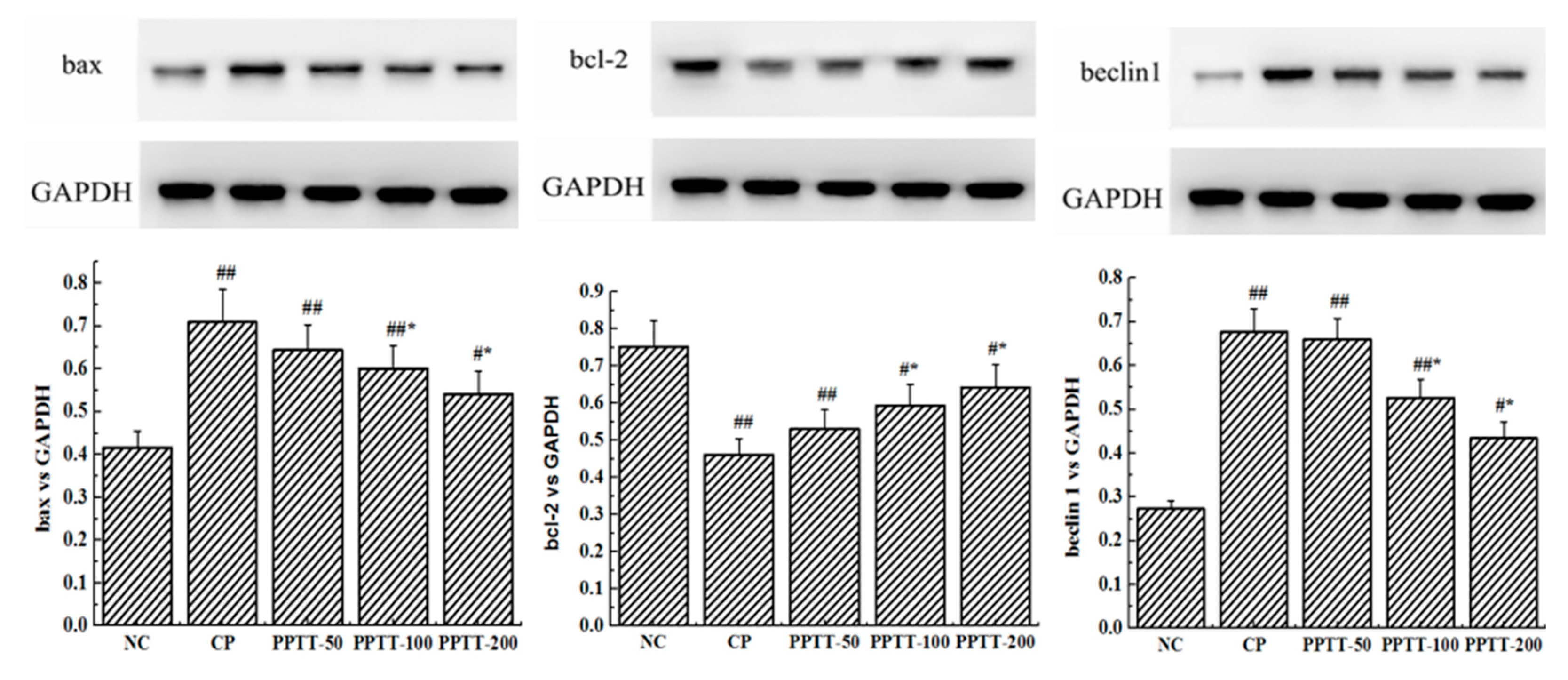
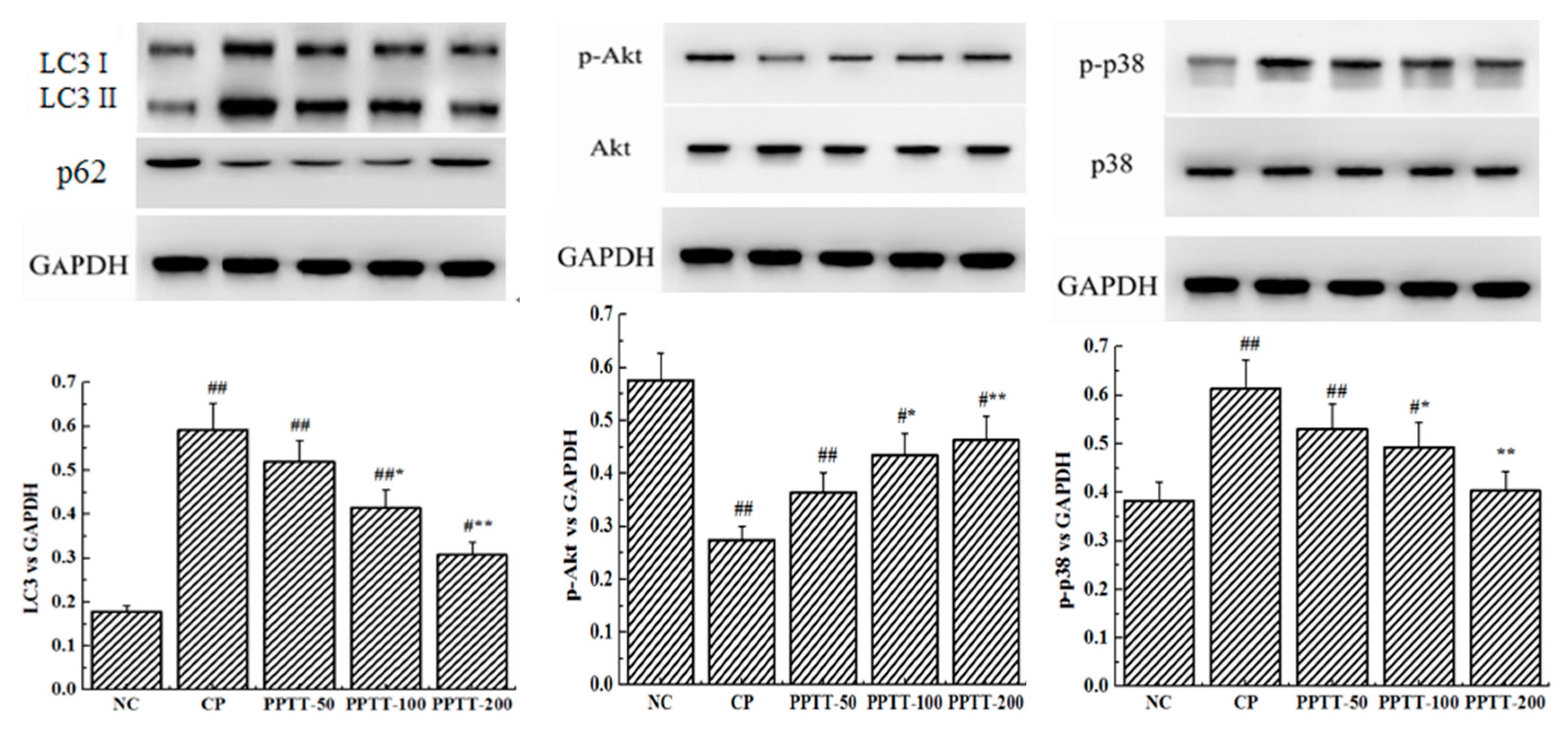
2.3. Western Blotting Analyses
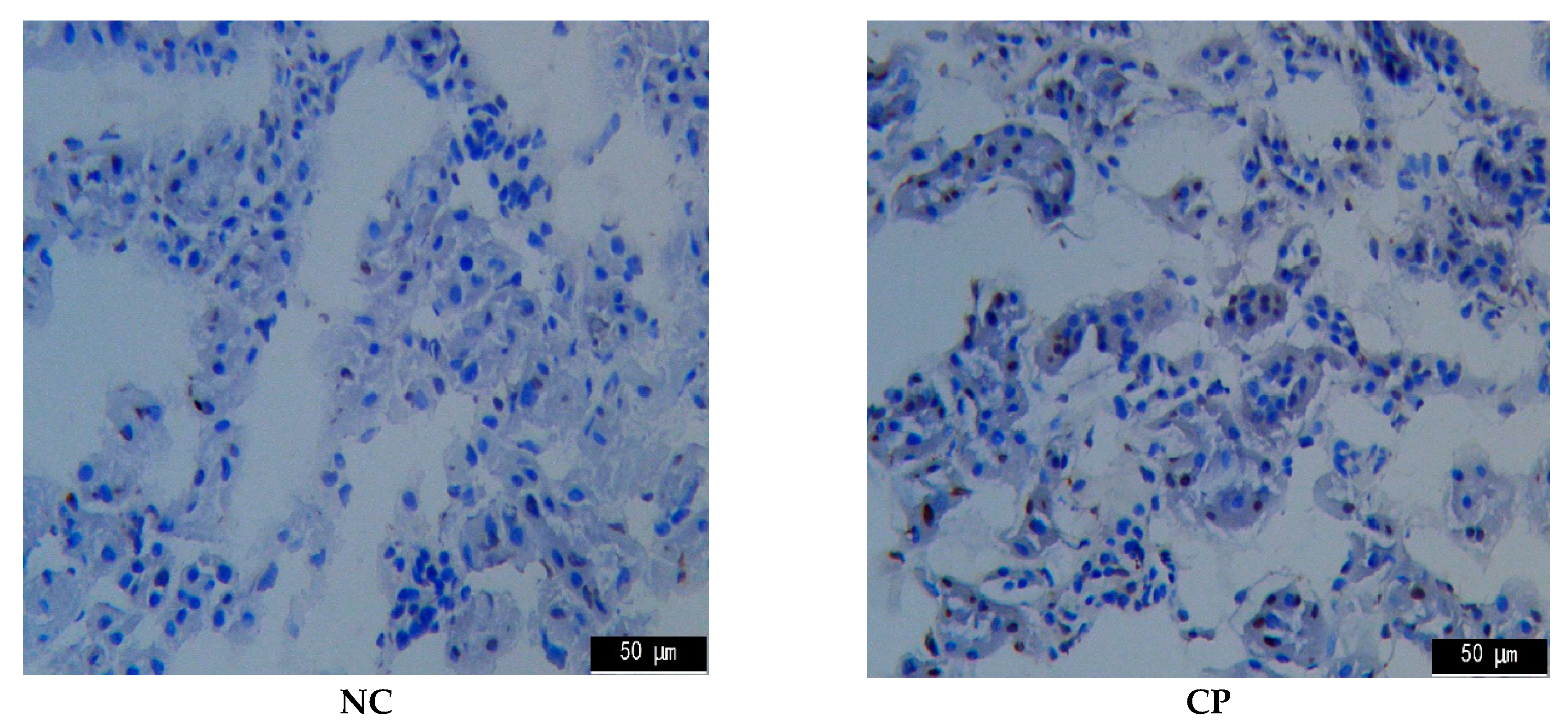
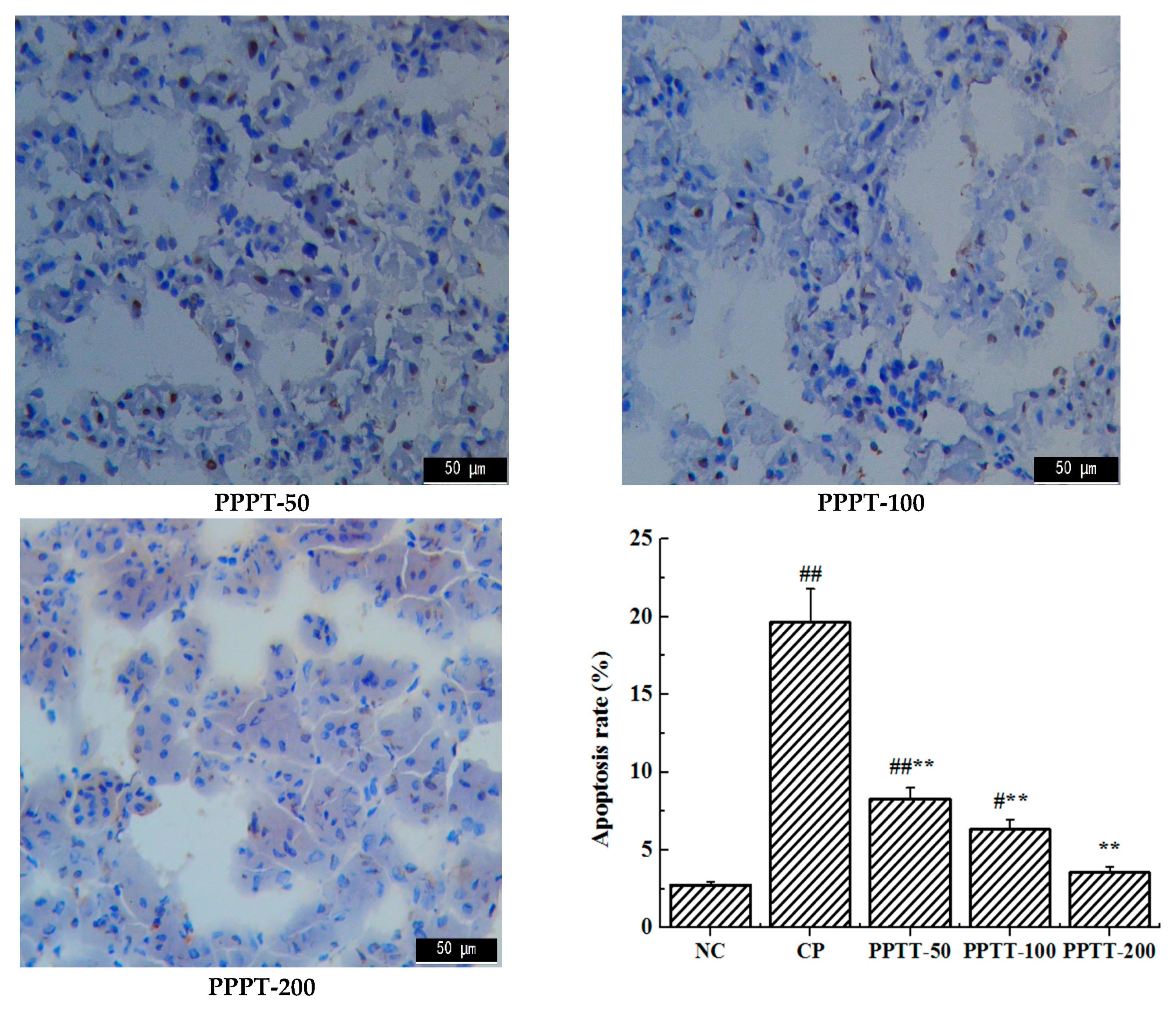
2.4. TUNEL Assay
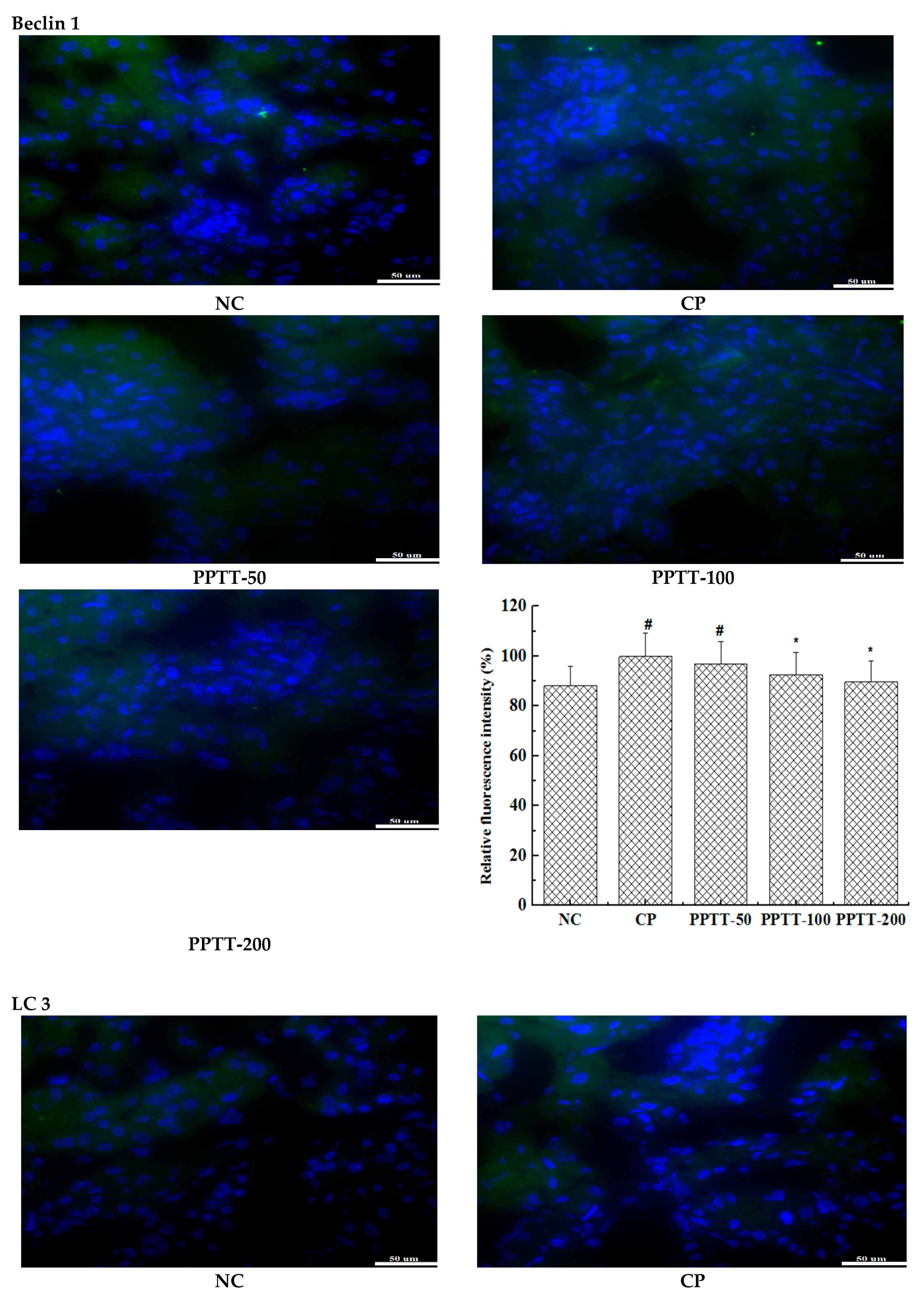
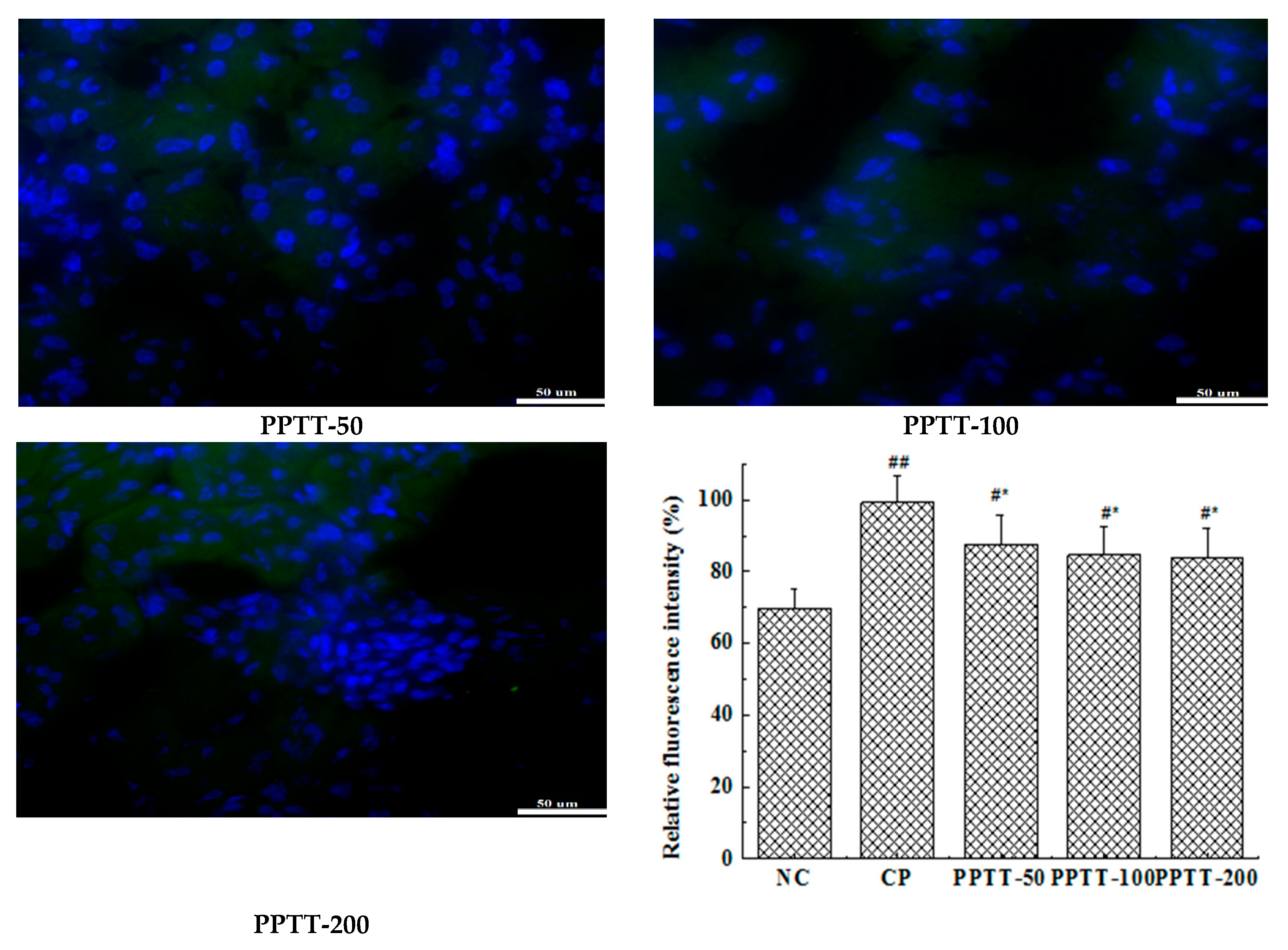
2.5. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Preparation of Plasma Protein from Tachypleus tridentatus (PPTT)
4.2. Animal Models and Treatment Protocol
4.3. Detection of Kidney Index
4.4. Measurement of Parameters Related to Oxidative Stress in Liver
4.5. Western Blotting Analysis
4.6. TUNEL Assay
4.7. Immunofluorescence Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hsu, R.K.; McCulloch, C.E.; Dudley, R.A.; Dudley, R.A.; Lo, L.J.; Hsu, C.Y. Temporal changes in incidence of dialysis-requiring AKI. J. Am. Soc. Nephrol. 2013, 24, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.X.; Yang, L. Epidemiology of acute kidney injury. Chin. J. Blood Purif. 2017, 16, 8–10. [Google Scholar]
- Moignet, A.; Hasanali, Z.; Zambello, R.; Pavan, L.; Bareau, B.; Tournilhac, O.; Roussel, M.; Fest, T.; Awwad, A.; Baab, K.; et al. Cyclophosphamide as a first-line therapy in LGL leukemia. Leukemia 2014, 28, 1134–1136. [Google Scholar] [CrossRef]
- Wagner, T. Ifosfamide Clinical Pharmacokinetics. Clin. Pharmacok. 1994, 26, 439–456. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular mechanisms of acrolein toxicity: Relevance to human disease. Toxicol. Sci. 2015, 143, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Tsai-Turton, M.; Luong, B.T.; Tan, Y.; Luderer, U. Cyclophosphamide-induced apoptosis in COV434 human granulosa cells involves oxidative stress and glutathione depletion. Toxicol. Sci. 2007, 98, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A.; El-Hosseiny, L.S.; Nashashibi, M.A. Protective effects of essential oils as natural antioxidants against hepatotoxicity induced by cyclophosphamide in mice. PLoS ONE 2016, 11, e0165667. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, D.; Shi, L.; Liu, X.W.; Zhang, Y.B.; Tong, C.C.; Song, D.Q.; Hou, M.X. Blueberry anthocyanins-enriched extracts attenuate cyclophosphamide-Induced cardiac injury. PLoS ONE 2015, 10, e0127813. [Google Scholar] [CrossRef]
- Gore, P.R.; Prajapati, C.P.; Mahajan, U.B. Protective effect of thymoquinone against cyclophosphamide-induced hemorrhagic cystitis through inhibiting DNA damage and upregulation of Nrf2 expression. Int. J. Biol. Sci. 2016, 12, 944–953. [Google Scholar] [CrossRef]
- Liu, Q.; Lin, X.M.; Li, H.; Yuan, J.; Peng, Y.P.; Dong, L.; Dai, S.J. Paeoniflorin ameliorates renal function in cyclophosphamide-induced mice via AMPK suppressed inflammation and apoptosis. Biomed. Pharmacother. 2016, 84, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.G.; Wei, J.G.; Jin, G.; Huo, G.H.; Cheng, W.; Xie, H.W.; Zhang, Y. Analysis of free amino acids and fatty acids in serum from horseshoe crab. Hubei Agric. Sci. 2007, 46, 606–607. [Google Scholar]
- Almeida, O.F.; Conde, G.L.; Crochemore, C.; Demeneix, B.A.; Fischer, D.; Hassan, A.H.; Meyer, M.; Holsboer, F.; Michaelidis, T.M. Subtle shifts in the ratio between pro- and antiapoptotic molecules after activation of corticosteroid receptors decide neuronal fate. FASEB J. 2000, 14, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Gorin, Y. The kidney: An organ in the front line of oxidative stress-associated pathologies. Antioxid. Redox Signal. 2016, 25, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Modaresi, A.; Nafar, M.; Sahraei, Z. Oxidative stress in chronic kidney disease. Iran. J. Kidney Dis. 2015, 9, 165–179. [Google Scholar] [PubMed]
- Otunctemur, A.; Ozbek, E.; Cakir, S.S.; Dursun, M.; Cekmen, M.; Polat, E.C.; Ozcan, L.; Somay, A.; Ozbay, N. Beneficial effects montelukast, cysteinyl-leukotriene receptor antagonist, on renal damage after unilateral ureteral obstruction in rats. Int. Braz. J. Urol. 2015, 41, 279–287. [Google Scholar] [CrossRef]
- Ren, Y.; Du, C.; Shi, Y.; Wei, J.; Wu, H.; Cui, H. The siet1 activator, SRT1720, attenuates renal fibrosis by inhibiting CTGF and oxidative stress. Int. Mol. Med. 2017, 39, 1317–1324. [Google Scholar] [CrossRef]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. ROS are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef]
- Du, C.; Zhang, T.; Xiao, X.; Shi, Y.; Duan, H.; Ren, Y. Protease-activated receptor-2 promotes kidney tubular epithelial inflammation by inhibiting autophagy via the PI3K/Akt/m TOR signalling pathway. Biochem. J. 2017, 474, 2733–2747. [Google Scholar] [CrossRef]
- Tetz, L.M.; Cheng, A.A.; Korte, C.S.; Giese, R.W.; Wang, P.; Harris, C.; Meeker, J.D.; Loch-caruso, R. Mono-2-ethylhexyl phthalate induces oxidative stress responses in human placental cells in vitro. Toxicol. Appl. Pharmacol. 2013, 268, 47–54. [Google Scholar] [CrossRef]
- Pan, W.R.; Chen, Y.L.; Hsu, H.C.; Chen, W.J. Antimicrobial peptide GW-H1-induced apoptosis of human gastric cancer AGS cell line is enhanced by suppression of autophagy. Mol. Cell Biochem. 2015, 400, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Yun, J.P.; Yang, Y.N.; Hua, F.; Zhang, X.W.; Lin, H.; Lv, X.X.; Zhang, P.C.; Hu, Z.W. A novel ECG analog 4-(S)-(2,4,6-trimethylthiobenzyl)-epigallocatechin gallate selectively induces apoptosis of B16-F10 melanoma via activation of autophagy and ROS. Sci. Rep. 2017, 7, 42194. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.M.; Du, J.X.; Kang, X.H.; He, L.Z.; Wen, Y.M. A Hydrolysis Method of Plasma Protein of Tachypleus tridentatus. CN Patent CN201610651742.X, 9 August 2016. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Mass (g) | Kidney Mass (g) | Kidney Index (%) |
|---|---|---|---|
| NC | 24.71 ± 1.36 | 0.20 ± 0.01 | 0.81 ± 0.05 |
| CP | 20.54 ± 1.04 ## | 0.24 ± 0.02 ## | 1.17 ± 0.10 ## |
| PPTT-50 | 20.94 ± 0.83 ## | 0.23 ± 0.02 ## | 1.10 ± 0.05 ## |
| PPTT-100 | 21.43 ± 1.19 | 0.22 ± 0.01 #* | 1.03 ± 0.06 # |
| PPTT-200 | 22.67 ± 2.05 | 0.21 ± 0.02 ** | 0.93 ± 0.09 * |
| Groups | Doses (mg/kg∙BW) | MDA (nmol/mg prot) | GSH-Px (U/mg prot) | SOD (U/mg prot) | CAT (U/mg prot) |
|---|---|---|---|---|---|
| NC | / | 3.37 ± 0.32 | 481.46 ± 35.28 | 901.05 ± 53.27 | 38.38 ± 2.80 |
| CP | / | 4.72 ± 0.71 ## | 346.88 ± 32.61 ## | 786.68 ± 75.34 ## | 22.01 ± 3.15 ## |
| PPTT-50 | 50 | 4.01 ± 0.47 #* | 336.70 ± 28.97 ## | 836.44 ± 77.59 # | 22.69 ± 2.71 ## |
| PPTT-100 | 100 | 3.83 ± 0.44 # | 385.57 ± 35.04 ##* | 841.13 ± 68.31 #* | 23.28 ± 2.25 ## |
| PPTT-200 | 200 | 3.68 ± 0.39 ** | 406.76 ± 34.08 #* | 881.84 ± 84.48 ** | 27.20 ± 2.32 ##* |
| Antibody Name | Antibody Species | Brand | Item No. |
|---|---|---|---|
| bax | rabbit | Bio swamp | PAB30040 |
| bcl-2 | rabbit | Bio swamp | PAB30041 |
| Beclin 1 | rabbit | Bio swamp | PAB35215 |
| LC3 | rabbit | Bio swamp | MAB37400 |
| P62 | rabbit | Bio swamp | PAB33349 |
| Akt | rabbit | Bio swamp | MAB37305 |
| p-Akt | rabbit | Bio swamp | PAB43181-p |
| P38 | rabbit | Bio swamp | MAB37199 |
| p-p38 | rabbit | Bio swamp | PAB43506-p |
| GAPDH | rabbit | Bio swamp | PAB36264 |
| Goat Anti-Rabbit IgG | goat | Bio swamp | PAB160011 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.; Jing, M.; Zhang, G.; He, L.; Hong, P.; Deng, C. The Ameliorating Effect of Plasma Protein from Tachypleus tridentatus on Cyclophosphamide-Induced Acute Kidney Injury in Mice. Mar. Drugs 2019, 17, 227. https://doi.org/10.3390/md17040227
Kang X, Jing M, Zhang G, He L, Hong P, Deng C. The Ameliorating Effect of Plasma Protein from Tachypleus tridentatus on Cyclophosphamide-Induced Acute Kidney Injury in Mice. Marine Drugs. 2019; 17(4):227. https://doi.org/10.3390/md17040227
Chicago/Turabian StyleKang, Xinhuang, Mengyao Jing, Guoguang Zhang, Lanzheng He, Pengzhi Hong, and Chunmei Deng. 2019. "The Ameliorating Effect of Plasma Protein from Tachypleus tridentatus on Cyclophosphamide-Induced Acute Kidney Injury in Mice" Marine Drugs 17, no. 4: 227. https://doi.org/10.3390/md17040227
APA StyleKang, X., Jing, M., Zhang, G., He, L., Hong, P., & Deng, C. (2019). The Ameliorating Effect of Plasma Protein from Tachypleus tridentatus on Cyclophosphamide-Induced Acute Kidney Injury in Mice. Marine Drugs, 17(4), 227. https://doi.org/10.3390/md17040227