Target Identification of the Marine Natural Products Dictyoceratin-A and -C as Selective Growth Inhibitors in Cancer Cells Adapted to Hypoxic Environments

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
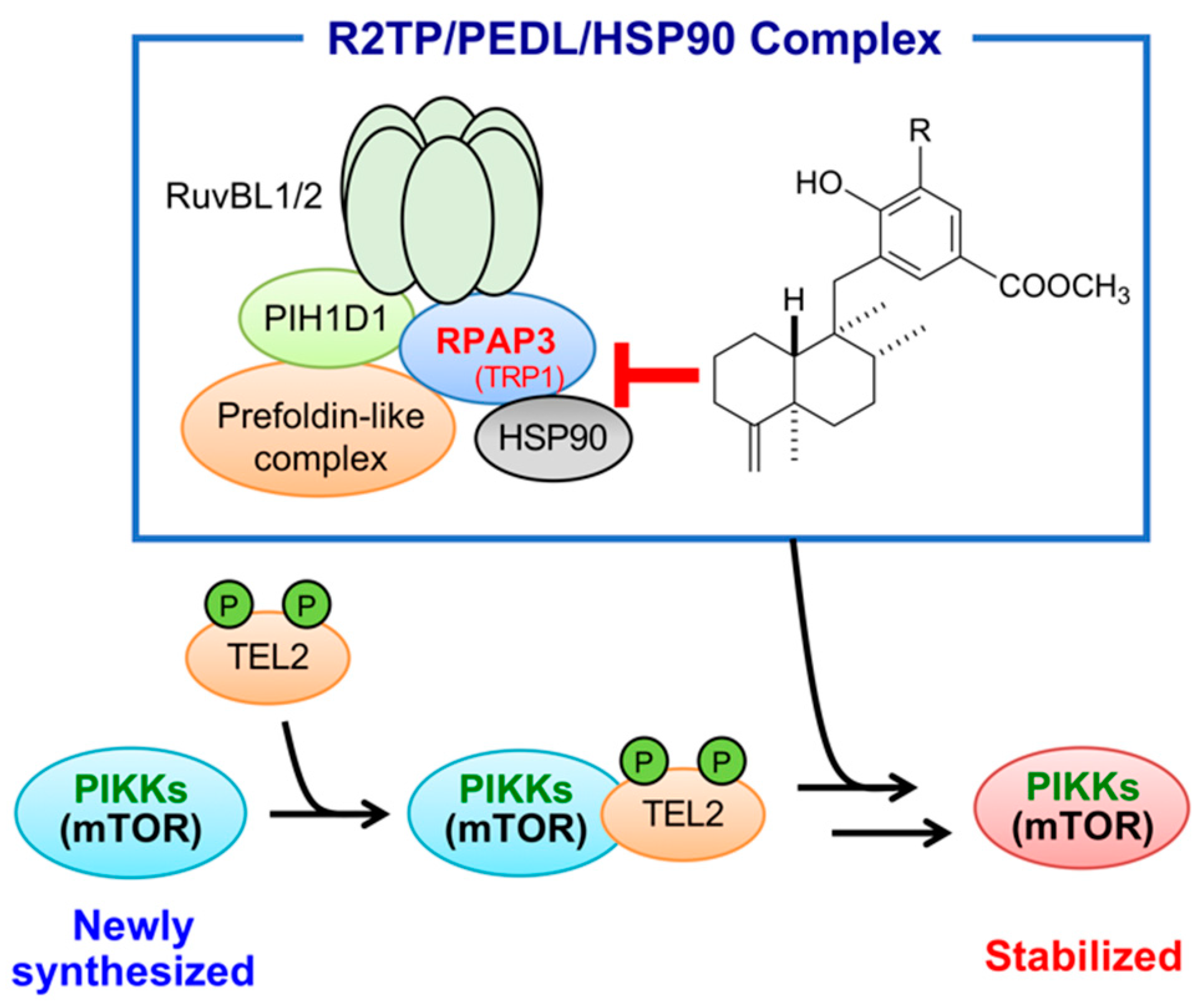
2. Results and Discussion
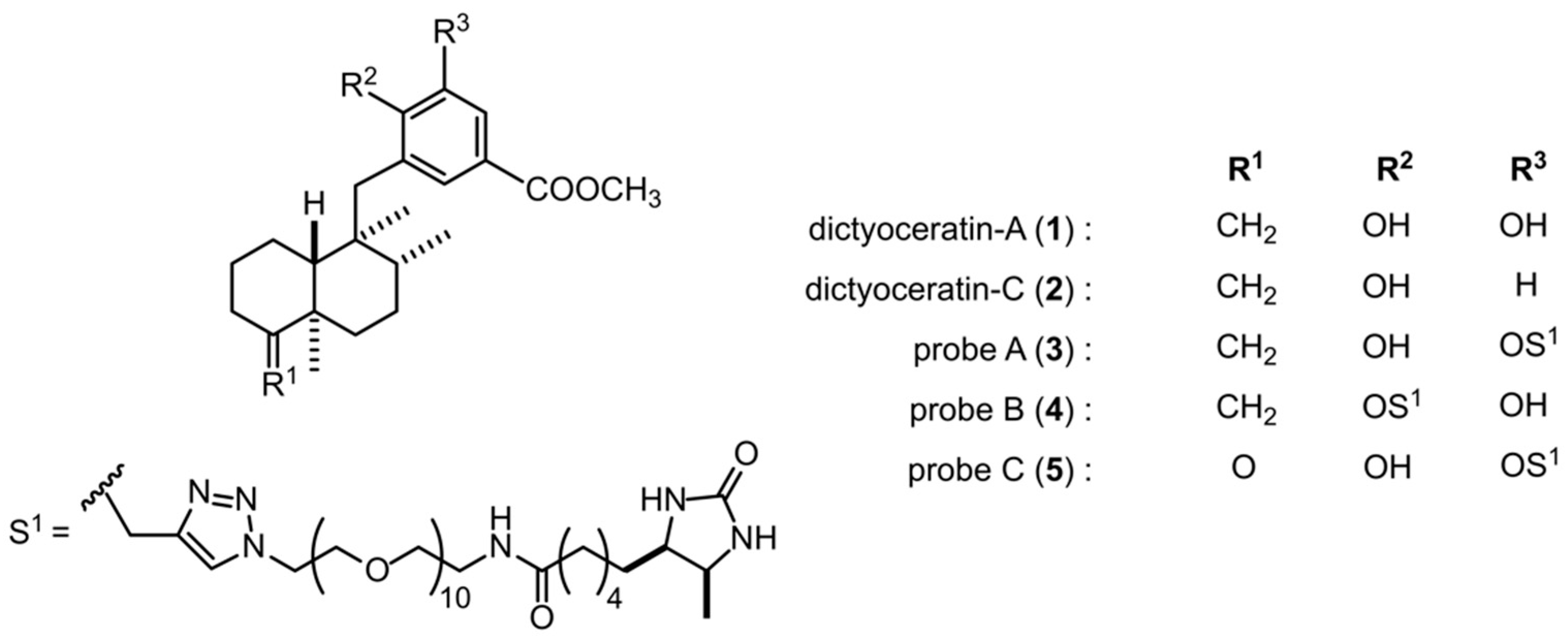
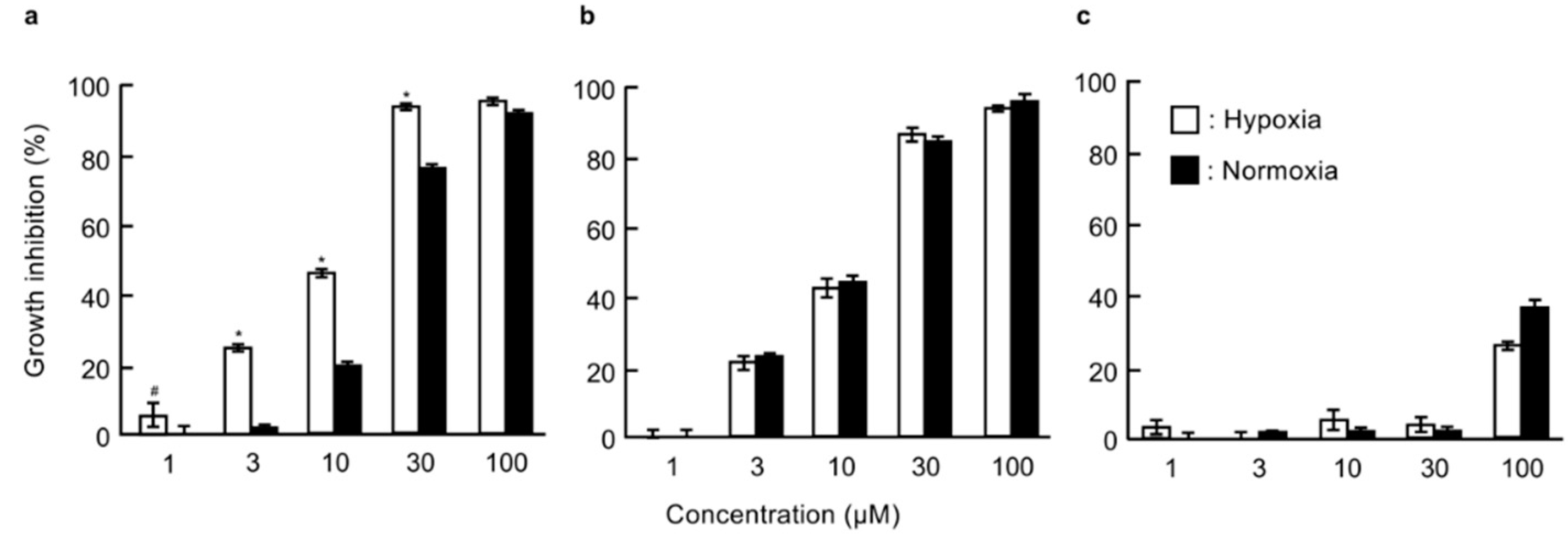
2.1. Effects of Probe Molecules on the Growth of DU145 Cells under Normoxic and Hypoxic Conditions
2.2. Analysis of Target Molecules Using Probe A (3) from a Peptide-Displayed Phage Library
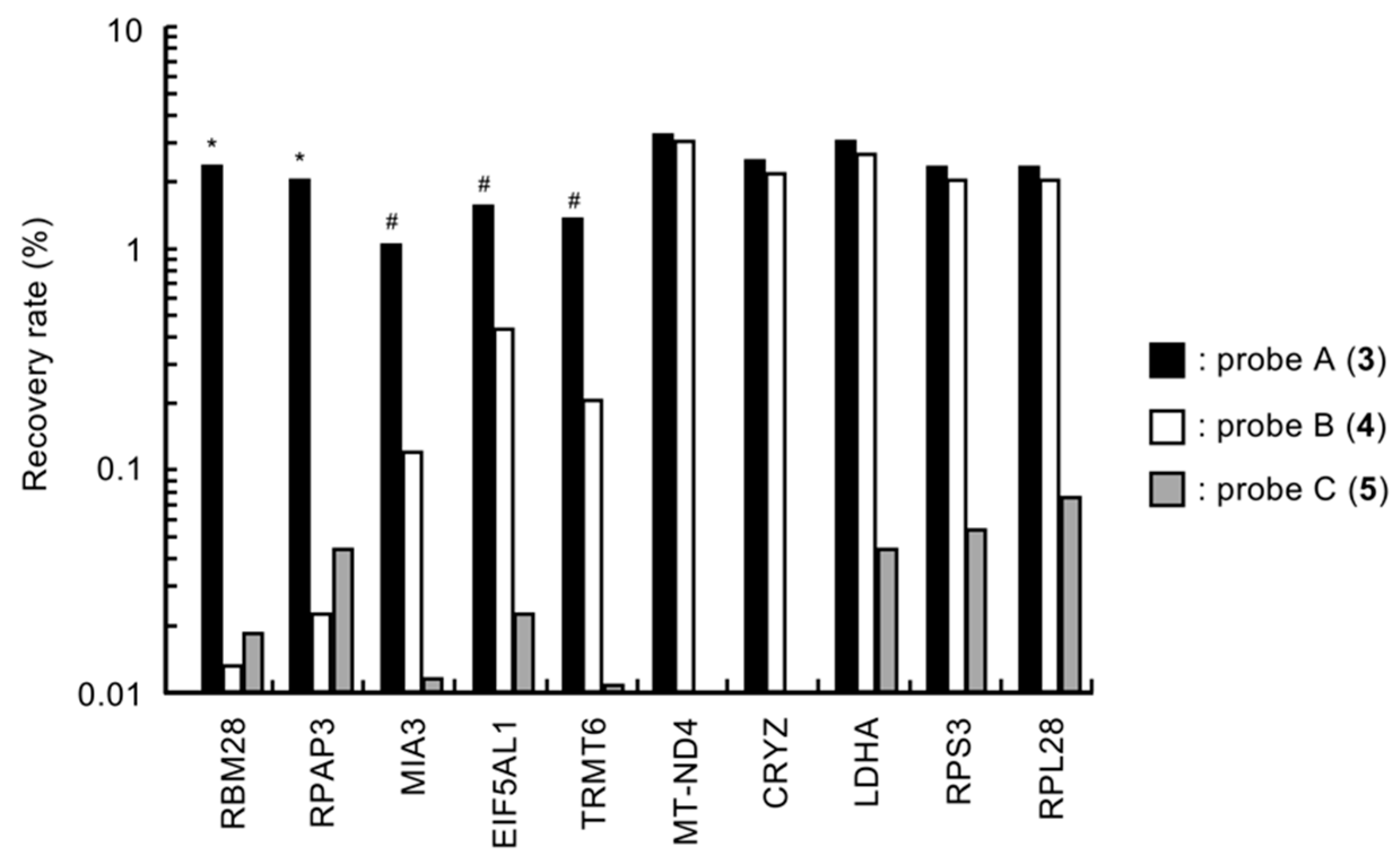
2.3. Binding Selectivity of the Selected Peptide-Displayed Phages against Probes A–C (3–5)
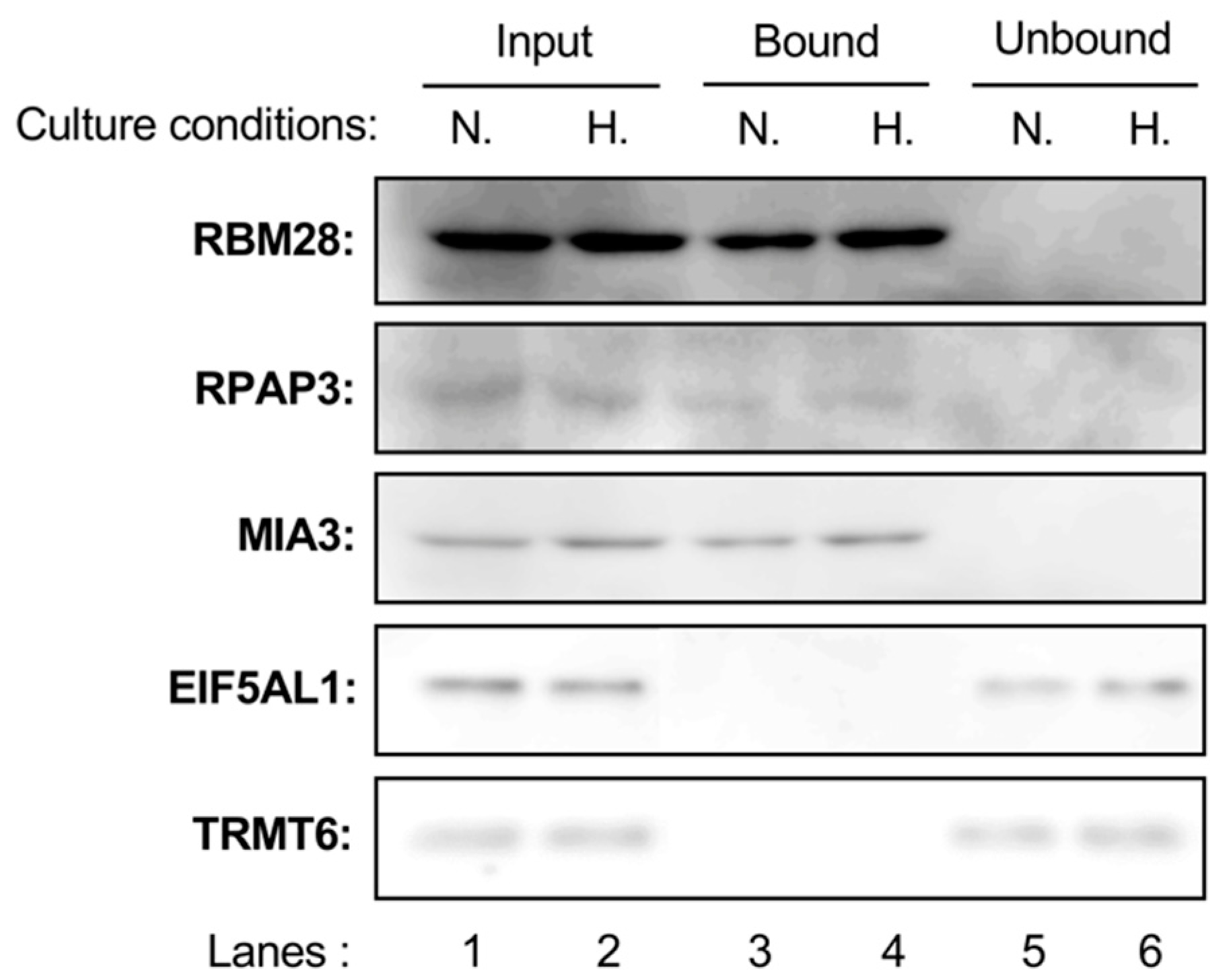
2.4. Binding Abilities of Probe A (3) with RBM28, RPAP3, MIA3, EIF5AL1, and TRMT6 in CELL lysates
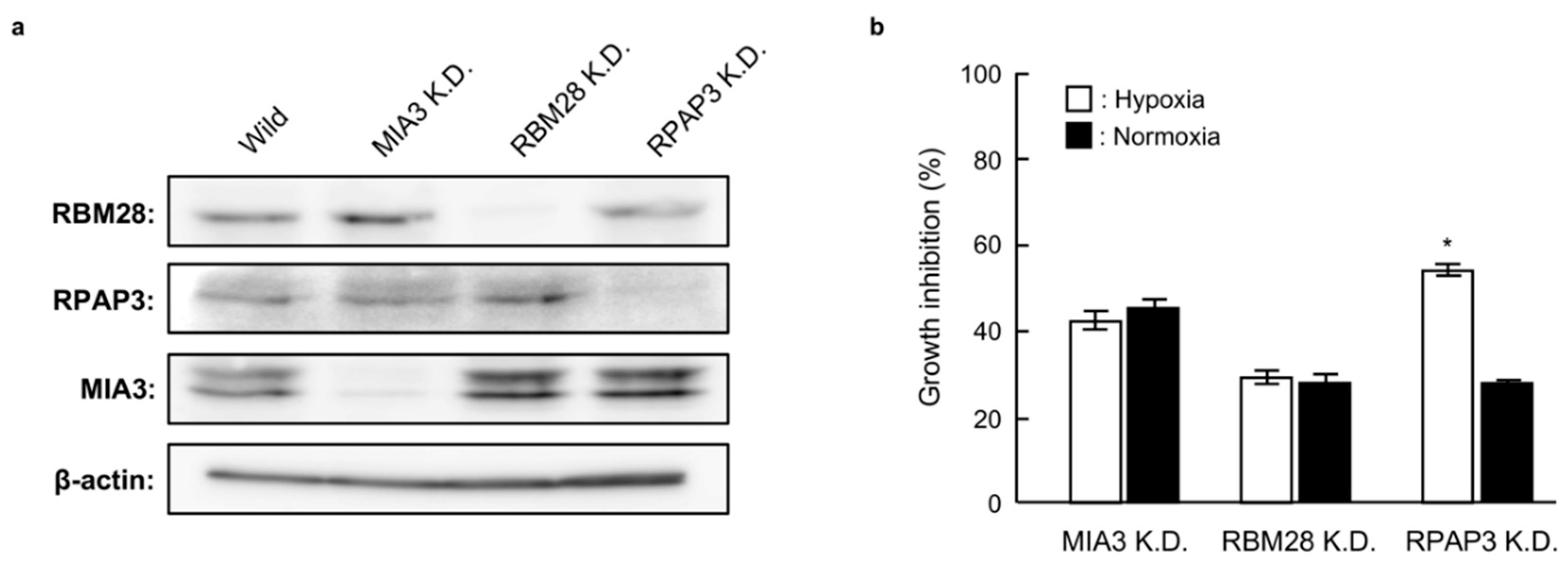
2.5. Growth of RBM28-, RPAP3-, and MIA3-Knockdown DU145 Cells under Normoxic and Hypoxic Conditions
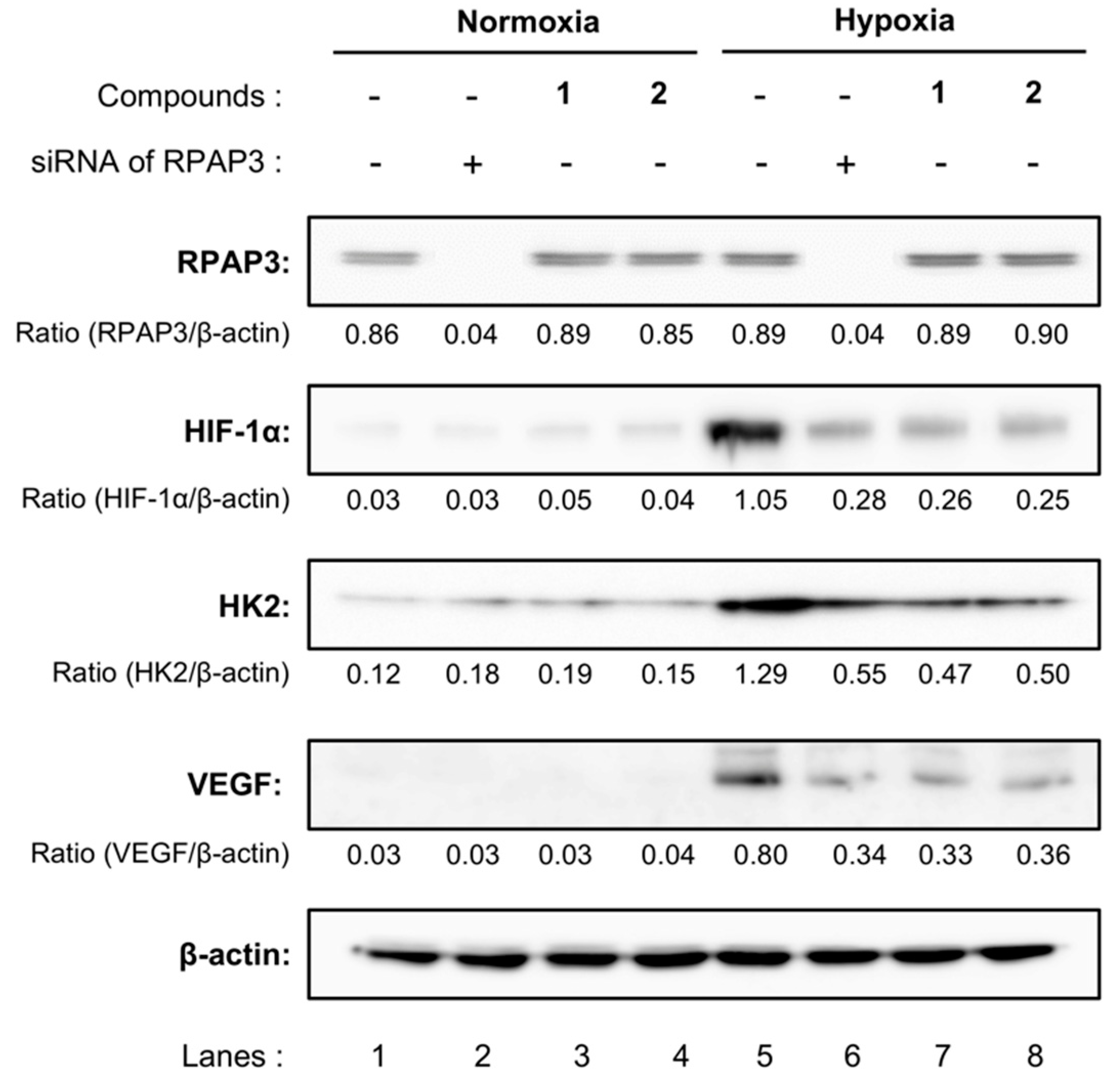
2.6. Characterization of RPAP3-Knockdown DU145 Cells
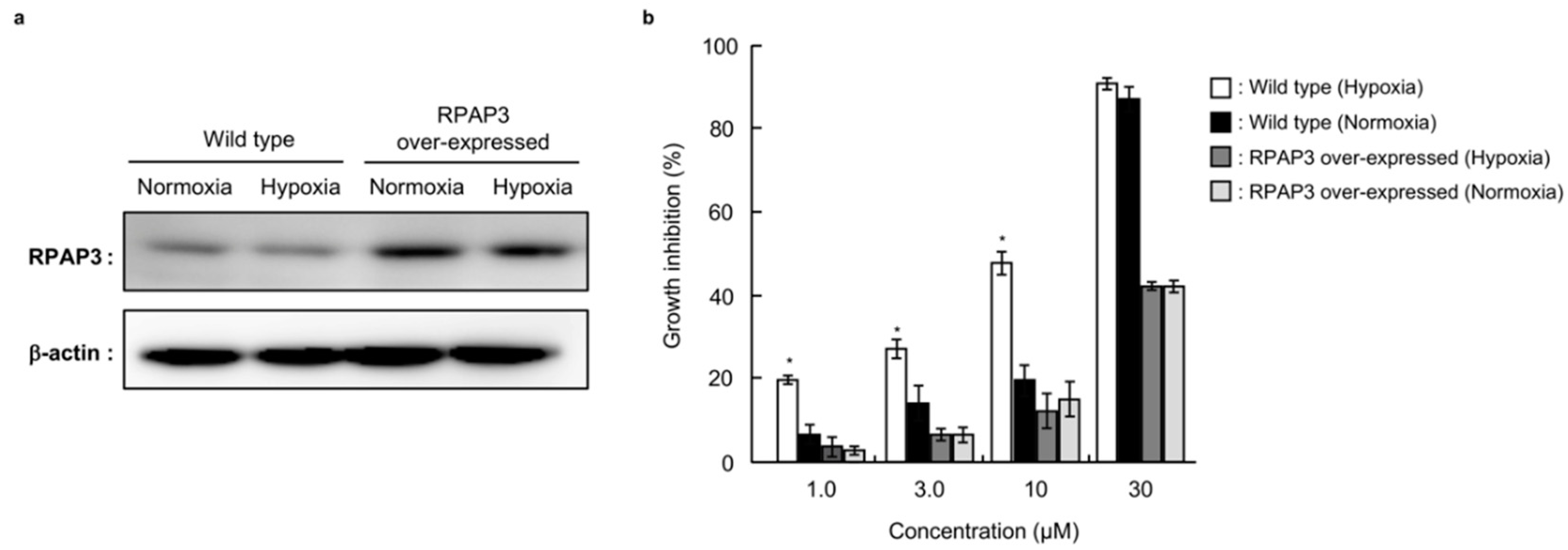
2.7. Effects of RPAP3 Overexpression on the Growth Inhibitory Activity of Dictyoceratin-A (1)
3. Materials and Methods
3.1. General Reagents and Materials
3.2. Cell Culture and Antiproliferative Activity
3.3. Synthesis of Probe Molecules
3.3.1. General Instruments
3.3.2. N-(32-Azido-3,6,9,12,15,18,21,24,27,30-decaoxadotriacontyl)-6-((4R,5S)-5-methyl-2-oxoimidazolidin-4-yl)hexanamide (6)
3.3.3. Methyl 4-Hydroxy-3-((1-(39-((4R,5S)-5-methyl-2-oxoimidazolidin-4-yl)-34-oxo-3,6,9,12,15,18,21,24,27,30-decaoxa-33-azanonatriacontyl)-1H-1,2,3-triazol-4-yl)methoxy)-5-(((1S,2R,4aR,8aR)-1,2,4a-trimethyl-5-methylenedecahydronaphthalen-1-yl)methyl)benzoate (3, probe A)
3.3.4. Methyl 3-hydroxy-4-((1-(39-((4R,5S)-5-methyl-2-oxoimidazolidin-4-yl)-34-oxo-3,6,9,12,15,18,21,24,27,30-decaoxa-33-azanonatriacontyl)-1H-1,2,3-triazol-4-yl)methoxy)-5-(((1S,2R,4aR,8aR)-1,2,4a-trimethyl-5-methylenedecahydronaphthalen-1-yl)methyl)benzoate (4, probe B)
3.3.5. Methyl 4-Hydroxy-3-(prop-2-yn-1-yloxy)-5-(((1S,2R,4aR,8aR)-1,2,4a-trimethyl-5-oxodecahydronaphthalen-1-yl)methyl)benzoate (10)
3.3.6. Methyl 4-Hydroxy-3-((1-(39-((4R,5S)-5-methyl-2-oxoimidazolidin-4-yl)-34-oxo-3,6,9,12,15,18,21,24,27,30-decaoxa-33-azanonatriacontyl)-1H-1,2,3-triazol-4-yl)methoxy)-5-(((1S,2R,4aR,8aR)-1,2,4a-trimethyl-5-oxodecahydronaphthalen-1-yl)methyl)benzoate (5, probe C)
3.4. Binding Ability of Probes A–C (3–5) to Proteins in Cell Lysates
3.5. Western Blotting Analysis
3.6. Generation of RPAP3-Overexpressing DU145 Cells
3.7. Preparation of MIA3-, RBM28-, and RPAP3-Knockdown DU145 Cells
3.8. Preparation of a Peptide-Display Phage Library and Screening of Probe A (3)-Binding Phages
3.9. Statistical Analaysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dewhirst, M.W.; Cao, Y.; Moeller, B. Cycling hypoxia and free radicals regulate angiogenesis and radiotherapy response. Nat. Rev. Cancer 2008, 8, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Lim, H.N.; Jung, H.J. Hovenia dulcis Thunb. and its active compound ampelopsin inhibit angiogenesis through suppression of VEGFR2 signaling and HIF-1α expression. Oncol. Rep. 2017, 38, 3430–3438. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.H.; Kim, S.H.; Hong, J.Y.; Park, H.J.; Oh, D.C.; Lee, S.K. Salternamide A Suppresses Hypoxia-Induced Accumulation of HIF-1α and Induces Apoptosis in Human Colorectal Cancer Cells. Mar. Drugs 2015, 13, 6962–6976. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.T.; Patel, P.R.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; Goey, A.K.; Cook, K.M.; Figg, W.D.; McMahon, J.B.; Schnermann, M.J.; et al. Structural Elucidation and Synthesis of Eudistidine A: An Unusual Polycyclic Marine Alkaloid that Blocks Interaction of the Protein Binding Domains of p300 and HIF-1α. J. Am. Chem. Soc. 2015, 137, 5569–5575. [Google Scholar] [CrossRef] [PubMed]
- Goey, A.K.; Chau, C.H.; Sissung, T.M.; Cook, K.M.; Venzon, D.J.; Castro, A.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; McMahon, J.B.; et al. Screening and Biological Effects of Marine Pyrroloiminoquinone Alkaloids: Potential Inhibitors of the HIF-1α/p300 Interaction. J. Nat. Prod. 2016, 79, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, K.A.; Hossain, C.F.; Zhang, L.; Bruick, R.K.; Zhou, Y.D.; Nagle, D.G. Laurenditerpenol, a new diterpene from the tropical marine alga Laurenciaintricata that potently inhibits HIF-1 mediated hypoxic signaling in breast tumor cells. J. Nat. Prod. 2004, 67, 2002–2007. [Google Scholar] [CrossRef]
- Mohammed, K.A.; Jadulco, R.C.; Bugni, T.S.; Harper, M.K.; Sturdy, M.; Ireland, C.M. Strongylophorines: Natural product inhibitors of hypoxia-inducible factor-1 transcriptional pathway. J. Med. Chem. 2008, 51, 1402–1405. [Google Scholar] [CrossRef]
- Arai, M.; Kawachi, T.; Setiawan, A.; Kobayashi, M. Hypoxia-selective growth inhibition of cancer cells by furospinosulin-1, a furanosesterterpene isolated from an Indonesian marine sponge. ChemMedChem 2010, 5, 1919–1926. [Google Scholar] [CrossRef]
- Arai, M.; Kawachi, T.; Kotoku, N.; Nakata, C.; Kamada, H.; Tsunoda, S.; Tsutsumi, Y.; Endo, H.; Inoue, M.; Sato, H.; et al. Furospinosulin-1, Marine Spongean Furanosesterterpene, Suppresses the Growth of Hypoxia-Adapted Cancer Cells by Binding to Transcriptional Regulators p54nrb and LEDGF/p75. ChemBioChem 2016, 17, 181–189. [Google Scholar] [CrossRef]
- Arai, M.; Kawachi, T.; Sato, H.; Setiawan, A.; Kobayashi, M. Marine spongian sesquiterpene phenols, dictyoceratin-C and smenospondiol, display hypoxia-selective growth inhibition against cancer cells. Bioorg. Med. Chem. Lett. 2014, 24, 3155–3157. [Google Scholar] [CrossRef] [PubMed]
- Sumii, Y.; Kotoku, N.; Fukuda, A.; Kawachi, T.; Sumii, Y.; Arai, M.; Kobayashi, M. Enantioselective synthesis of dictyoceratin-A (smenospondiol) and -C, hypoxia-selective growth inhibitors from marine sponge. Bioorg. Med. Chem. 2015, 23, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Sumii, Y.; Kotoku, N.; Fukuda, A.; Kawachi, T.; Arai, M.; Kobayashi, M. Structure-Activity Relationship and in Vivo Anti-Tumor Evaluations of Dictyoceratin-A and -C, Hypoxia-Selective Growth Inhibitors from Marine Sponge. Mar. Drugs 2015, 13, 7419–7432. [Google Scholar] [CrossRef] [PubMed]
- Takakusagi, Y.; Takakusagi, K.; Sugawara, F.; Sakaguchi, K. Use of phage display technology for the determination of the targets for small-molecule therapeutics. Expert Opin. Drug Discov. 2010, 5, 361–389. [Google Scholar] [CrossRef] [PubMed]
- Kakihara, Y.; Houry, W.A. The R2TP complex: Discovery and functions. Biochim. Biophys. Acta 2012, 1823, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Von Morgen, P.; Hořejší, Z.; Macurek, L. Substrate recognition and function of the R2TP complex in response to cellular stress. Front. Genet. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, P.; Al-Khoury, R.; Lavallée-Adam, M.; Faubert, D.; Jiang, H.; Poitras, C.; Bouchard, A.; Forget, D.; Blanchette, M.; Coulombe, B. High-resolution mapping of the protein interaction network for the human transcription machinery and affinity purification of RNA polymerase II-associated complexes. Methods 2009, 48, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Martino, F.; Pal, M.; Muñoz-Hernández, H.; Rodríguez, C.F.; Núñez-Ramírez, R.; Gil-Carton, D.; Degliesposti, G.; Skehel, J.M.; Roe, S.M.; Prodromou, C.; et al. RPAP3 provides a flexible scaffold for coupling HSP90 to the human R2TP co-chaperone complex. Nat. Commun. 2018, 9, 1501. [Google Scholar] [CrossRef]
- Boulon, S.; Pradet-Balade, B.; Verheggen, C.; Molle, D.; Boireau, S.; Georgieva, M.; Azzag, K.; Robert, M.C.; Ahmad, Y.; Neel, H.; et al. HSP90 and its R2TP/Prefoldin-like cochaperone are involved in the cytoplasmic assembly of RNA polymerase II. Mol. Cell 2010, 39, 912–924. [Google Scholar] [CrossRef]
- Abraham, R.T. PI 3-kinase related kinases: ‘Big’ players in stress-induced signaling pathways. DNA Repair 2004, 3, 883–887. [Google Scholar] [CrossRef]
- Wouters, B.G.; Koritzinsky, M. Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat. Rev. Cancer 2008, 8, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Horejsi, Z.; Takai, H.; Adelman, C.A.; Collis, S.J.; Flynn, H.; Maslen, S.; Skehel, J.M.; de Lange, T.; Boulton, S.J. CK2 phospho-dependent binding of R2TP complex to TEL2 is essential for mTOR and SMG1 stability. Mol. Cell 2010, 39, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Takai, H.; Wang, R.C.; Takai, K.K.; Yang, H.; de Lange, T. Tel2 regulates the stability of PI3K-related protein kinases. Cell 2007, 131, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hirsch, J.D.; Eslamizar, L.; Filanoski, B.J.; Malekzadeh, N.; Haugland, R.P.; Beechem, J.M.; Haugland, R.P. Easily reversible desthiobiotin binding to streptavidin, avidin, and other biotin-binding proteins: Uses for protein labeling, detection, and isolation. Anal. Biochem. 2002, 308, 343–357. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawachi, T.; Tanaka, S.; Fukuda, A.; Sumii, Y.; Setiawan, A.; Kotoku, N.; Kobayashi, M.; Arai, M. Target Identification of the Marine Natural Products Dictyoceratin-A and -C as Selective Growth Inhibitors in Cancer Cells Adapted to Hypoxic Environments. Mar. Drugs 2019, 17, 163. https://doi.org/10.3390/md17030163
Kawachi T, Tanaka S, Fukuda A, Sumii Y, Setiawan A, Kotoku N, Kobayashi M, Arai M. Target Identification of the Marine Natural Products Dictyoceratin-A and -C as Selective Growth Inhibitors in Cancer Cells Adapted to Hypoxic Environments. Marine Drugs. 2019; 17(3):163. https://doi.org/10.3390/md17030163
Chicago/Turabian StyleKawachi, Takashi, Shun Tanaka, Akinori Fukuda, Yuji Sumii, Andi Setiawan, Naoyuki Kotoku, Motomasa Kobayashi, and Masayoshi Arai. 2019. "Target Identification of the Marine Natural Products Dictyoceratin-A and -C as Selective Growth Inhibitors in Cancer Cells Adapted to Hypoxic Environments" Marine Drugs 17, no. 3: 163. https://doi.org/10.3390/md17030163
APA StyleKawachi, T., Tanaka, S., Fukuda, A., Sumii, Y., Setiawan, A., Kotoku, N., Kobayashi, M., & Arai, M. (2019). Target Identification of the Marine Natural Products Dictyoceratin-A and -C as Selective Growth Inhibitors in Cancer Cells Adapted to Hypoxic Environments. Marine Drugs, 17(3), 163. https://doi.org/10.3390/md17030163