Physicochemical and Biological Properties of Gelatin Extracted from Marine Snail Rapana venosa


 ,
,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Physicochemical and Structural Properties of Marine Gelatins
2.1.1. Yield Extraction and Gelatin Characteristics
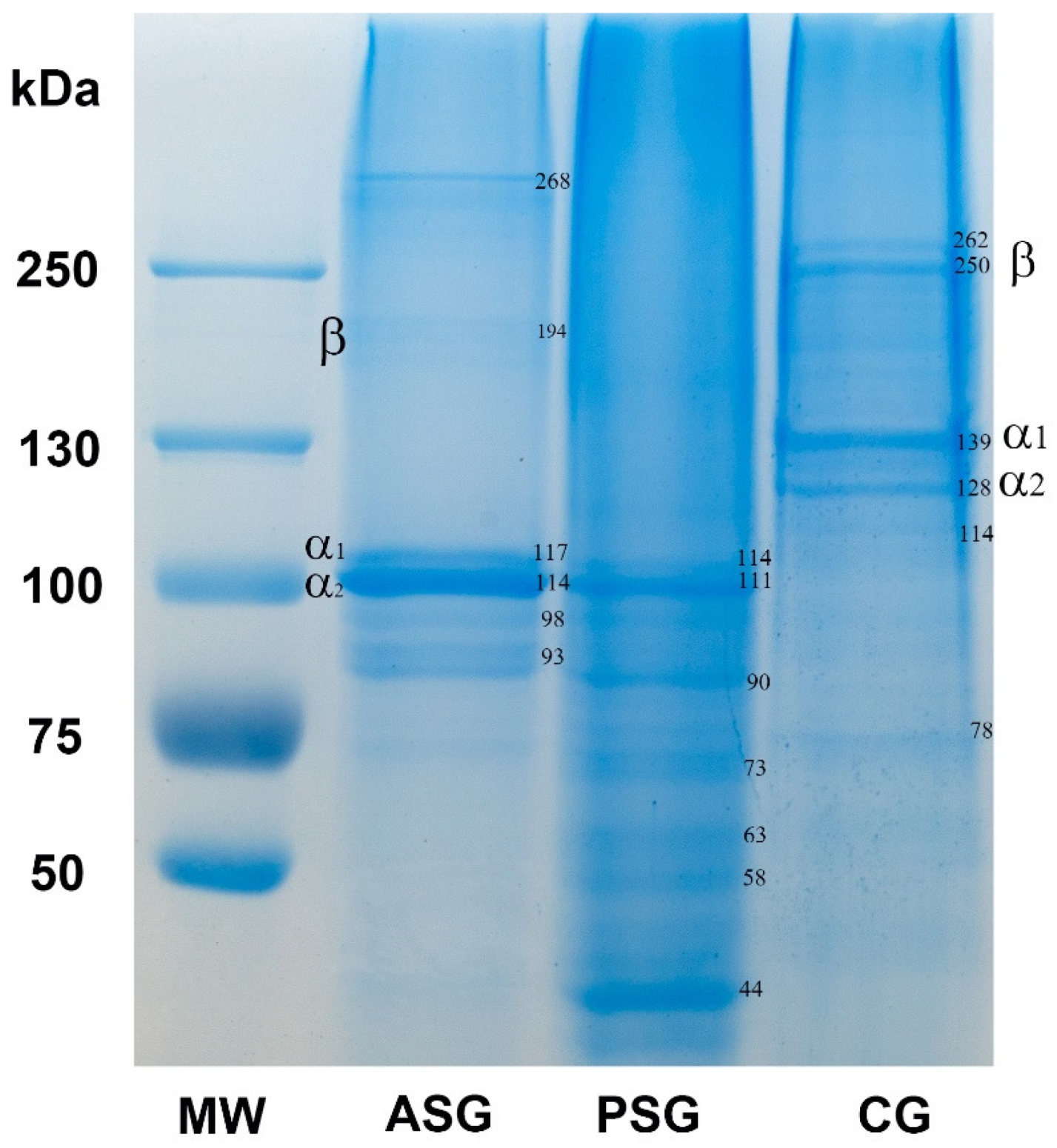
2.1.2. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
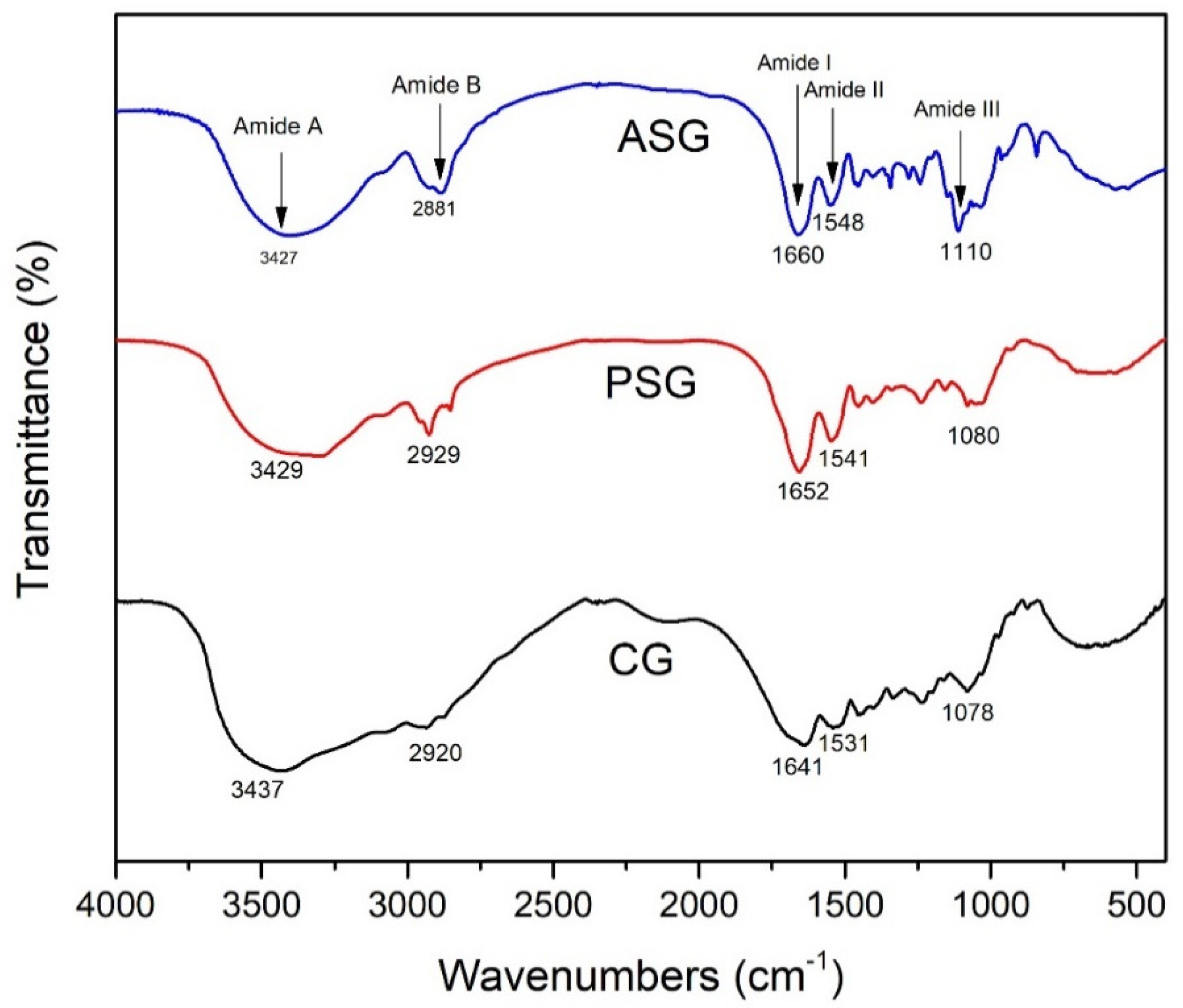
2.1.3. Fourier Transform Infrared (FTIR) Spectroscopy
2.1.4. Scanning Electron Microscopy (SEM) Observations
2.2. In Vitro Biocompatibility of Marine Gelatins
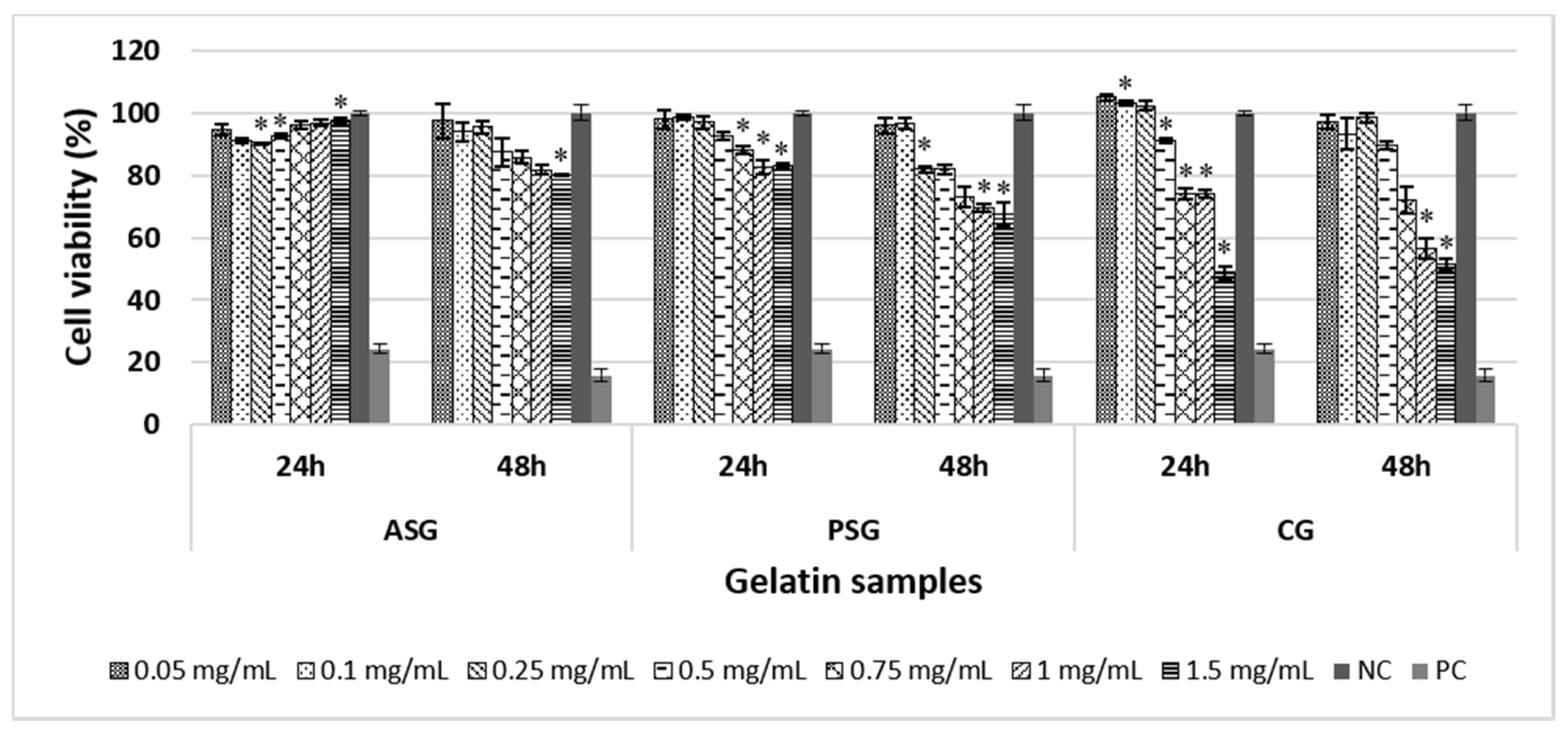
2.2.1. Evaluation of Cell Viability
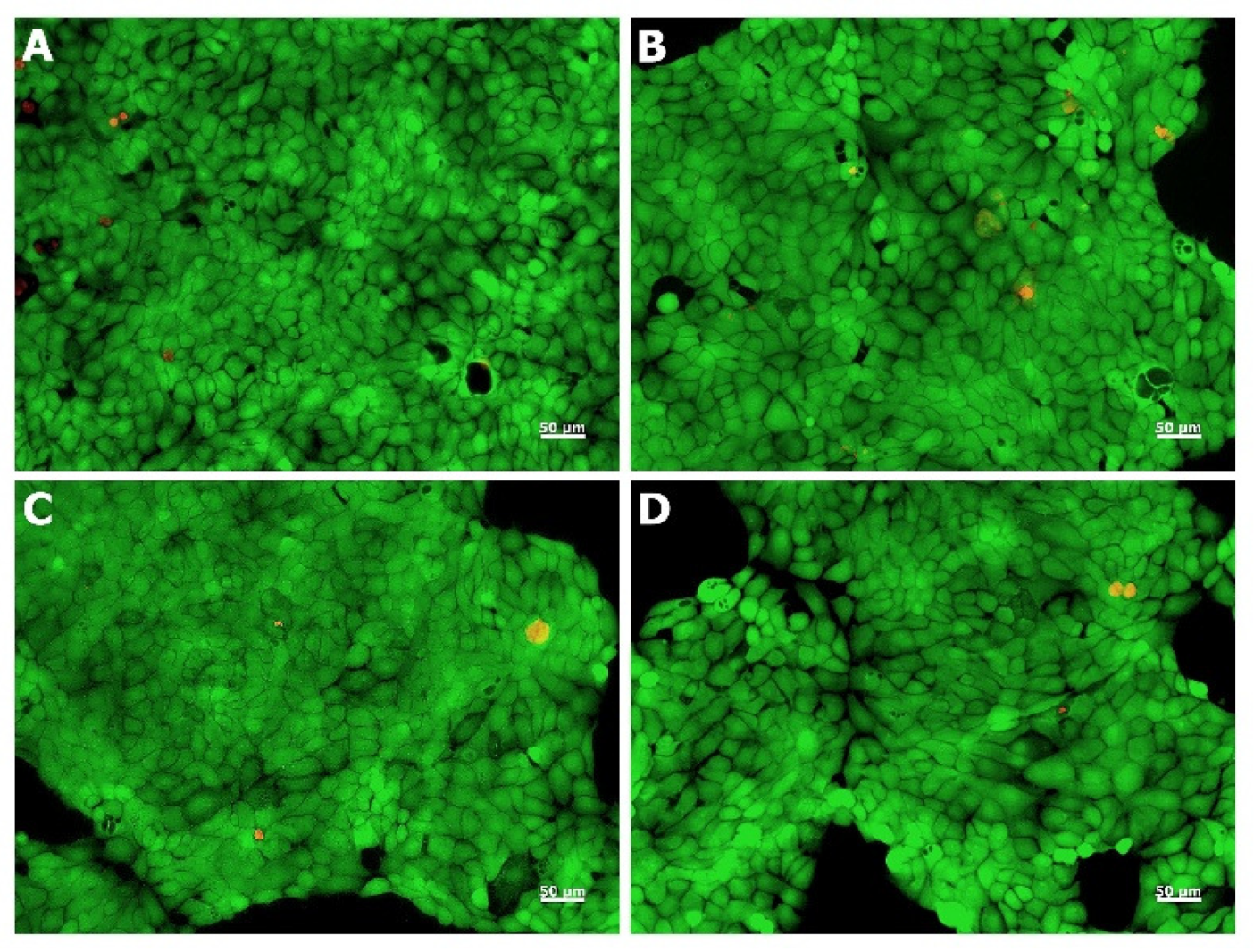
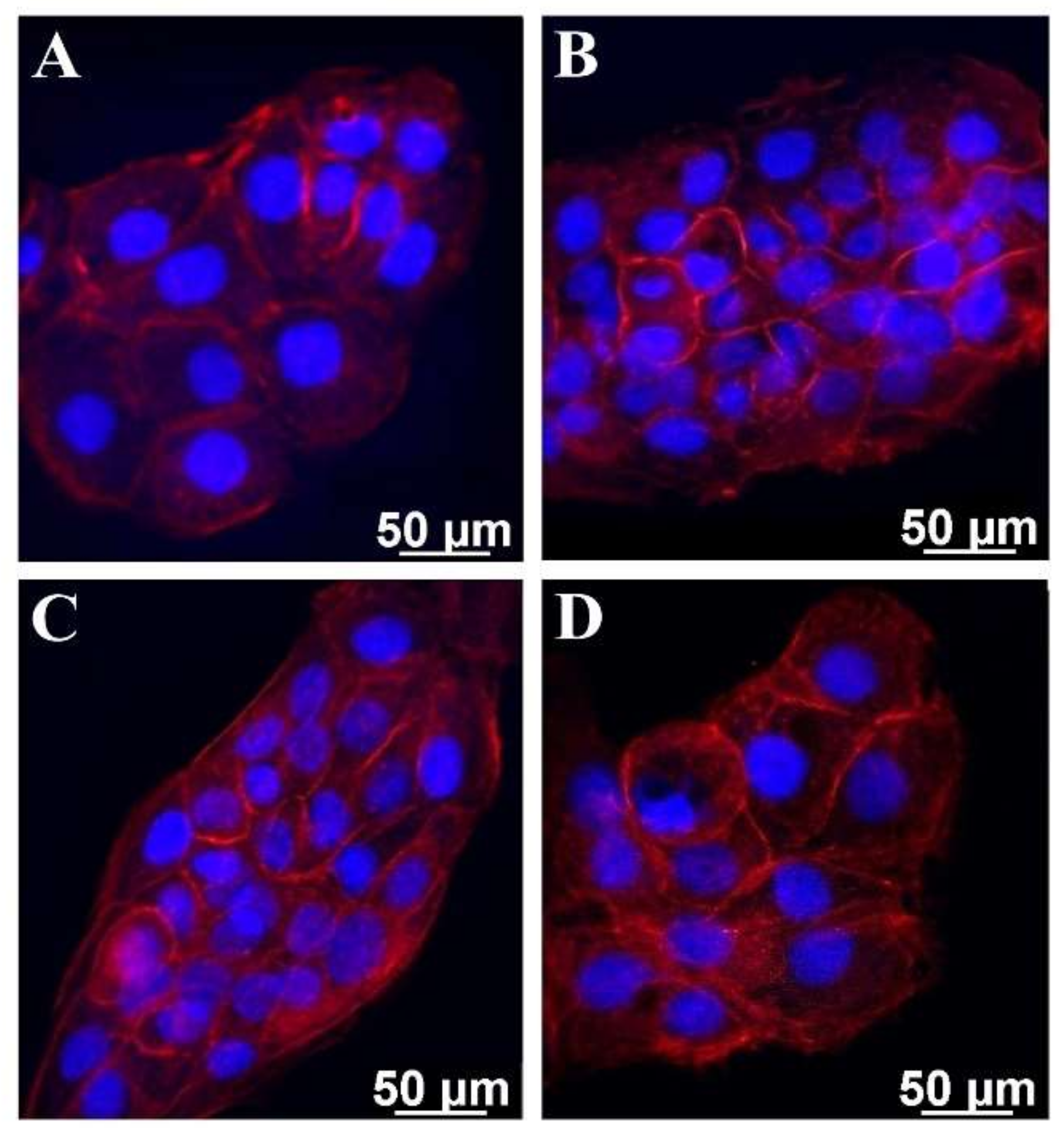
2.2.2. Morphological Examination
2.3. Biological Properties of Marine Gelatins
2.3.1. Cell Adhesion Capacity
2.3.2. Irritant Potential
3. Experimental Section
3.1. Raw Materials
3.2. Gelatin Extraction
3.3. Yield of Gelatin Extraction
3.4. Protein Content
3.5. Hydroxyproline Content
3.6. Differential Scanning Calorimetry
3.7. SDS-PAGE Analysis
3.8. FTIR Spectroscopy
3.9. Scanning Electron Microscopy
3.10. Cell Viability Evaluation
3.11. Live/Dead Assay
3.12. Cell Adhesion Assay
3.13. Irritant Potential Test
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gorgieva, S.; Kokol, V. Collagen- vs. gelatin-based biomaterials and their biocompatibility: Review and perspectives. In Biomaterials Applications for Nanomedicine; Rosario, P., Ed.; InTech: Rijeka, Croatia, 2011; pp. 17–52. ISBN 978-953-307-661-4. [Google Scholar]
- Duconseille, A.; Astruc, T.; Quintana, N.; Meersman, F.; Sante-Lhoutellier, V. Gelatin structure and composition linked to hard capsule dissolution: A review. Food Hydrocoll. 2015, 43, 360–376. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Ann. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- See, S.F.; Ghassem, M.; Mamot, S.; Babji, A.S. Effect of different pretreatments on functional properties of African catfish (Clarias gariepinus) skin gelatin. Int. J. Food Sci. Technol. 2015, 52, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Nikkhah, M.; Akbari, M.; Paul, A.; Memic, A.; Dolatshahi-Pirouz, A.; Khademhosseini, A. Gelatin-based biomaterials for tissue engineering and stem cell bioengineering. In Biomaterials from Nature for Advanced Devices and Therapies; Neves, N.M., Reis, R.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 37–62. ISBN 9781119126218. [Google Scholar]
- Karim, A.A.; Bhat, R. Fish gelatin: Properties, challenges, and prospects as an alternative to mammalian gelatins. Food Hydrocoll. 2009, 23, 563–576. [Google Scholar] [CrossRef]
- Silva, T.; Moreira-Silva, J.; Marques, A.; Domingues, A.; Bayon, Y.; Reis, R. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.A.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Tziveleka, L.A.; Ioannou, E.; Tsiourvas, D.; Berillis, P.; Foufa, E.; Roussis, V. Collagen from the marine sponges Axinella cannabina and Suberites carnosus: Isolation and morphological, biochemical, and biophysical characterization. Mar. Drugs 2017, 15, 152. [Google Scholar] [CrossRef]
- Silva, J.C.; Barros, A.A.; Aroso, I.M.; Fassini, D.; Silva, T.H.; Reis, R.L.; Duarte, A.R.C. Extraction of collagen/gelatin from the marine demosponge Chondrosia reniformis (Nardo, 1847) using water acidified with carbon dioxide–process optimization. Ind. Eng. Chem. Res. 2016, 55, 6922–6930. [Google Scholar] [CrossRef]
- Barros, A.A.; Aroso, I.M.; Silva, T.H.; Mano, J.F.; Duarte, A.R.C.; Reis, R.L. Water and carbon dioxide: Green solvents for the extraction of collagen/gelatin from marine sponges. ACS Sustain. Chem. Eng. 2015, 3, 254–260. [Google Scholar] [CrossRef]
- Chancharern, P.; Laohakunjit, N.; Kerdchoechuen, O.; Thumthanaruk, B. Extraction of type A and type B gelatin from jellyfish (Lobonema smithii). Int. Food Res. J. 2016, 23, 419. [Google Scholar]
- Cho, S.; Ahn, J.R.; Koo, J.S.; Kim, S.B. Physicochemical properties of gelatin from jellyfish Rhopilema hispidum. Fish. Aquat. Sci. 2014, 17, 299–304. [Google Scholar] [CrossRef]
- Chan-Higuera, J.E.; Robles-Sánchez, R.M.; Burgos-Hernández, A.; Márquez-Ríos, E.; Velázquez-Contreras, C.A.; Ezquerra-Brauer, J.M. Squid by-product gelatines: Effect on oxidative stress biomarkers in healthy rats. Czech J. Food Sci. 2016, 34, 105–110. [Google Scholar] [CrossRef]
- Uriarte-Montoya, M.H.; Santacruz-Ortega, H.; Cinco-Moroyoqui, F.J.; Rouzaud-Sández, O.; Plascencia-Jatomea, M.; Ezquerra-Brauer, J.M. Giant squid skin gelatin: Chemical composition and biophysical characterization. Food Res. Int. 2011, 44, 3243–3249. [Google Scholar] [CrossRef]
- Nazeer, R.A.; Suganya, U.S. Porous scaffolds of gelatin from the marine gastropod Ficus variegate with commercial cross linkers for biomedical applications. Food Sci. Biotechnol. 2014, 23, 327–335. [Google Scholar] [CrossRef]
- Zarai, Z.; Balti, R.; Mejdoub, H.; Gargouri, Y.; Sayari, A. Process for extracting gelatin from marine snail (Hexaplex trunculus): Chemical composition and functional properties. Process. Biochem. 2012, 47, 1779–1784. [Google Scholar] [CrossRef]
- Luo, F.; Xing, R.; Wang, X.; Peng, Q.; Li, P. Proximate composition, amino acid and fatty acid profiles of marine snail Rapana venosa meat, visceral mass and operculum. J. Sci. Food Agric. 2017, 97, 5361–5368. [Google Scholar] [CrossRef] [PubMed]
- Badiu, D.L.; Balu, A.M.; Barbes, L.; Luque, R.; Nita, R.; Radu, M.; Rosoiu, N. Physico-chemical characterisation of lipids from Mytilus galloprovincialis (L.) and Rapana venosa and their healing properties on skin burns. Lipids 2008, 43, 829–849. [Google Scholar] [CrossRef]
- Guven, K.C.; Yazic, Z.; Akinci, S.; Okus, E. Fatty acids and sterols of Rapana venosa (Valenciennes, 1846). J. Shellfish Res. 1999, 18, 601–604. [Google Scholar]
- Zolotarev, V. The Black Sea ecosystem changes related to the introduction of new mollusc species. Mar. Ecol. 1996, 17, 227–236. [Google Scholar] [CrossRef]
- Merdzhanova, A.; Panayotova, V.; Dobreva, D.A.; Stancheva, R.; Peycheva, K. Lipid composition of raw and cooked Rapana venosa from the Black Sea. Ovidius Univ. Ann. Chem. 2018, 29, 49–55. [Google Scholar] [CrossRef]
- Badiu, D.L.; Luque, R.; Dumitrescu, E.; Craciun, A.; Dinca, D. Amino acids from Mytilus galloprovincialis (L.) and Rapana venosa molluscs accelerate skin wounds healing via enhancement of dermal and epidermal neoformation. Protein J. 2010, 29, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Xing, R.; Wang, X.; Yang, H.; Li, P. Antioxidant activities of Rapana venosa meat and visceral mass during simulated gastrointestinal digestion and their membrane ultrafiltration fractions. Int. J. Food Sci. Technol. 2018, 53, 395–403. [Google Scholar] [CrossRef]
- Schmidt, M.M.; Dornelles, R.C.P.; Mello, R.O.; Kubota, E.H.; Mazutti, M.A.; Kempka, A.P.; Demiate, I.M. Collagen extraction process. Int. Food Res. J. 2016, 23, 913–922. [Google Scholar]
- Nalinanon, S.; Benjakul, S.; Visessanguan, W.; Kishimura, H. Improvement of gelatin extraction from bigeye snapper skin using pepsin-aided process in combination with protease inhibitor. Food Hydrocoll. 2008, 22, 615–622. [Google Scholar] [CrossRef]
- Shyni, K.; Hema, G.S.; Ninan, G.; Mathew, S.; Joshy, C.G.; Lakshmanan, P.T. Isolation and characterization of gelatin from the skins of skipjack tuna (Katsuwonus pelamis), dog shark (Scoliodon sorrakowah), and rohu (Labeo rohita). Food Hydrocoll. 2014, 39, 68–76. [Google Scholar] [CrossRef]
- Lin, C.C.; Chiou, T.K.; Sung, W.C. Characteristics of gelatin from giant grouper (Epinephelus lanceolatus) skin. Int. J. Food Prop. 2015, 18, 2339–2348. [Google Scholar] [CrossRef]
- Jamilah, B.; Harvinder, K.G. Properties of gelatins from skins of fish—black tilapia (Oreochromis mossambicus) and red tilapia (Oreochromis nilotica). Food Chem. 2002, 77, 81–84. [Google Scholar] [CrossRef]
- Milovanovic, I.; Hayes, M. Marine Gelatine from rest raw materials. Appl. Sci. 2018, 8, 2407. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Effect of extraction temperature on functional properties and antioxidative activities of gelatin from shark skin. Food Bioproc. Tech. 2012, 5, 2646–2654. [Google Scholar] [CrossRef]
- Gudipati, V. Fish gelatin: A versatile ingredient for the food and pharmaceutical industries. In Marine Proteins and Peptides: Biological Activities and Applications; Kim, S., Wijesekara, I., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 271–295. ISBN 9781118375082. [Google Scholar]
- Vallejos, N.; González, G.; Troncoso, E.; Zúñiga, R.N. Acid and enzyme-aided collagen extraction from the byssus of Chilean mussels (Mytilus Chilensis): Effect of process parameters on extraction performance. Food Biophys. 2014, 9, 322–331. [Google Scholar] [CrossRef]
- Grossman, S.; Bergman, M. Process for the Production of Gelatin from Fish Skins. U.S. Patent No 5093474A, 3 March 1992. [Google Scholar]
- Dos Reis, R.L. Method to Obtain Collagen/Gelatin from Marine Sponges. WO Patent WO2015151030A1, 8 October 2015. [Google Scholar]
- Alfaro, A.T.; Biluca, F.C.; Marquetti, C.; Tonial, I.B.; de Souza, N.E. African catfish (Clarias gariepinus) skin gelatin: Extraction optimization and physical–chemical properties. Food Res. Int. 2014, 65, 416–422. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Comparative study on characteristics of gelatin from the skins of brownbanded bamboo shark and blacktip shark as affected by extraction conditions. Food Hydrocoll. 2010, 24, 164–171. [Google Scholar] [CrossRef]
- Sotelo, C.G.; Comesaña, M.B.; Ariza, P.R.; Pérez-Martín, R.I. Characterization of collagen from different discarded fish species of the West coast of the Iberian Peninsula. J. Aquat. Food Prod. T. 2016, 25, 388–399. [Google Scholar] [CrossRef]
- Blanco, M.; Vázquez, J.; Pérez-Martín, R.; Sotelo, C. Hydrolysates of fish skin collagen: An opportunity for valorizing fish industry byproducts. Mar. Drugs 2017, 15, 131. [Google Scholar] [CrossRef]
- Cumming, M.H.; Hall, B.; Hofman, K. Isolation and characterization of major and minor collagens from hyaline cartilage of hoki (Macruronus novaezelandiae). Mar. Drugs 2019, 17, 223. [Google Scholar] [CrossRef]
- Carvalho, A.; Marques, A.; Silva, T.; Reis, R. Evaluation of the potential of collagen from codfish skin as a biomaterial for biomedical applications. Mar. Drugs 2018, 16, 495. [Google Scholar] [CrossRef]
- Lee, K.J.; Park, H.Y.; Kim, Y.K.; Park, J.I.; Yoon, H.D. Biochemical characterization of collagen from the starfish Asterias amurensis. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 221–226. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef]
- Di Paolo, N.C.; Shayakhmetov, D.M. Interleukin 1α and the inflammatory process. Nat. Immunol. 2016, 17, 906. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Zhang, Q.; Dai, T.; Zhang, L.; Zhang, M.; Xiao, X.; Hu, H.; Huang, Y. Identification of potential biomarkers for predicting acute dermal irritation by proteomic analysis. J. Appl. Toxicol. 2011, 31, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Marques, A.; Martins, E.; Silva, T.; Reis, R. Cosmetic potential of marine fish skin collagen. Cosmetics 2017, 4, 39. [Google Scholar] [CrossRef]
- Edwards, C.A.; O’Brien, W.D., Jr. Modified assay for determination of hydroxyproline in a tissue hydrolyzate. Clin. Chim. Acta 1980, 104, 161–167. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gelatin Type | Extraction Yield (%) | Protein Content (%) | Hyp Content (%) | Melting Temperature (°C) |
|---|---|---|---|---|
| ASG | 9.71 ± 0.38 | 91.48 ± 4.61 | 10.62 ± 0.37 | 35.30 ± 1.56 |
| PSG | 8.65 ± 0.42 | 83.12 ± 3.30 | 9.39 ± 0.51 * | 33.20 ± 1.38 |
| CG | - | 86.12 ± 3.23 | 11.17 ± 0.21 | 28.80 ± 1.93 |
| Sample | Tested Concentrations | ||||
|---|---|---|---|---|---|
| 0.1 mg/mL | 0.25 mg/mL | 0.5 mg/mL | 0.75 mg/mL | ||
| ASG | 1.39 ± 0.27 * | 1.10 ± 0.20 * | 3.40 ± 0.55 * | 6.72 ± 1.27 * | |
| PSG | 1.90 ± 0.87 * | 2.82 ± 0.94 * | 5.85 ± 1.83 * | 7.35 ± 1.00 * | |
| CG | 0.37 ± 0.11 * | 0.61 ± 0.18 * | 0.77 ± 0.21 * | 1.06 ± 0.13 * | |
| NC | 0.34 ± 0.07 * | ||||
| PC | 40.93 ± 4.02 | ||||
| Sample | Tested Concentrations | ||||
|---|---|---|---|---|---|
| 0.1 mg/mL | 0.25 mg/mL | 0.5 mg/mL | 0.75 mg/mL | ||
| ASG | 54.08 ± 1.49 * | 162.81 ± 3.03 * | 244.24 ± 19.70 | 279.65 ± 41.15 * | |
| PSG | 92.67 ± 7.55 * | 196.44 ± 6.79 * | 311.89 ± 16.10 * | 384.02 ± 15.64 | |
| CG | 46.46 ± 3.10 * | 30.49 ± 1.59 * | 98.52 ± 1.78 * | 42.53 ± 4.65 * | |
| NC | 6.45 ± 2.30 * | ||||
| PC | 1094.88 ± 188.96 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaspar-Pintiliescu, A.; Stefan, L.M.; Anton, E.D.; Berger, D.; Matei, C.; Negreanu-Pirjol, T.; Moldovan, L. Physicochemical and Biological Properties of Gelatin Extracted from Marine Snail Rapana venosa. Mar. Drugs 2019, 17, 589. https://doi.org/10.3390/md17100589
Gaspar-Pintiliescu A, Stefan LM, Anton ED, Berger D, Matei C, Negreanu-Pirjol T, Moldovan L. Physicochemical and Biological Properties of Gelatin Extracted from Marine Snail Rapana venosa. Marine Drugs. 2019; 17(10):589. https://doi.org/10.3390/md17100589
Chicago/Turabian StyleGaspar-Pintiliescu, Alexandra, Laura Mihaela Stefan, Elena Daniela Anton, Daniela Berger, Cristian Matei, Ticuta Negreanu-Pirjol, and Lucia Moldovan. 2019. "Physicochemical and Biological Properties of Gelatin Extracted from Marine Snail Rapana venosa" Marine Drugs 17, no. 10: 589. https://doi.org/10.3390/md17100589
APA StyleGaspar-Pintiliescu, A., Stefan, L. M., Anton, E. D., Berger, D., Matei, C., Negreanu-Pirjol, T., & Moldovan, L. (2019). Physicochemical and Biological Properties of Gelatin Extracted from Marine Snail Rapana venosa. Marine Drugs, 17(10), 589. https://doi.org/10.3390/md17100589