Actinomycin V Suppresses Human Non-Small-Cell Lung Carcinoma A549 Cells by Inducing G2/M Phase Arrest and Apoptosis via the p53-Dependent Pathway

 and
and
Abstract
1. Introduction
2. Results
2.1. Cytotoxicity of Actinomycin V on Human Non-Small Lung Carcinoma Cells
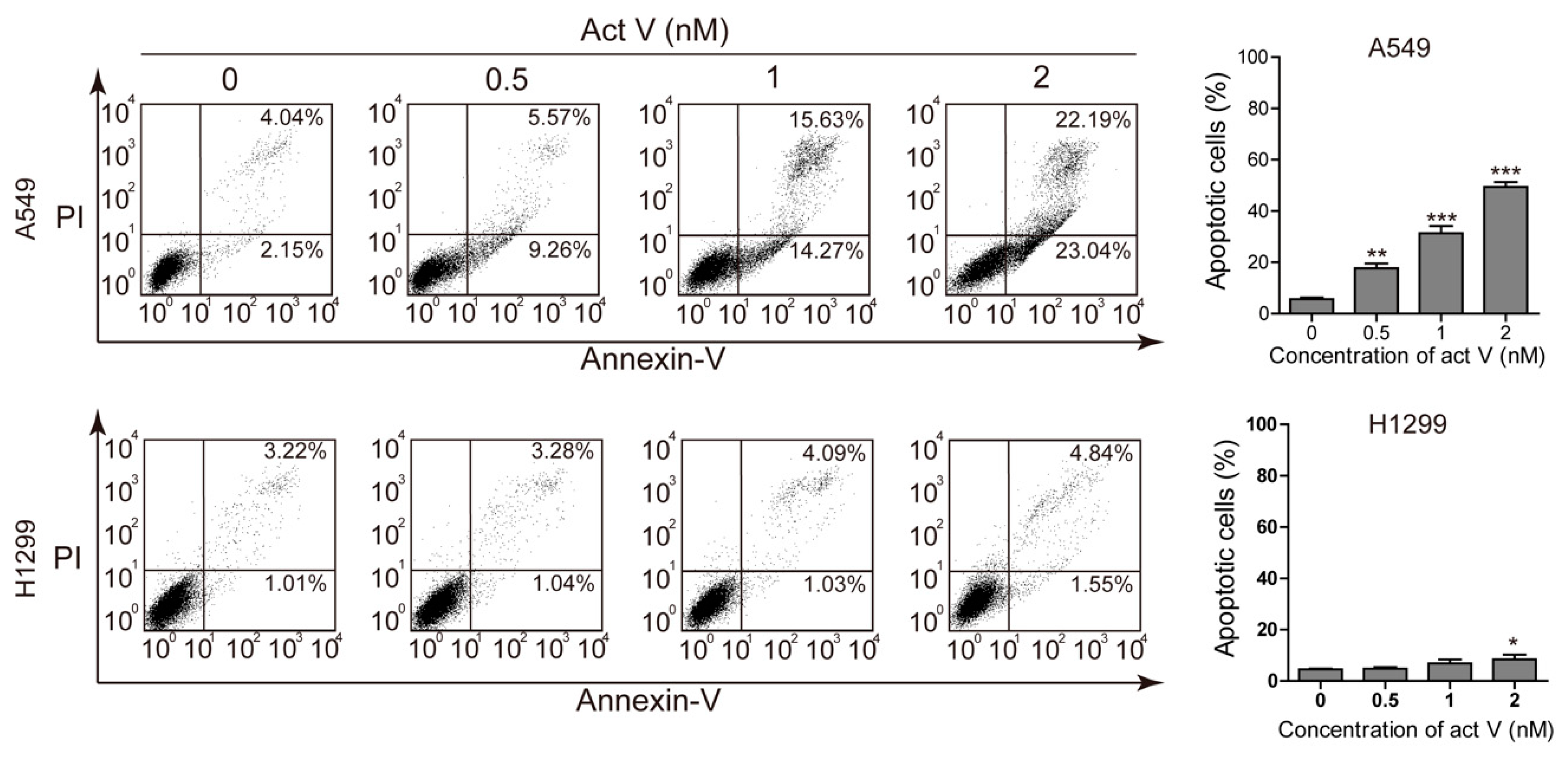
2.2. Actinomycin V Induces Apoptosis in A549 Cells
2.3. Actinomycin V Induces G2/M Phase Arrest in A549 Cells
2.4. Actinomycin V Modulates the Expression of M-Phase-Related Proteins
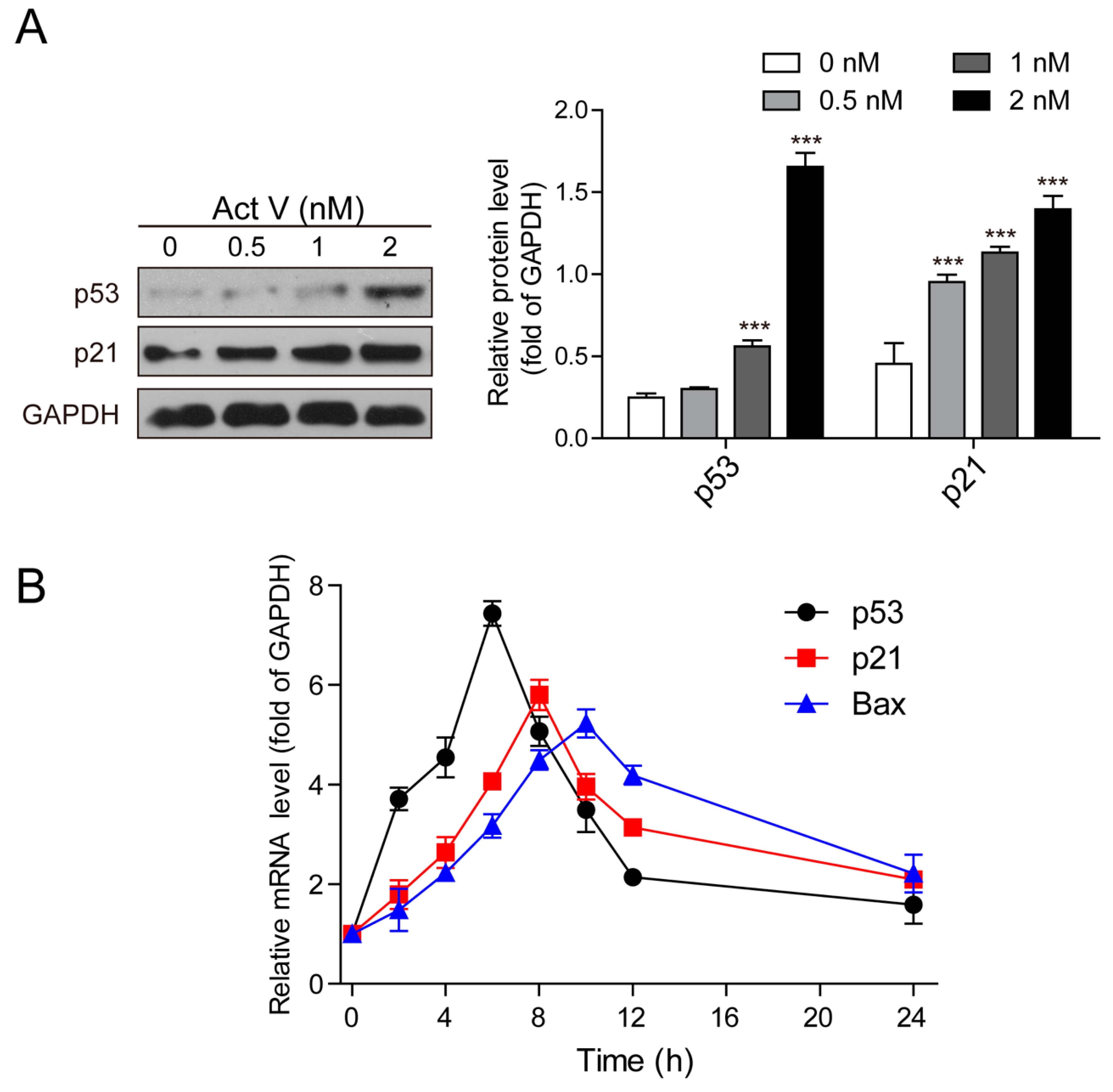
2.5. Actinomycin V Induces the Expression of p53 and p21Waf1/Cip1
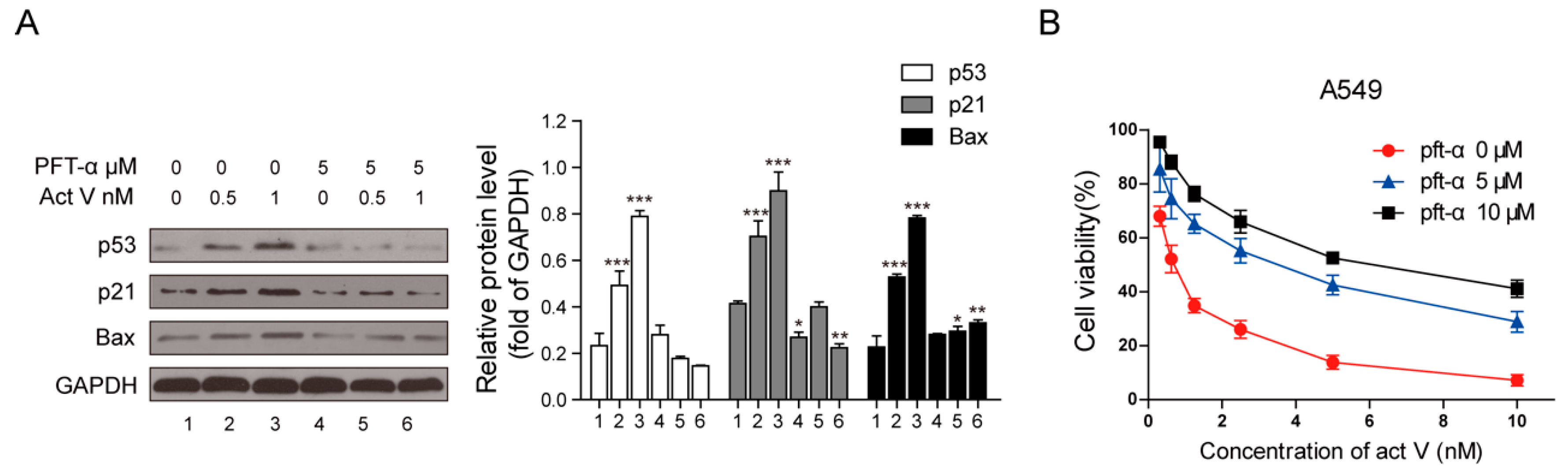
2.6. Inhibition of p53 Blocked Actinomycin V-Induced Cell Cycle Arrest and Apoptosis
2.7. Inhibition of p53 Decreased the Cytotoxicity of Actinomycin V
3. Discussion
4. Materials and Methods
4.1. Reagents, Cell Lines and Cell Culture
4.2. MTT Assay
4.3. DAPI Staining
4.4. Apoptosis Detection via Flow Cytometry
4.5. Cell Cycle Distribution
4.6. Western Blot Analysis
4.7. Real-Time PCR Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Sherr, C.J.; McCormick, F. The RB and p53 pathways in cancer. Cancer Cell 2002, 2, 103–112. [Google Scholar] [CrossRef]
- Hofseth, L.J.; Hussain, S.P.; Harris, C.C. p53: 25 years after its discovery. Trends Pharmacol. Sci. 2004, 25, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Boehme, K.A.; Blattner, C. Regulation of p53-insights into a complex process. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 367–392. [Google Scholar] [CrossRef] [PubMed]
- Soussi, T. The p53 tumor suppressor gene: From molecular biology to clinical investigation. In Colorectal Cancer: New Aspects of Molecular Biology and Immunology and Their Clinical Applications; Hanski, C., Scherubl, H., Mann, B., Eds.; New York Academy of Sciences: New York, NY, USA, 2000; Volume 910, pp. 121–139. [Google Scholar]
- Vassilev, L.T.; Vu, B.T.; Graves, B.; Carvajal, D.; Podlaski, F.; Filipovic, Z.; Kong, N.; Kammlott, U.; Lukacs, C.; Klein, C.; et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science 2004, 303, 844–848. [Google Scholar] [CrossRef]
- Sur, S.; Pagliarini, R.; Bunz, F.; Rago, C.; Diaz, L.A., Jr.; Kinzler, K.W.; Vogelstein, B.; Papadopoulos, N. A panel of isogenic human cancer cells suggests a therapeutic approach for cancers with inactivated p53. Proc. Natl. Acad. Sci. USA 2009, 106, 3964–3969. [Google Scholar] [CrossRef]
- Sobell, H.M. Actinomycin and DNA transcription. Proc. Natl. Acad. Sci. USA 1985, 82, 5328–5331. [Google Scholar] [CrossRef]
- Brockmann, H. Actinomycines. Angew. Chem. Int. Edit. 1960, 72, 939–947. [Google Scholar] [CrossRef]
- Choong, M.L.; Yang, H.; Lee, M.A.; Lane, D.P. Specific activation of the p53 pathway by low dose actinomycin D A new route to p53 based cyclotherapy. Cell Cycle 2009, 8, 2810–2818. [Google Scholar] [CrossRef]
- Zajkowicz, A.; Gdowicz-Klosok, A.; Krzesniak, M.; Scieglinska, D.; Rusin, M. Actinomycin D and nutlin-3a synergistically promote phosphorylation of p53 on serine 46 in cancer cell lines of different origin. Cell. Signal. 2015, 27, 1677–1687. [Google Scholar] [CrossRef]
- Cai, W.; Wang, X.; Elshahawi, S.I.; Ponomareva, L.V.; Liu, X.; McErlean, M.R.; Cui, Z.; Arlinghaus, A.L.; Thorson, J.S.; Van Lanen, S.G. Antibacterial and Cytotoxic Actinomycins Y-6-Y-9 and Zp from Streptomyces sp Strain Go-GS12. J. Nat. Prod. 2016, 79, 2731–2739. [Google Scholar] [CrossRef]
- Morioka, H.; Takezawa, M.; Shibai, H. Actinomycin-V as a potent differentiation inducer of F5-5 friend-leukemia cells. Agric. Biol. Chem. 1985, 49, 2835–2842. [Google Scholar]
- Wang, D.; Wang, C.; Gui, P.; Liu, H.; Khalaf, S.M.H.; Elsayed, E.A.; Wadaan, M.A.M.; Hozzein, W.N.; Zhu, W. Identification, Bioactivity, and Productivity of Actinomycins from the Marine-Derived Streptomyces heliomycini. Front. Microbiol. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Sananes, M.R.; Kapuy, O.; Hunt, T.; Novak, B. Switches and latches: A biochemical tug-of-war between the kinases and phosphatases that control mitosis. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3584–3594. [Google Scholar] [CrossRef] [PubMed]
- Baselga, J.; Swain, S.M. Novel anticancer targets: Revisiting ERBB2 and discovering ERBB3. Nat. Rev. Cancer 2009, 9, 463–475. [Google Scholar] [CrossRef]
- Brown, C.J.; Lain, S.; Verma, C.S.; Fersht, A.R.; Lane, D.P. Awakening guardian angels: Drugging the p53 pathway. Nat. Rev. Cancer 2009, 9, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Jeffers, J.R.; Parganas, E.; Lee, Y.; Yang, C.Y.; Wang, J.L.; Brennan, J.; MacLean, K.H.; Han, J.W.; Chittenden, T.; Ihle, J.N.; et al. Puma is an essential mediator of p53-dependent and -independent apoptotic pathways. Cancer Cell 2003, 4, 321–328. [Google Scholar] [CrossRef]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef]
- Ohki, R.; Nemoto, J.; Murasawa, H.; Oda, E.; Inazawa, J.; Tanaka, N.; Taniguchi, T. Reprimo, a new candidate mediator of the p53-mediated cell cycle arrest at the G(2) phase. J. Biol. Chem. 2000, 275, 22627–22630. [Google Scholar] [CrossRef]
- Sancar, A.; Lindsey-Boltz, L.A.; Unsal-Kacmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef]
- Li, L.; Zheng, B.-B.; Ma, L.-S.; Sun, X.; Chang, J.-J.; Xie, W.-D.; Li, X. Telekin suppresses human hepatocellular carcinoma cells in vitro by inducing G(2)/M phase arrest via the p38 MAPK signaling pathway. Acta Pharmacol. Sin. 2014, 35, 1311–1322. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, P.D.S.; Ornellas, A.A.; Campos, M.R.M.; Scheiner, M.A.M.; Fiedler, W.; Alves, G. Bax and Bcl-2 imbalance and HPB infection in penile tumors and adjacent tissues. Progrès En Urologie 2004, 14, 353–359. [Google Scholar]
- Sun, X.; Li, L.; Ma, H.-G.; Sun, P.; Wang, Q.-L.; Zhang, T.-T.; Shen, Y.-M.; Zhu, W.-M.; Li, X. Bisindolylmaleimide alkaloid BMA-155Cl induces autophagy and apoptosis in human hepatocarcinoma HepG-2 cells through the NF-kappa B p65 pathway. Acta Pharmacol. Sin. 2017, 38, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lin, S.; Zhang, C.; Ma, J.; Han, Z.; Jia, F.; Xie, W.; Li, X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina 2019, 55, 470. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds IC50 (nmol/L) | BEAS-2B | A549 | NCI-H1299 |
|---|---|---|---|
| Actinomycin V | 4.20 ± 0.48 | 0.68 ± 0.06 | 16.37 ± 1.07 |
| Actinomycin D | 16.52 ± 1.03 | 12.84 ± 0.72 | 26.64 ± 1.32 |
| Doxorubicin | 1091.00 ± 11.01 | 941.60 ± 15.37 | 1546.33 ± 41.09 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-q.; Jia, F.-j.; Zhang, C.-y.; Liu, F.-y.; Ma, J.-h.; Han, Z.; Xie, W.-d.; Li, X. Actinomycin V Suppresses Human Non-Small-Cell Lung Carcinoma A549 Cells by Inducing G2/M Phase Arrest and Apoptosis via the p53-Dependent Pathway. Mar. Drugs 2019, 17, 572. https://doi.org/10.3390/md17100572
Lin S-q, Jia F-j, Zhang C-y, Liu F-y, Ma J-h, Han Z, Xie W-d, Li X. Actinomycin V Suppresses Human Non-Small-Cell Lung Carcinoma A549 Cells by Inducing G2/M Phase Arrest and Apoptosis via the p53-Dependent Pathway. Marine Drugs. 2019; 17(10):572. https://doi.org/10.3390/md17100572
Chicago/Turabian StyleLin, Shi-qi, Fu-juan Jia, Cai-yun Zhang, Fang-yuan Liu, Jia-hui Ma, Zhuo Han, Wei-dong Xie, and Xia Li. 2019. "Actinomycin V Suppresses Human Non-Small-Cell Lung Carcinoma A549 Cells by Inducing G2/M Phase Arrest and Apoptosis via the p53-Dependent Pathway" Marine Drugs 17, no. 10: 572. https://doi.org/10.3390/md17100572
APA StyleLin, S.-q., Jia, F.-j., Zhang, C.-y., Liu, F.-y., Ma, J.-h., Han, Z., Xie, W.-d., & Li, X. (2019). Actinomycin V Suppresses Human Non-Small-Cell Lung Carcinoma A549 Cells by Inducing G2/M Phase Arrest and Apoptosis via the p53-Dependent Pathway. Marine Drugs, 17(10), 572. https://doi.org/10.3390/md17100572