A Marine Isolate of Bacillus pumilus Secretes a Pumilacidin Active against Staphylococcus aureus




Abstract
1. Introduction
2. Results and Discussion
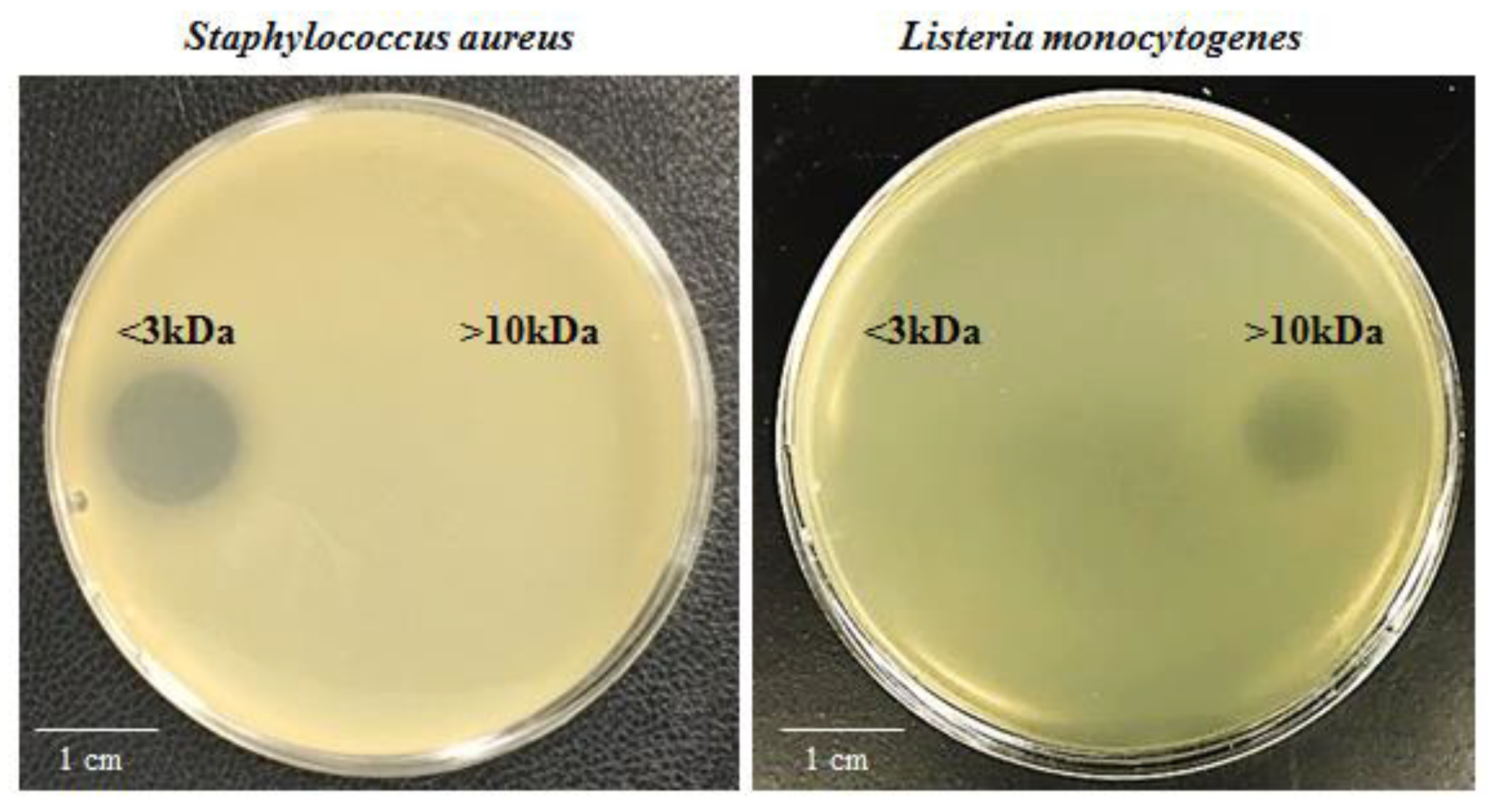
2.1. SF214 Secretes Two Antimicrobials Active Against Staphylococcus aureus or Listeria monocytogenes
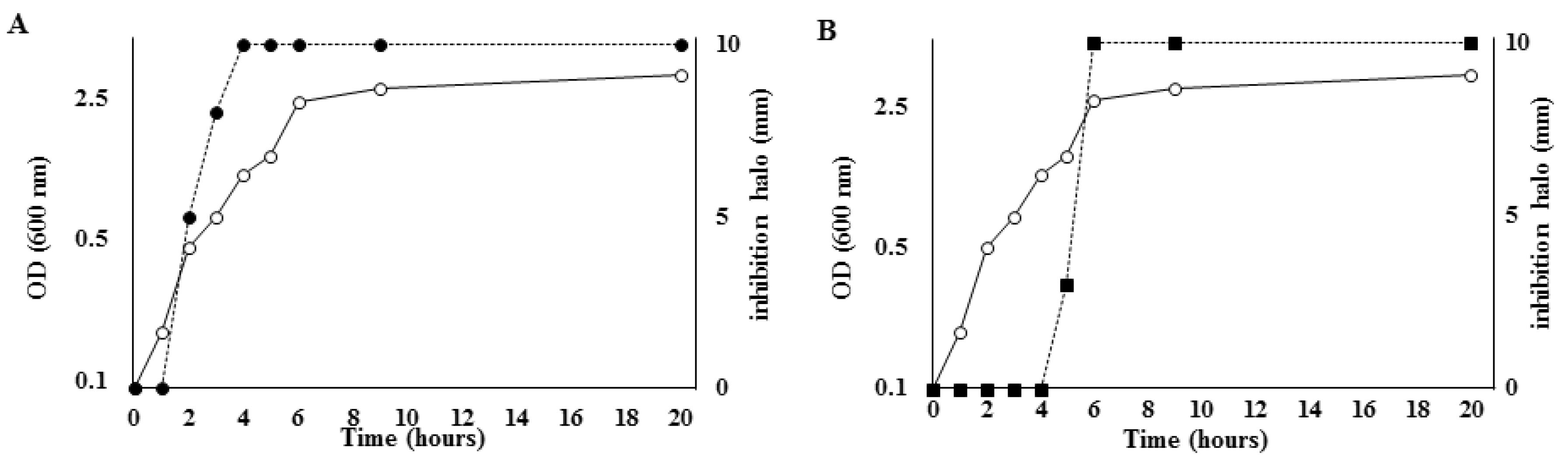
2.2. Production of Both Antimicrobials Is Strictly Regulated
2.3. The Anti-Staphylococcus Molecule Was Stable to Heat, pH, and Treatments with Chemicals and Enzymes
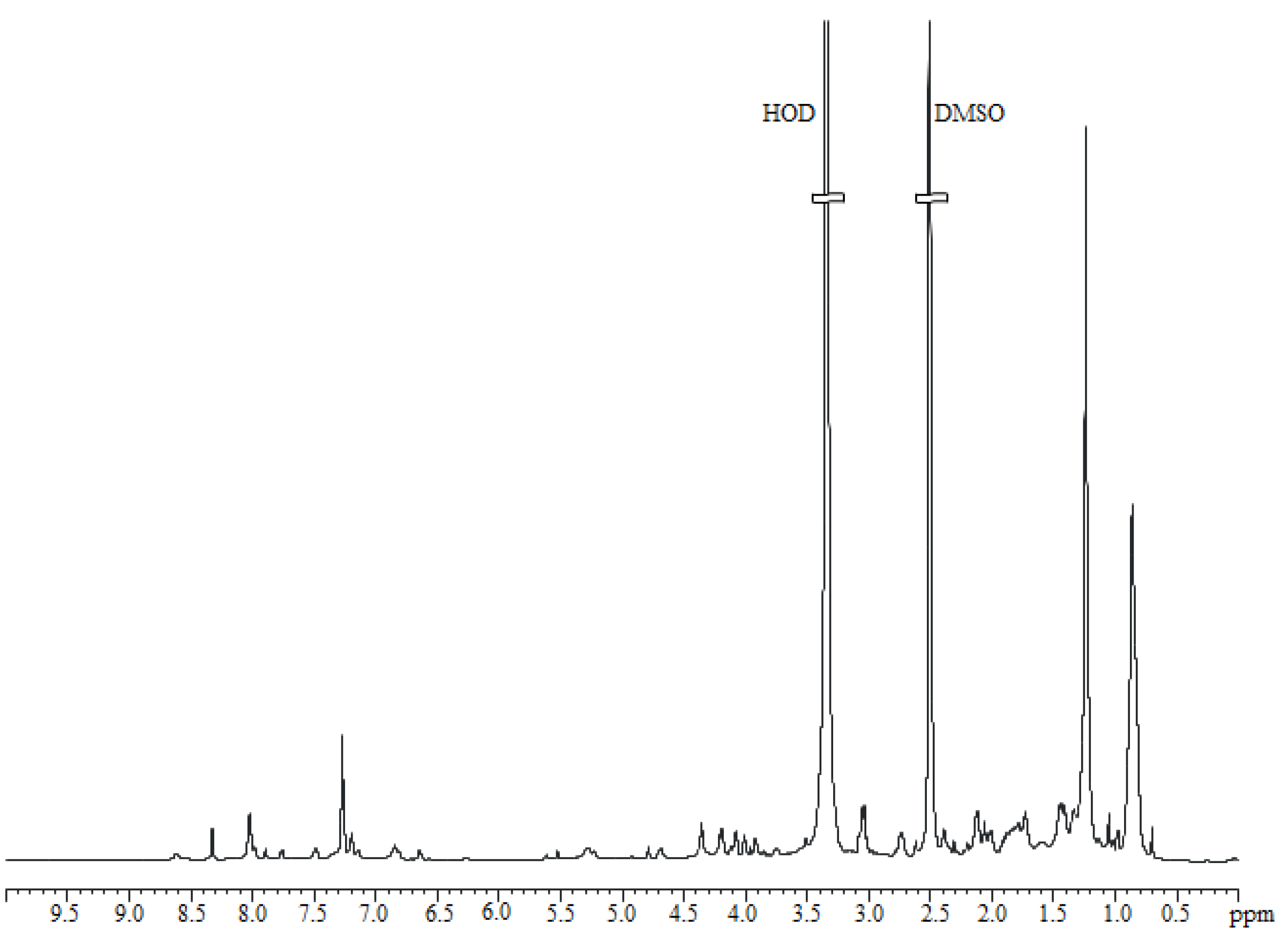
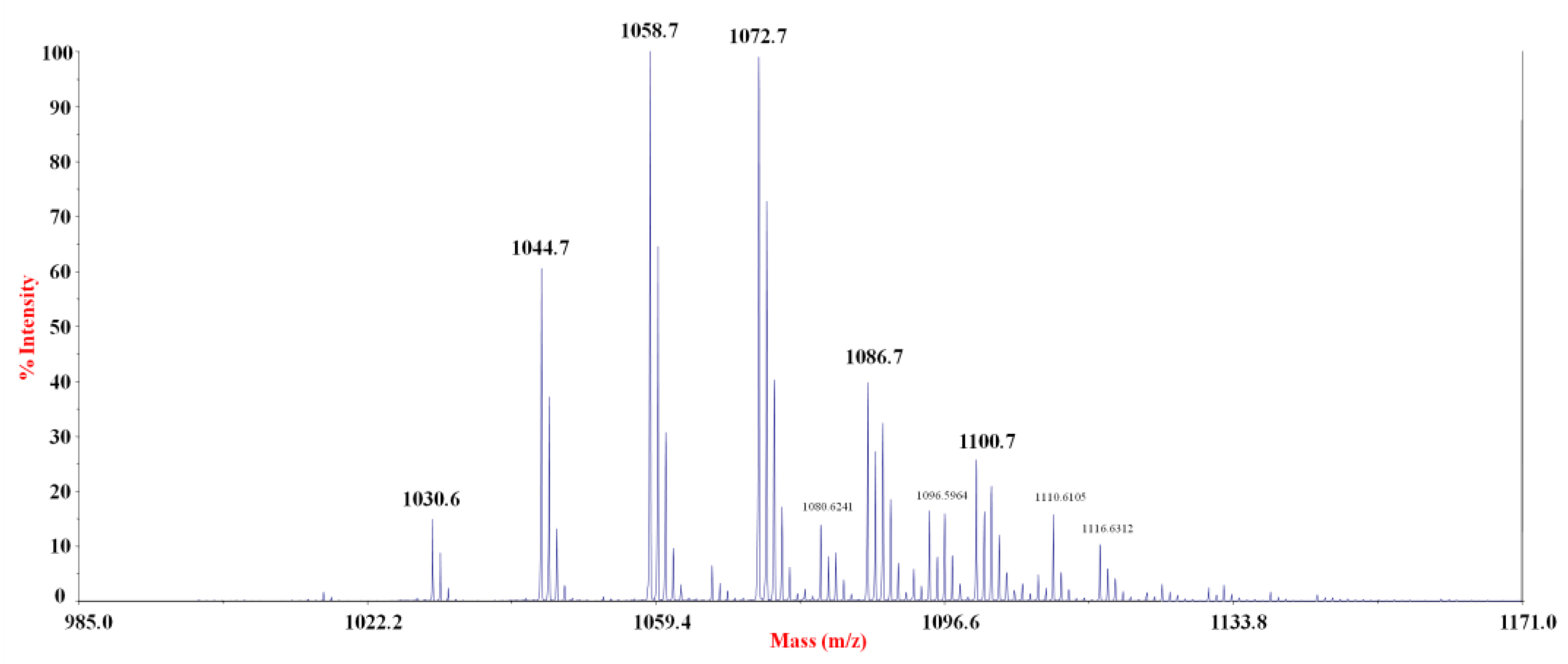
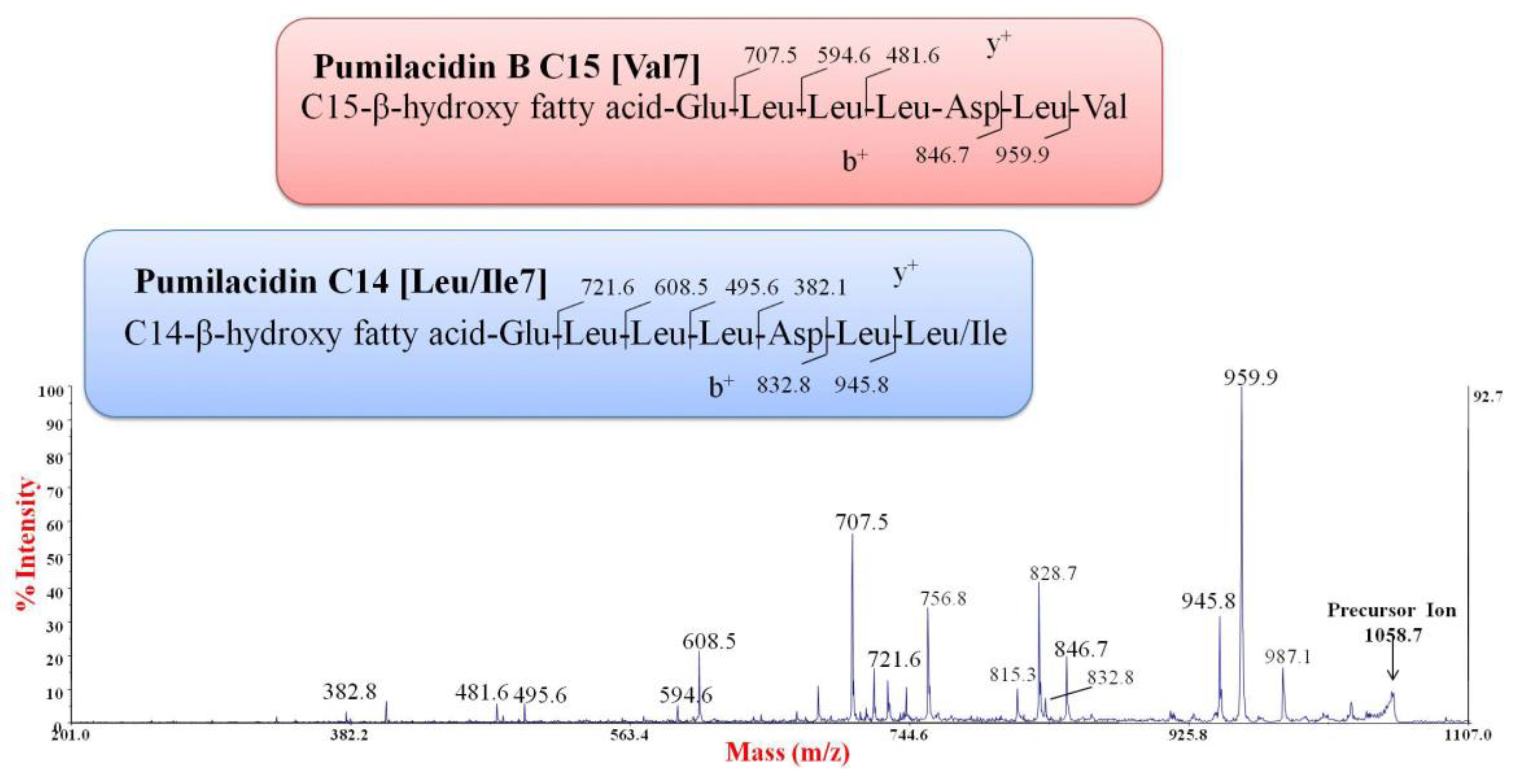
2.4. Purification and Chemical Characterization of the Anti-Staphylococcus Molecule
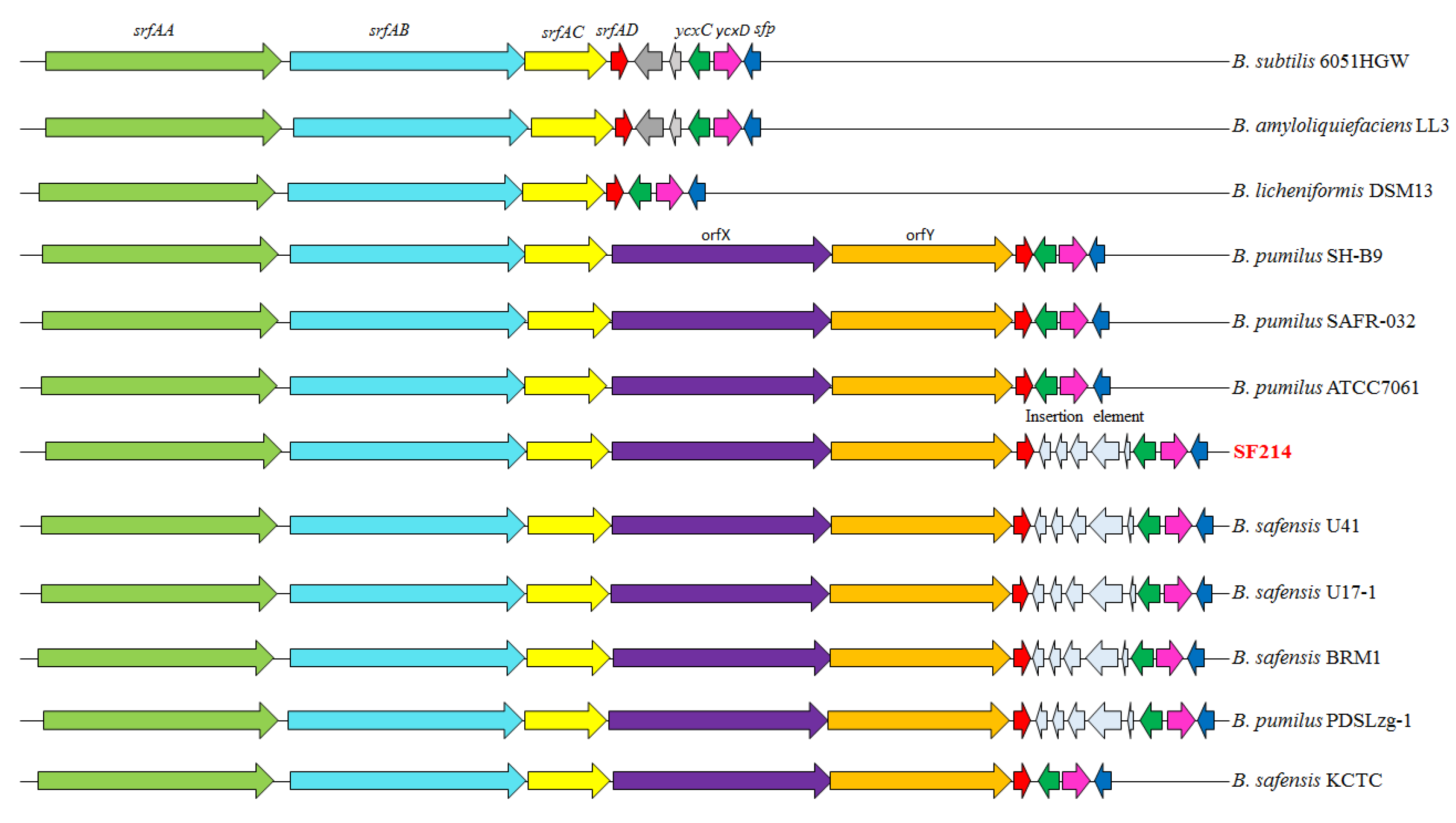
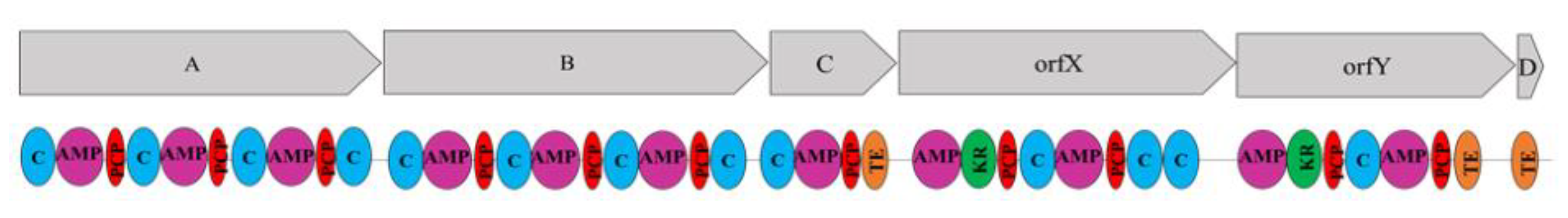
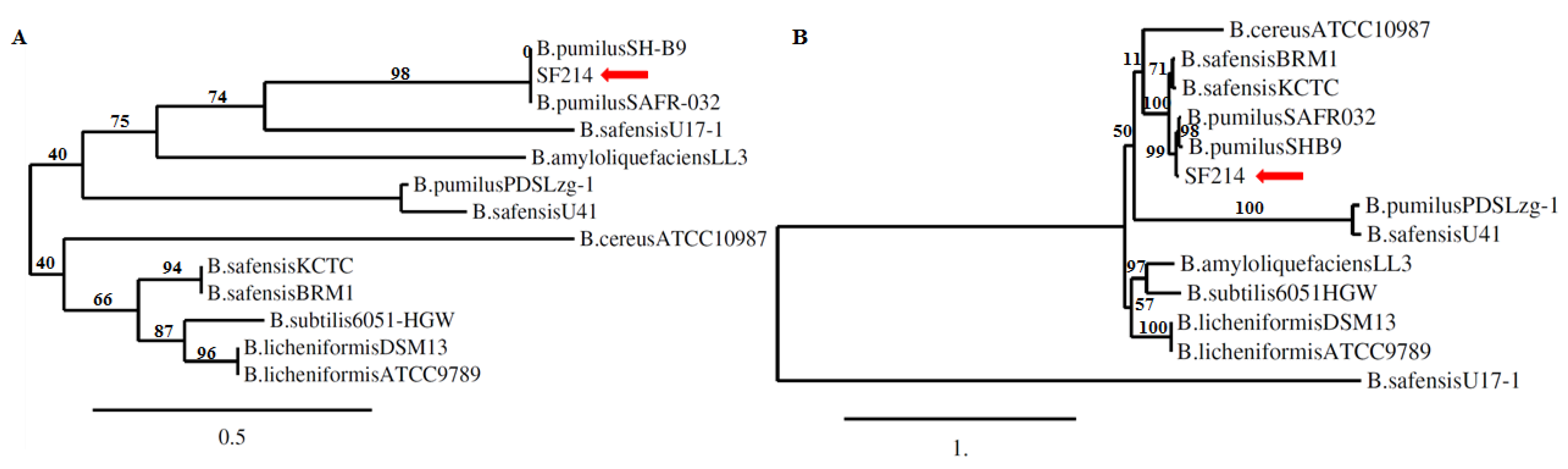
2.5. The srfA-sfp Locus of SF214
3. Experimental Section
3.1. Bacterial Strains
3.2. Filter-Sterilized and Size-Fractionated Supernatants
3.3. Antimicrobial Plate Assay
3.4. Stability of Antimicrobials at Different pH, Temperature, Chemical, and Enzyme Conditions
3.5. Lipopeptide Purification and Preliminary Analysis
3.6. Characterization of Lipopeptide
3.7. Bioinformatic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal Peptides from Marine Microbes and their Antimicrobial and Anticancer Potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bedoya, L.M.; Bermejo, P. Marine Compounds and their Antimicrobial Activities. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2011; pp. 1293–1306. [Google Scholar]
- Zhang, D.J.; Liu, R.F.; Li, Y.G.; Tao, L.M.; Tian, L. Two new antifungal cyclic lipopeptides from Bacillus marinus B-9987. Chem. Pharm. Bull. 2010, 58, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Desjardine, K.; Pereira, A.; Wright, H.; Matainahu, T.; Kelly, M.; Andersen, R.J. Tauramamide, a lipopeptide antibiotic produced in culture by Brevibacillus laterosporus isolated from a marine habitat: Structure elucidation and synthesis. J. Nat. Prod. 2007, 70, 1850–1853. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, K.; Degnes, K.F.; Kemmler, M.; Bredholt, H.; Fjaervik, E.; Klinkenberg, G.; Sletta, H.; Ellingsen, T.E.; Zotcheu, S.B. Production of a new thiopeptide antibiotic, TP-1161, by a marine Nocardiopsis species. Appl. Environ. Microbiol. 2010, 76, 4969–4976. [Google Scholar] [CrossRef] [PubMed]
- Silva-Stenico, M.E.; Souza, C.; Sturion, A.; Keiko, T.; Etchegaray, A.; Possedente, S.; Beraldo, L.A.; Fatima, M. Non-ribosomal peptides produced by Brazilian cyanobacterial isolates with antimicrobial activity. Microbiol. Res. 2011, 166, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Martínez Núñez, M.A.; López, V.E. Nonribosomal peptides synthetases and their applications in industry. Sustain. Chem. Process. 2016, 4, 13. [Google Scholar] [CrossRef]
- Shaligram, N.S.; Singhal, R.S. Surfactin—A Review on biosynthesis, fermentation, purification and applications. Food Technol. Biotechnol. 2010, 48, 119–134. [Google Scholar]
- Liu, X.-Y.; Yang, S.-Z.; Mu, B.-Z. Production and characterization of a C15-surfactin-O-methyl ester by a lipopeptide producing strain Bacillus subtilis HSO121. Process. Biochem. 2009, 44, 1144–1151. [Google Scholar] [CrossRef]
- Snook, M.E.; Mitchell, T.; Hinton, D.M.; Bacon, C.W. Isolation and characterization of leu7-surfactin from the endophytic bacterium Bacillus mojavensis RRC 101, a biocontrol agent for Fusarium verticillioides. J. Agric. Food Chem. 2009, l57, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Inès, M.; Dhouha, G. Lipopeptide surfactants: Production, recovery and pore forming capacity. Peptides 2015, 71, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Domingos, D.F.; De Faria, A.F.; Galaverna, R.S.; Eberlin, M.N.; Greenfield, P.; Zucchi, T.D.; Melo, I.S.; Tran-Dinh, N.; Midgley, D.; De Oliveira, V.M. Genomic and chemical insights into biosurfactant production by the mangrove-derived strain Bacillus safensis CCMA-560. Appl. Genet. Mol. Biotechnol. 2015, 99, 3155–3167. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine Bacillus circulans. J. Appl. Microbiol. 2008, 104, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.H.; Liao, Z.Y.; Wang, W.Y.; Lu, M.F. Evaluation of a lipopeptide biosurfactant from Bacillus natto TK-1 as a potential source of anti-adhesive, antimicrobial and antitumor activities. Braz. J. Microbiol. 2009, 40, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Naruse, N.; Tenmyo, O.; Kobaru, S.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Pumilacidin, a complex of new antiviral antibiotics production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1990, 43, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Xiu, P.; Liu, R.; Zhang, D.; Sun, C. Pumilacidin-Like Lipopeptides Derived from Marine Bacterium Bacillus sp. Strain 176 Suppress the Motility of Vibrio alginolyticus. Appl. Environ. Microbiol. 2017, 83, e00450-17. [Google Scholar] [CrossRef] [PubMed]
- Khaneja, R.; Perez-Fons, L.; Fakhry, S.; Baccigalupi, L.; Steiger, S.; To, E.; Sandmann, G.; Dong, T.C.; Ricca, E.; Fraser, P.D.; et al. Carotenoids Found in Bacillus. J. Appl. Microbiol. 2010, 108, 1889–1902. [Google Scholar] [PubMed]
- Manzo, N.; Di Luccia, B.; Isticato, R.; D’Apuzzo, E.; De Felice, M.; Ricca, E. Pigmentation and sporulation are alternative cell fates in Bacillus pumilus SF214. PLoS ONE 2013, 8, e62093. [Google Scholar] [CrossRef] [PubMed]
- Di Luccia, B.; Riccio, A.; Vanacore, A.; Baccigalupi, L.; Molinaro, A.; Ricca, E. Matrix Production, Pigment Synthesis, and Sporulation in a Marine Isolated Strain of Bacillus pumilus. Mar. Drugs 2015, 13, 6472–6488. [Google Scholar] [CrossRef] [PubMed]
- Naclerio, G.; Ricca, E.; Sacco, M.; De Felice, M. Antimicrobial activity of a newly identified bacteriocin of Bacillus cereus. Appl. Environ. Microbiol. 1993, 59, 4313–4316. [Google Scholar] [PubMed]
- Yang, H.; Li, X.; Li, X.; Yu, H.; Shen, Z. Identification of lipopeptide isoforms by MALDI-TOF-MS/MS based on the simultaneous purification of iturin, fengycin, and surfactin by RP-HPLC. Anal. Bioanal. Chem. 2015, 407, 2529–2542. [Google Scholar] [CrossRef] [PubMed]
- Nawel, J.; Manresa, A.; Rabanal, F.; Ben Ayed, H.; Hmidet, N.; Nasri, M. Structural characterization and identification of cyclic lipopeptides produced by Bacillus methylotrophicus DCS1 strain. J. Chromatogr. B 2017, 1060, 374–386. [Google Scholar]
- Finn, R.D.; Clements, J.; Arndt, J.; Miller, B.L.; Wheeler, F.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Baccigalupi, L.; Di Donato, A.; Parlato, M.; Luongo, D.; Carbone, V.; Rossi, M.; Ricca, E.; De Felice, M. Two small, surface-associated factors mediate adhesion of a food-isolated strain of Lactobacillus fermentum to Caco-2 cells. Res. Microbiol. 2005, 56, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Shillinger, U.; Lucke, F.K. Antibacterial activity of Lactobacillus sake isolated from meat. Appl. Environ. Microbiol. 1989, 55, 1901–1906. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, K.T.; Ramette, A.; Tiedje, J.M. The bacterial species definition in the genomic era. Philos. Trans. R. Soc. Lond B Biol. Sci. 2006, 361, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Brack, C.; Mikolasch, A.; Schlueter, R.; Otto, A.; Becher, D.; Wenger, U.; Albrecht, D.; Riedel, K.; Shauer, F. Antibacterial Metabolites and Bacteriolytic Enzymes Produced by Bacillus pumilus During Bacteriolysis of Arthrobacter citreus. Mar. Biotechnol. 2015, 17, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Kalinovskaya, N.I.; Kuznetosova, T.A.; Ivanova, E.P.; Romanenko, L.A.; Voinov, V.G.; Huth, F.; Laatsch, H. Characterization of Surfactin-like Cyclic Depsipeptides Synthesized by Bacillus pumilus from Ascidian Halocynthia aurantium. Mar. Biotechnol. 2002, 4, 179–188. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Temperature (°C) | Supernatant Activity a > 10 kDa against L. monocytogenes | Supernatant Activity a < 3 kDa against S. aureus |
|---|---|---|---|
| LB (Luria-Bertani) | 25 | 7 | 5 |
| BHI (Brain Heart Infusion) | 25 | 10 | 5 |
| DS (Difco Sporulation) | 25 | 7 | 5 |
| S7 | 25 | 10 | 10 |
| S7 | 30 | 10 | 7 |
| S7 | 37 | 5 | 5 |
| pH | Supernatant Activity a < 3 kDa against S. aureus | |
|---|---|---|
| 1 h | 5 h | |
| 2 | 7 | 7 |
| 4 | 10 | 10 |
| 7 | 10 | 10 |
| 10 | 10 | 10 |
| 13 | 10 | 10 |
| Treatment | Supernatant Activity b < 3 kDa against S. aureus |
|---|---|
| None | 10 |
| Trypsin a | 10 |
| Proteinase K a | 10 |
| DNase a | 10 |
| Ribonuclease A a | 10 |
| Acetone c | 10 |
| Ethyl alcohol c | 10 |
| Chloroform c | 10 |
| Toluene c | 10 |
| Incubation (15 min) at: | |
| 60 °C | 10 |
| 80 °C | 10 |
| 100 °C | 5 |
| Lipopeptide | [M + Na]+ (m/z) | Isoforms |
|---|---|---|
| Pumilacidin | 1030.6 | C13 [Val7] |
| 1044.7 | C14 [Val7], C13 [Leu/Ile7] | |
| 1058.7 | C15 [Val7] (pumilacidin B) a, C14 [Leu/Ile7] | |
| 1072.7 | C16 [Val7] (pumilacidin F/G) a, C15 [Leu/Ile7] (pumilacidin A) a | |
| 1086.7 | C17 [Val7] (pumilacidin D) a, C16 [Leu/Ile7] (pumilacidin E) a | |
| 1100.7 | C18 [Val7], C17 [Leu/Ile7] (pumilacidin C) a |
| Genes | Putative Encoded Protein | Protein Identity (%) a | ||
|---|---|---|---|---|
| B. pumilus SF214 vs. B. pumilus SAFR-032 | B. pumilus SF214 vs. B. safensis U17-1 | B. pumilus SAFR-032 vs. B. safensis U17-1 | ||
| srfAA | surfactin synthase subunit 1 | 95 | 91 | 90 |
| srfAB | surfactin synthase subunit 2 | 94 | 91 | 90 |
| srfAC | surfactin synthase subunit 3 | 95 | 93 | 91 |
| orfX | nonribosomal peptide synthetase | 94 | 91 | 89 |
| orfY | nonribosomal peptide synthetase | 94 | 92 | 90 |
| srfAD | surfactin synthase thioesterase subunit | 94 | 89 | 87 |
| 1 | hypothetical protein | - | 87 | - |
| 2 | hypothetical protein | - | 89 | - |
| 3 | hypothetical protein | - | 90 | - |
| 4 | hypothetical protein | - | 84 | - |
| 5 | hypothetical protein | - | 90 | - |
| ycxC | transporter | 96 | 93 | 92 |
| ycxD | transcriptional regulator | 95 | 93 | 93 |
| sfp | 4′-phosphopantetheinyl transferase | 94 | 88 | 88 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saggese, A.; Culurciello, R.; Casillo, A.; Corsaro, M.M.; Ricca, E.; Baccigalupi, L. A Marine Isolate of Bacillus pumilus Secretes a Pumilacidin Active against Staphylococcus aureus. Mar. Drugs 2018, 16, 180. https://doi.org/10.3390/md16060180
Saggese A, Culurciello R, Casillo A, Corsaro MM, Ricca E, Baccigalupi L. A Marine Isolate of Bacillus pumilus Secretes a Pumilacidin Active against Staphylococcus aureus. Marine Drugs. 2018; 16(6):180. https://doi.org/10.3390/md16060180
Chicago/Turabian StyleSaggese, Anella, Rosanna Culurciello, Angela Casillo, Maria Michela Corsaro, Ezio Ricca, and Loredana Baccigalupi. 2018. "A Marine Isolate of Bacillus pumilus Secretes a Pumilacidin Active against Staphylococcus aureus" Marine Drugs 16, no. 6: 180. https://doi.org/10.3390/md16060180
APA StyleSaggese, A., Culurciello, R., Casillo, A., Corsaro, M. M., Ricca, E., & Baccigalupi, L. (2018). A Marine Isolate of Bacillus pumilus Secretes a Pumilacidin Active against Staphylococcus aureus. Marine Drugs, 16(6), 180. https://doi.org/10.3390/md16060180