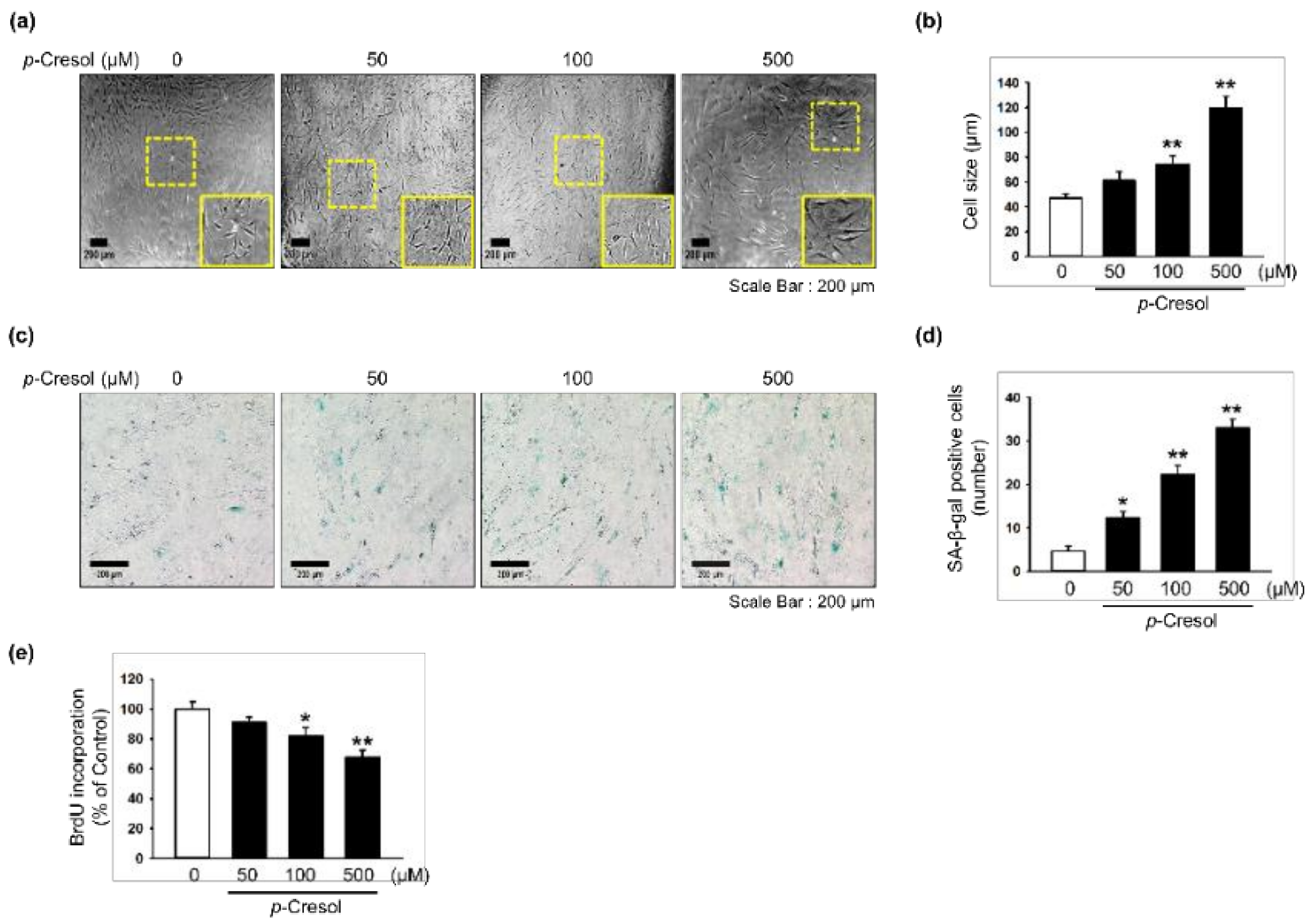
Figure 1.
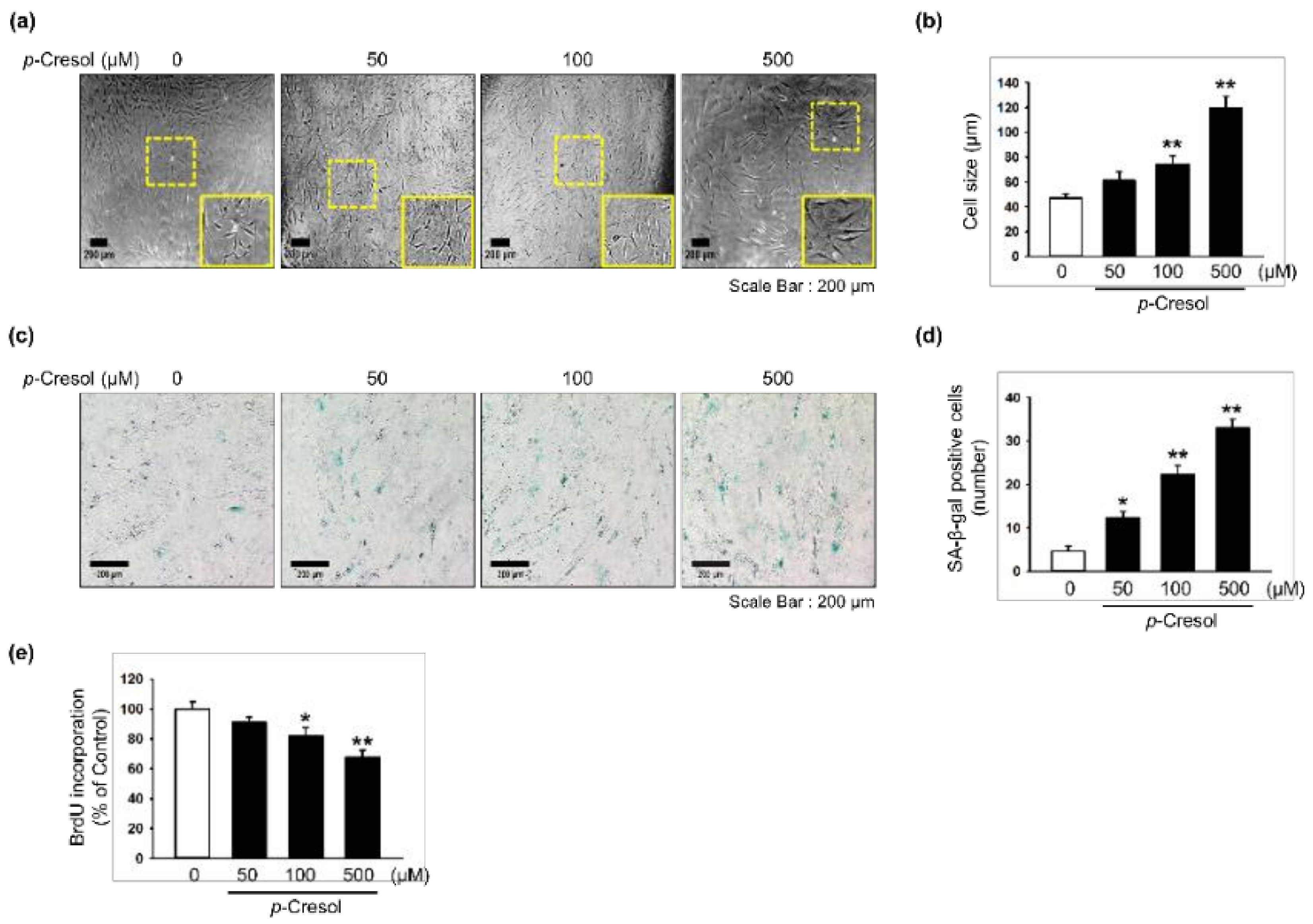
Effect of p-cresol on cellular senescence in MSCs. (a) Morphological changes in MSCs after treatment with p-cresol (50, 100, and 500 μM) for 72 h. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (b) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control. (c) After treatment with p-cresol (50, 100, and 500 μM) for 72 h, senescence was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (d) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 10 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control. (e) After treatment with p-cresol (50, 100 and 500 μM) for 72 h, proliferation was assessed using a BrdU incorporation assay (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control.
Figure 1.
Effect of p-cresol on cellular senescence in MSCs. (a) Morphological changes in MSCs after treatment with p-cresol (50, 100, and 500 μM) for 72 h. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (b) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control. (c) After treatment with p-cresol (50, 100, and 500 μM) for 72 h, senescence was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (d) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 10 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control. (e) After treatment with p-cresol (50, 100 and 500 μM) for 72 h, proliferation was assessed using a BrdU incorporation assay (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control.
![Marinedrugs 16 00121 g001]()
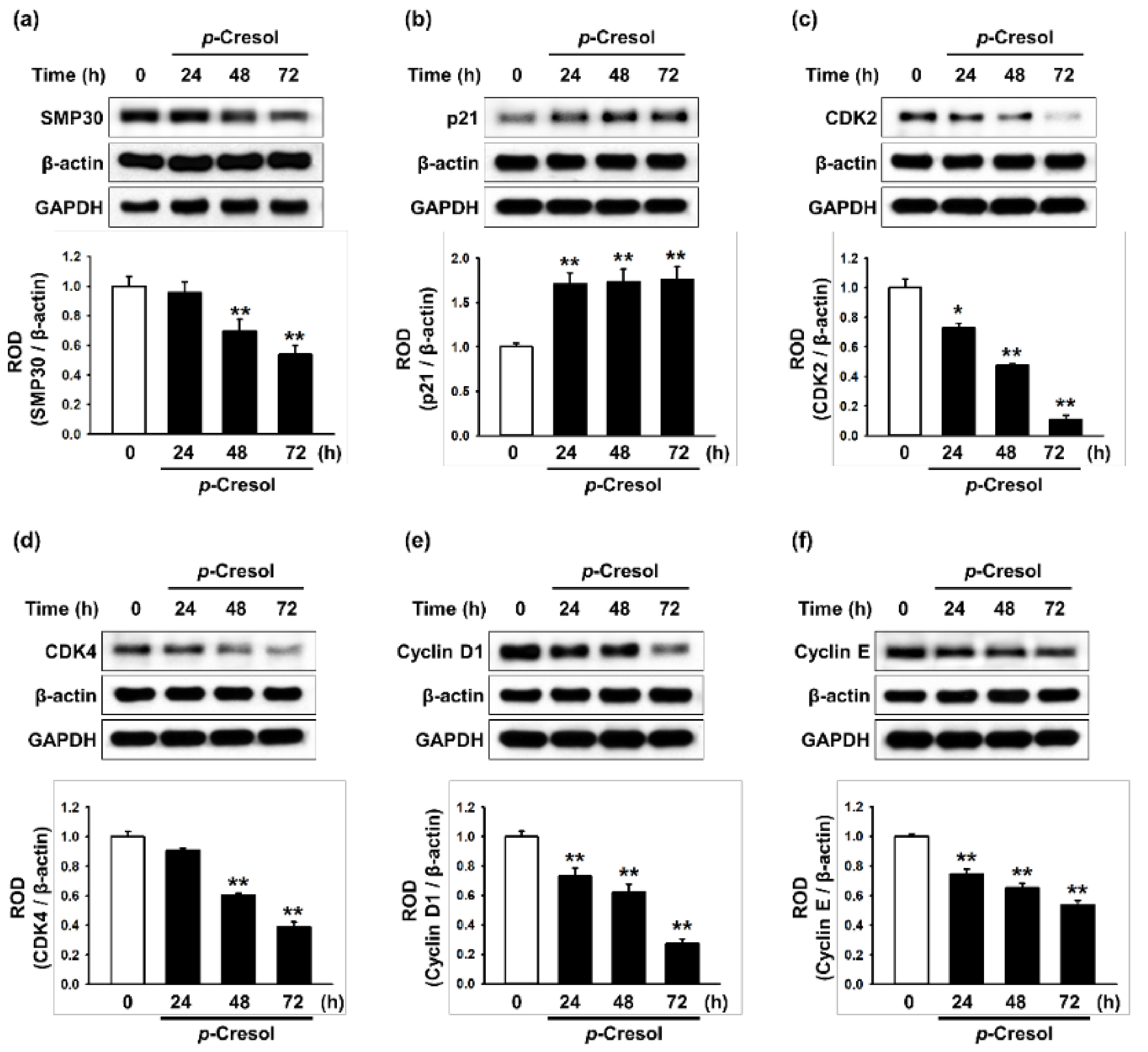
Figure 2.
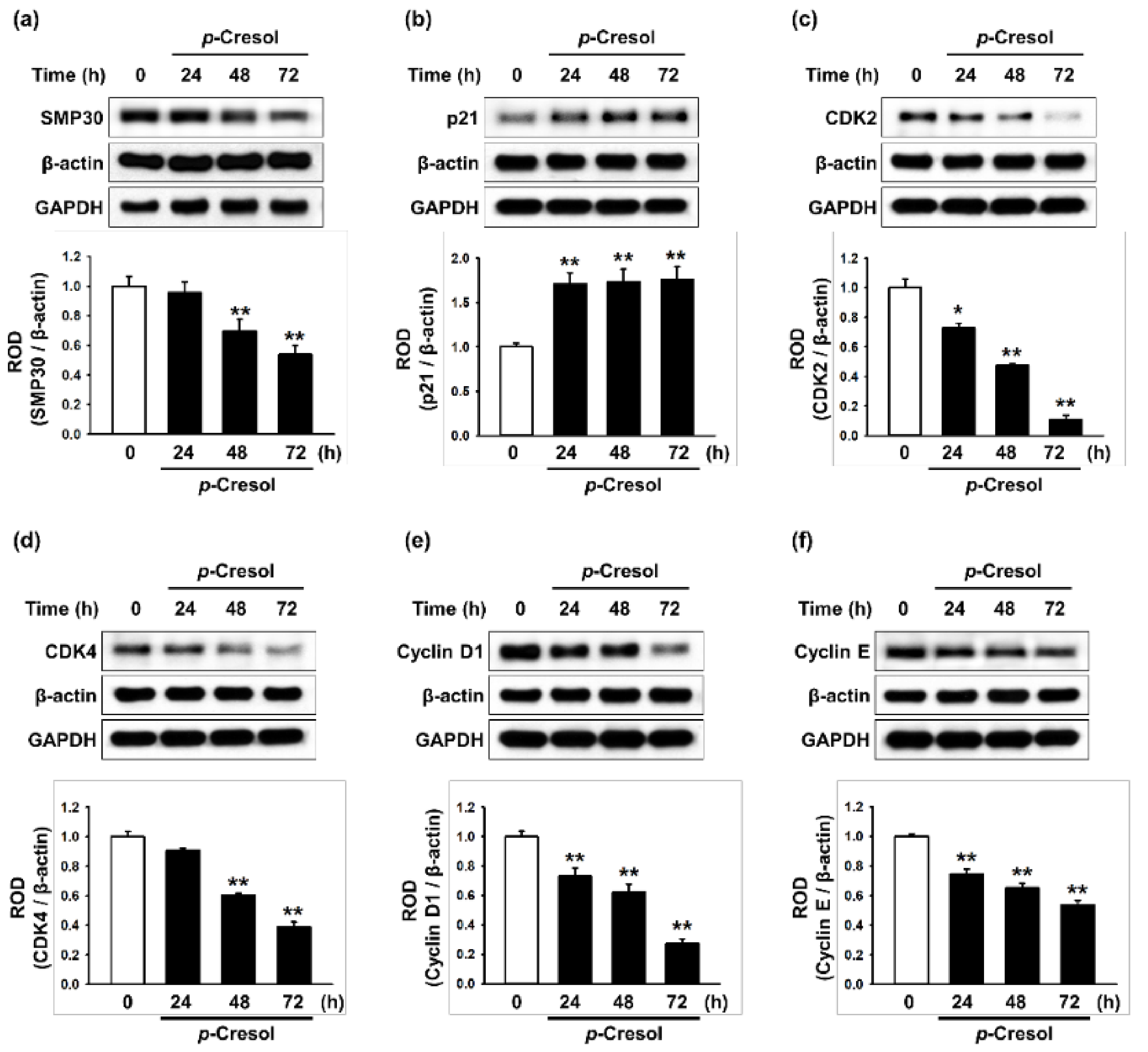
Effect of p-cresol on senescence- and cell cycle-associated protein expression in MSCs. (a–f) After treatment with p-cresol (500 μM) for 0, 24, 48 or 72 h, expression of SMP30 (a), p21 (b), CDK2 (c), CDK4 (d), cyclin D1 (e), and cyclin E (f) was assessed by western blotting. Protein levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control.
Figure 2.
Effect of p-cresol on senescence- and cell cycle-associated protein expression in MSCs. (a–f) After treatment with p-cresol (500 μM) for 0, 24, 48 or 72 h, expression of SMP30 (a), p21 (b), CDK2 (c), CDK4 (d), cyclin D1 (e), and cyclin E (f) was assessed by western blotting. Protein levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control.
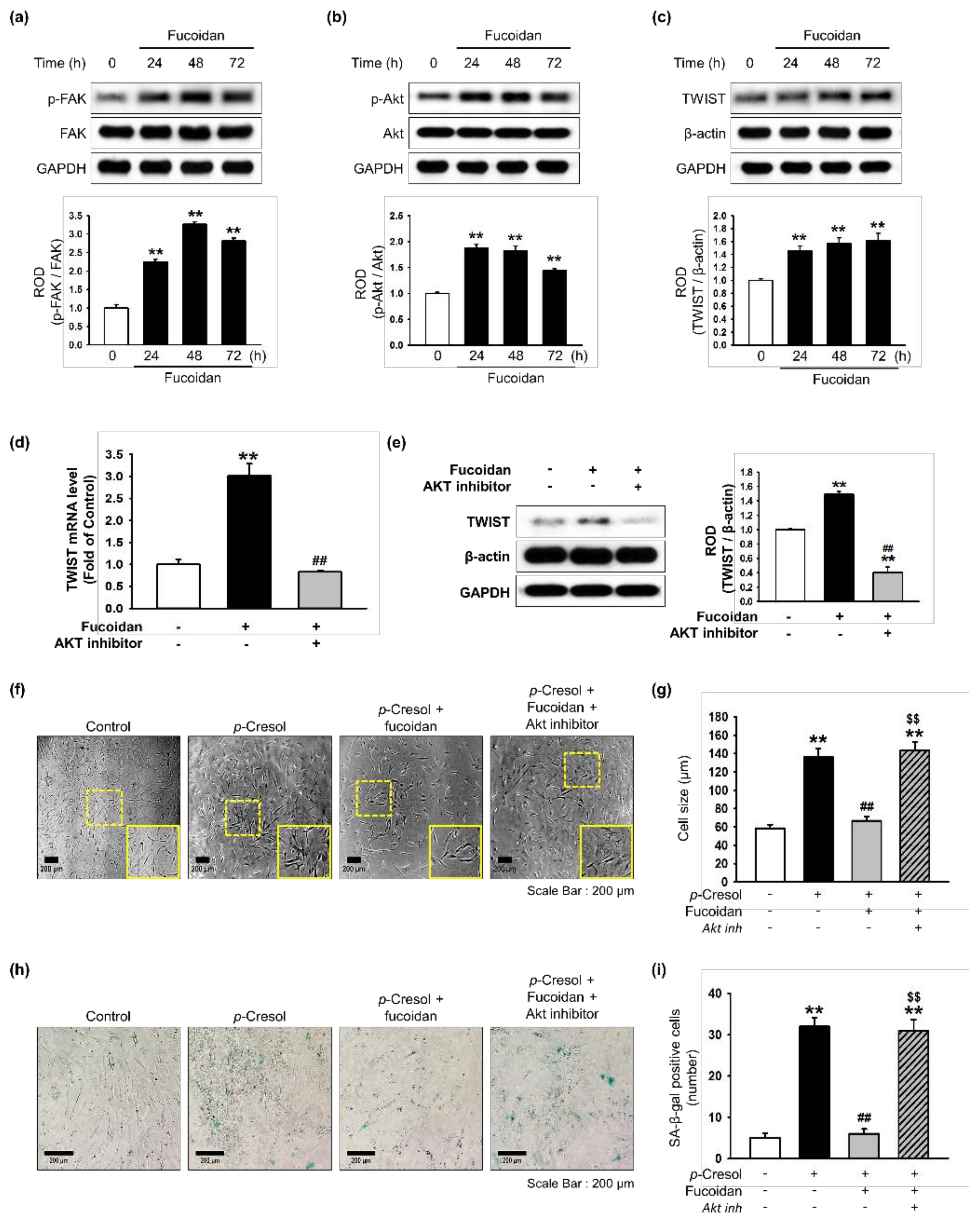
Figure 3.
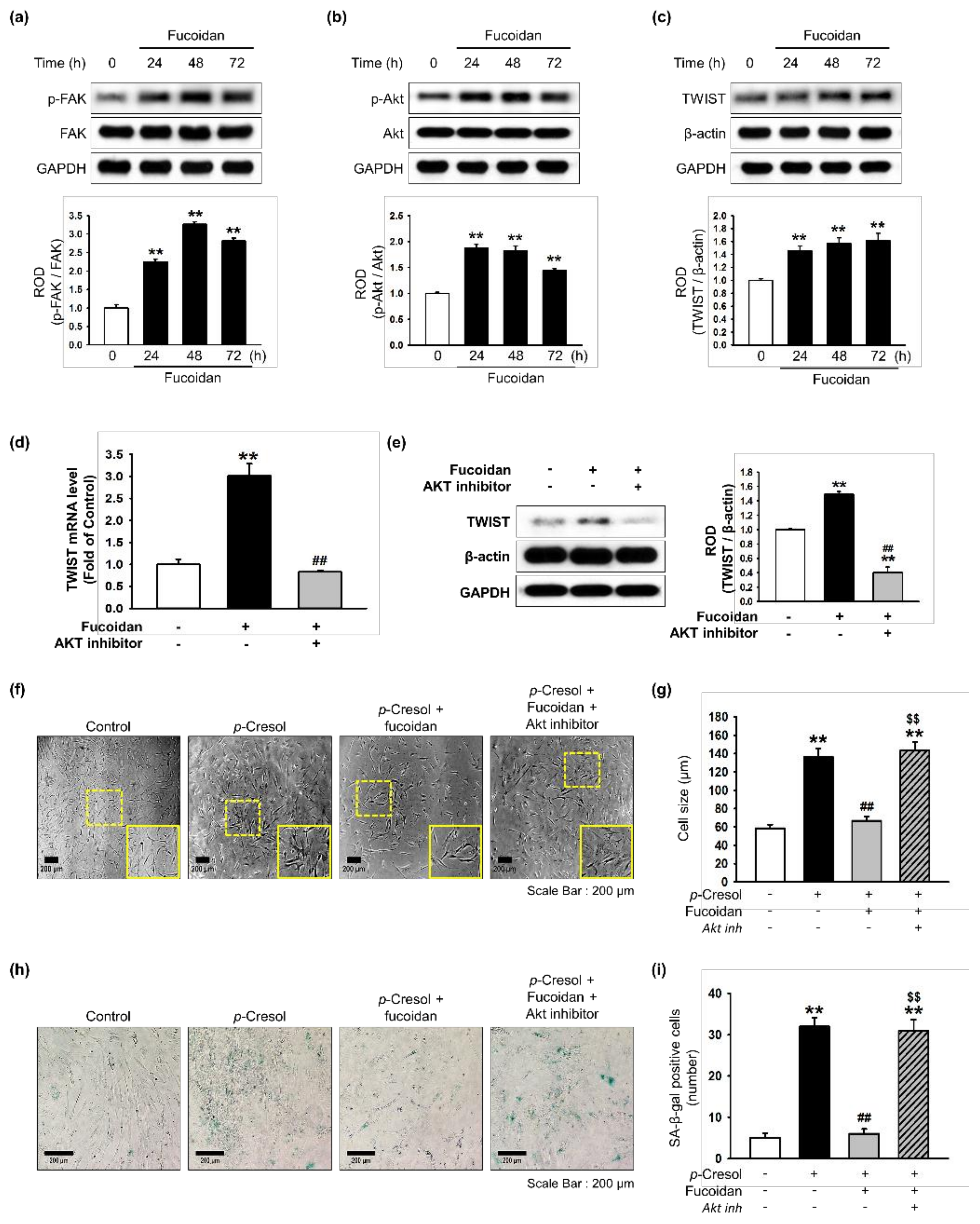
Effect of fucoidan on FAK-Akt-TWIST signaling in MSCs. (a–c) After treatment with fucoidan (10 μg/mL) for 0, 24, 48 or 72 h, expression of p-FAK (a), p-Akt (b), and TWIST (c) was assessed by western blot. Protein levels were quantified by densitometry relative to FAK, Akt and β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control. (d) MSCs were pretreated with an Akt inhibitor (10−6 M) for 30 min before treatment with fucoidan for 72 h; TWIST mRNA level was assessed by qPCR. (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control and ## p < 0.01 vs. treatment with fucoidan alone. (e) MSCs were pretreated with an Akt inhibitor (10−6 M) for 30 min before treatment with fucoidan for 72 h; then TWIST expression was assessed by western blotting. Expression of TWIST was quantified by densitometry relative to β-actin expression (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. treatment with fucoidan alone. (f) Morphological changes after pretreatment with or without Akt inhibitor (10−6 M; 30 min) and fucoidan (10 μg/mL) in MSCs treated with p-cresol (500 μM; 72 h). Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (g) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol alone, $$ p < 0.01 vs. fucoidan pretreatment with p-cresol. (h) Pretreatment with or without Akt inhibitor (10−6 M; 30 min) and fucoidan (10 μg/mL) in MSCs after treatment with p-cresol (500 μM; 72 h, senescence was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (i) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol only, $$ p < 0.01 vs. fucoidan pretreatment with p-cresol.
Figure 3.
Effect of fucoidan on FAK-Akt-TWIST signaling in MSCs. (a–c) After treatment with fucoidan (10 μg/mL) for 0, 24, 48 or 72 h, expression of p-FAK (a), p-Akt (b), and TWIST (c) was assessed by western blot. Protein levels were quantified by densitometry relative to FAK, Akt and β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control. (d) MSCs were pretreated with an Akt inhibitor (10−6 M) for 30 min before treatment with fucoidan for 72 h; TWIST mRNA level was assessed by qPCR. (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control and ## p < 0.01 vs. treatment with fucoidan alone. (e) MSCs were pretreated with an Akt inhibitor (10−6 M) for 30 min before treatment with fucoidan for 72 h; then TWIST expression was assessed by western blotting. Expression of TWIST was quantified by densitometry relative to β-actin expression (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. treatment with fucoidan alone. (f) Morphological changes after pretreatment with or without Akt inhibitor (10−6 M; 30 min) and fucoidan (10 μg/mL) in MSCs treated with p-cresol (500 μM; 72 h). Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (g) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol alone, $$ p < 0.01 vs. fucoidan pretreatment with p-cresol. (h) Pretreatment with or without Akt inhibitor (10−6 M; 30 min) and fucoidan (10 μg/mL) in MSCs after treatment with p-cresol (500 μM; 72 h, senescence was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (i) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol only, $$ p < 0.01 vs. fucoidan pretreatment with p-cresol.
![Marinedrugs 16 00121 g003]()
Figure 4.
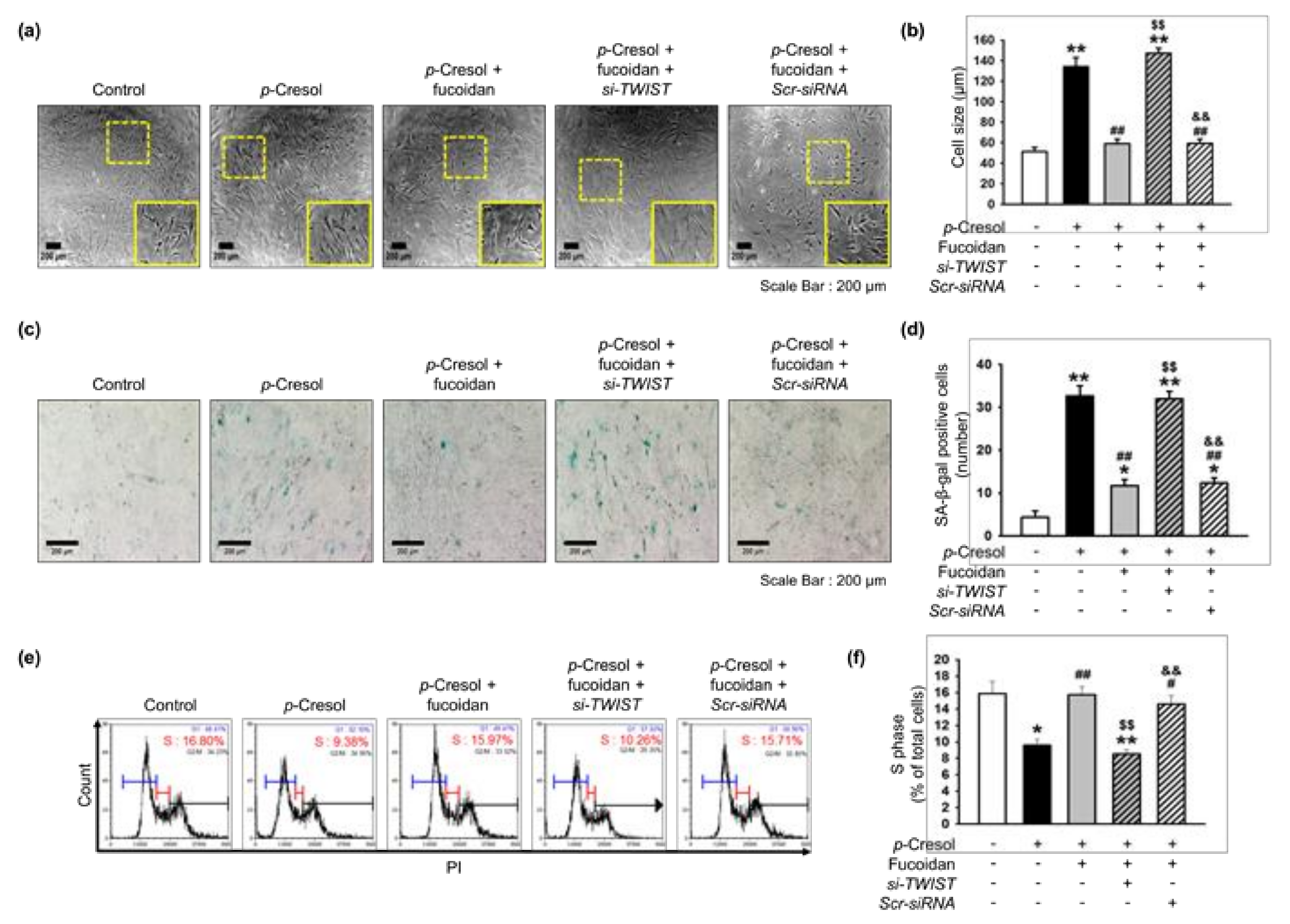
Fucoidan rescues p-cresol-induced cellular senescence in MSCs via the FAK-Akt-TWIST axis. (a) Morphological changes in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with TWIST siRNA (si-TWIST) after treatment with p-cresol (500 μM) for 72 h. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (b) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control (non-treatment), ## p < 0.01 vs. treatment with p-cresol alone (p-cresol), $$ p < 0.01 vs. treatment with p-cresol and fucoidan (p-cresol+fucoidan), && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST (p-cresol+fucoidan+si-TWIST). (c) After p-cresol (500 μM, 72 h) exposure, senescence of MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (d) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 20 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol, $$ p < 0.01 vs. p-cresol+fucoidan, && p < 0.01 vs. p-cresol+fucoidan+si-TWIST. (e) After p-cresol (500 μM, 72 h) exposure, the cell cycle was assessed in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST by flow cytometry with propidium iodide (PI) staining. (f) Proliferation capacity was quantified as the percentage of cells in the S phase (n = 3). Values represent mean ± SEM. * p < 0.05 or ** p < 0.01 vs. control, # p < 0.05 and ## p < 0.01 vs. p-cresol, $$ p < 0.01 vs. p-cresol+fucoidan, && p < 0.01 vs. p-cresol+fucoidan+si-TWIST.
Figure 4.
Fucoidan rescues p-cresol-induced cellular senescence in MSCs via the FAK-Akt-TWIST axis. (a) Morphological changes in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with TWIST siRNA (si-TWIST) after treatment with p-cresol (500 μM) for 72 h. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (b) Determination of cell size (n = 10 images/cultured dishes). Values represent mean ± SEM. ** p < 0.01 vs. control (non-treatment), ## p < 0.01 vs. treatment with p-cresol alone (p-cresol), $$ p < 0.01 vs. treatment with p-cresol and fucoidan (p-cresol+fucoidan), && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST (p-cresol+fucoidan+si-TWIST). (c) After p-cresol (500 μM, 72 h) exposure, senescence of MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by senescence-associated β-galactosidase (SA-β-gal) staining. SA-β-gal positive cells appear blue. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (d) Cellular senescence was quantified as the number of SA-β-gal positive cells (n = 20 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol, $$ p < 0.01 vs. p-cresol+fucoidan, && p < 0.01 vs. p-cresol+fucoidan+si-TWIST. (e) After p-cresol (500 μM, 72 h) exposure, the cell cycle was assessed in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST by flow cytometry with propidium iodide (PI) staining. (f) Proliferation capacity was quantified as the percentage of cells in the S phase (n = 3). Values represent mean ± SEM. * p < 0.05 or ** p < 0.01 vs. control, # p < 0.05 and ## p < 0.01 vs. p-cresol, $$ p < 0.01 vs. p-cresol+fucoidan, && p < 0.01 vs. p-cresol+fucoidan+si-TWIST.
![Marinedrugs 16 00121 g004]()
Figure 5.
Regulation of senescence- and cell cycle-associated proteins in senescent MSCs is dependent on fucoidan-mediated TWIST expression. (a–f) After p-cresol (500 μM, 72 h) exposure, expression of SMP30 (a), p21 (b), CDK2 (c), CDK4 (d), cyclin D1 (e), and cyclin E (f) in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by western blotting. Protein levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control (non-treatment), # p < 0.05 and ## p < 0.01 vs. treatment with p-cresol alone, $ p < 0.05 and $$ p < 0.01 vs. treatment with p-cresol and fucoidan, & p < 0.05 and && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST.
Figure 5.
Regulation of senescence- and cell cycle-associated proteins in senescent MSCs is dependent on fucoidan-mediated TWIST expression. (a–f) After p-cresol (500 μM, 72 h) exposure, expression of SMP30 (a), p21 (b), CDK2 (c), CDK4 (d), cyclin D1 (e), and cyclin E (f) in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by western blotting. Protein levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control (non-treatment), # p < 0.05 and ## p < 0.01 vs. treatment with p-cresol alone, $ p < 0.05 and $$ p < 0.01 vs. treatment with p-cresol and fucoidan, & p < 0.05 and && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST.
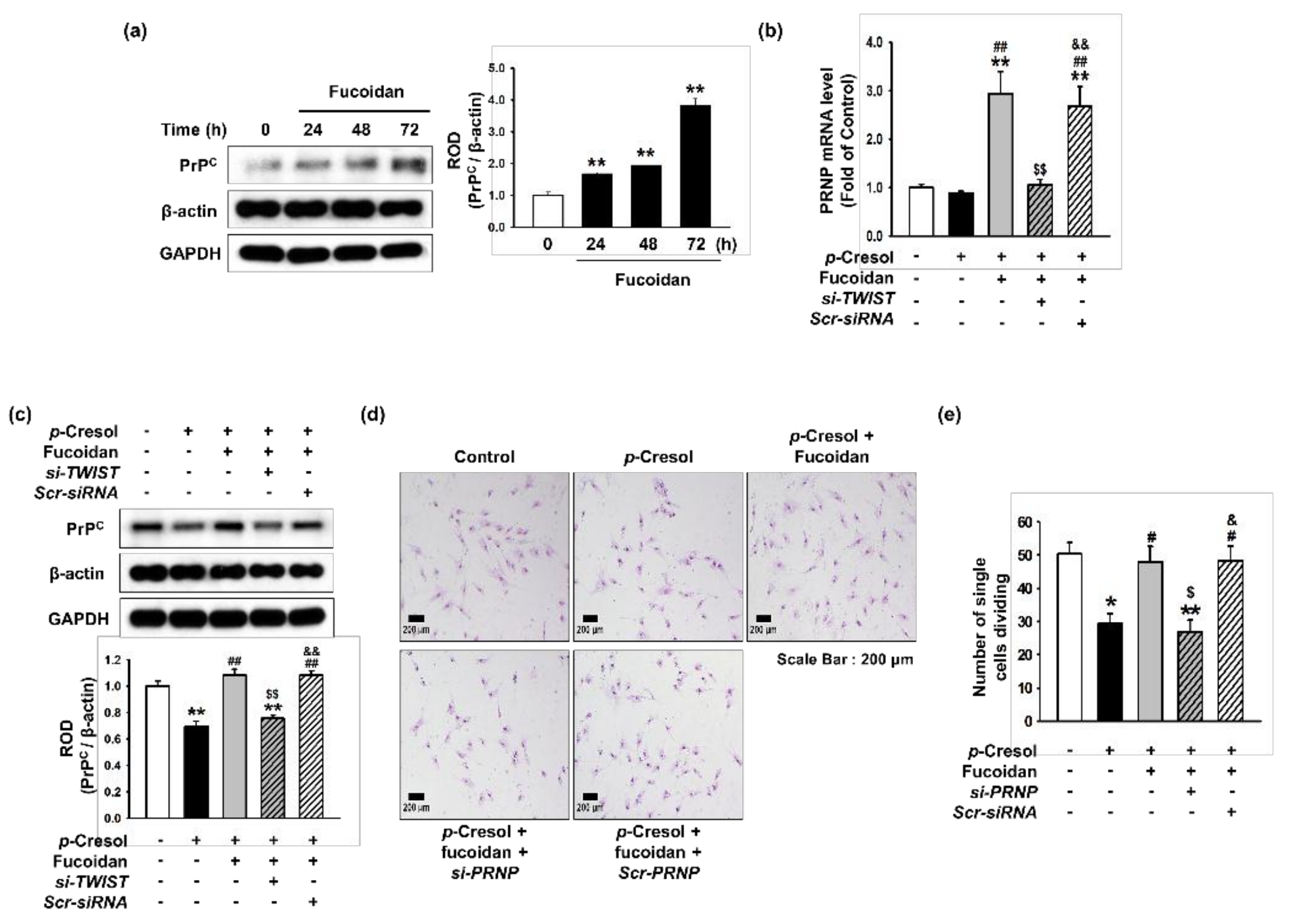
Figure 6.
Effect of fucoidan on cell expansion in p-cresol-induced senescent MSCs through upregulation of PrPC expression. (a) After treatment with fucoidan for 0, 24, 48, or 72 h, expression of PrPC was assessed by western blotting. PrPC levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control. (b) TWIST siRNA- or Scramble siRNA-transfected MSCs pretreated with fucoidan (10 μg/mL) and p-cresol (500 μM; 72 h); then, PRNP mRNA level was assessed by qPCR. (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol alone, $$ p < 0.01 vs. Fucoidan-MSC with p-cresol, && p < 0.01 vs. Fucoidan-si-TWIST transfected MSCs with p-cresol. (c) After p-cresol (500 μM; 72 h) exposure, expression of PrPC in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by western blotting. PrPC levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control (non-treatment), ## p < 0.01 vs. treatment with p-cresol alone, $$ p < 0.01 vs. treatment with p-cresol and fucoidan, && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST. (d) After p-cresol (500 μM, 72 h) exposure, proliferative capacity in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-PRNP was assessed using a single cell expansion assay. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (e) Single cell expansion capacity was quantified as the cell number (n = 10 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control (non-treatment), # p < 0.05 vs. treatment with p-cresol alone, $ p < 0.05 vs. treatment with p-cresol and fucoidan, & p < 0.05 vs. treatment with p-cresol, fucoidan, and si-PRNP.
Figure 6.
Effect of fucoidan on cell expansion in p-cresol-induced senescent MSCs through upregulation of PrPC expression. (a) After treatment with fucoidan for 0, 24, 48, or 72 h, expression of PrPC was assessed by western blotting. PrPC levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control. (b) TWIST siRNA- or Scramble siRNA-transfected MSCs pretreated with fucoidan (10 μg/mL) and p-cresol (500 μM; 72 h); then, PRNP mRNA level was assessed by qPCR. (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control, ## p < 0.01 vs. p-cresol alone, $$ p < 0.01 vs. Fucoidan-MSC with p-cresol, && p < 0.01 vs. Fucoidan-si-TWIST transfected MSCs with p-cresol. (c) After p-cresol (500 μM; 72 h) exposure, expression of PrPC in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-TWIST was assessed by western blotting. PrPC levels were quantified by densitometry relative to β-actin levels (n = 3). Values represent mean ± SEM. ** p < 0.01 vs. control (non-treatment), ## p < 0.01 vs. treatment with p-cresol alone, $$ p < 0.01 vs. treatment with p-cresol and fucoidan, && p < 0.01 vs. treatment with p-cresol, fucoidan, and si-TWIST. (d) After p-cresol (500 μM, 72 h) exposure, proliferative capacity in MSCs, fucoidan-treated MSCs, and fucoidan-treated MSCs pretreated with si-PRNP was assessed using a single cell expansion assay. Representative images are shown from one out of three independent experiments. Scale bar = 200 μm. (e) Single cell expansion capacity was quantified as the cell number (n = 10 images/cultured dishes). Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. control (non-treatment), # p < 0.05 vs. treatment with p-cresol alone, $ p < 0.05 vs. treatment with p-cresol and fucoidan, & p < 0.05 vs. treatment with p-cresol, fucoidan, and si-PRNP.
![Marinedrugs 16 00121 g006]()
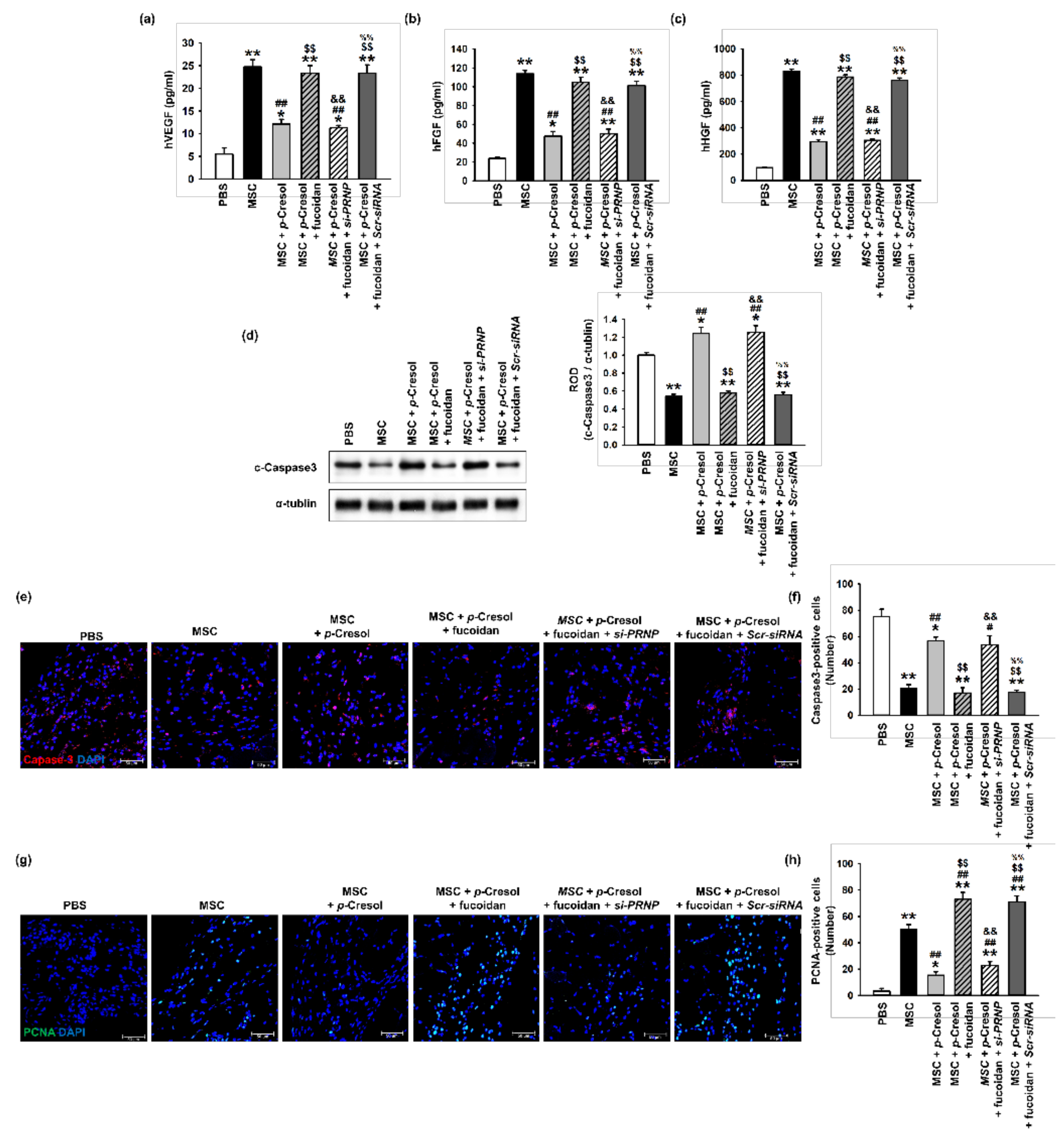
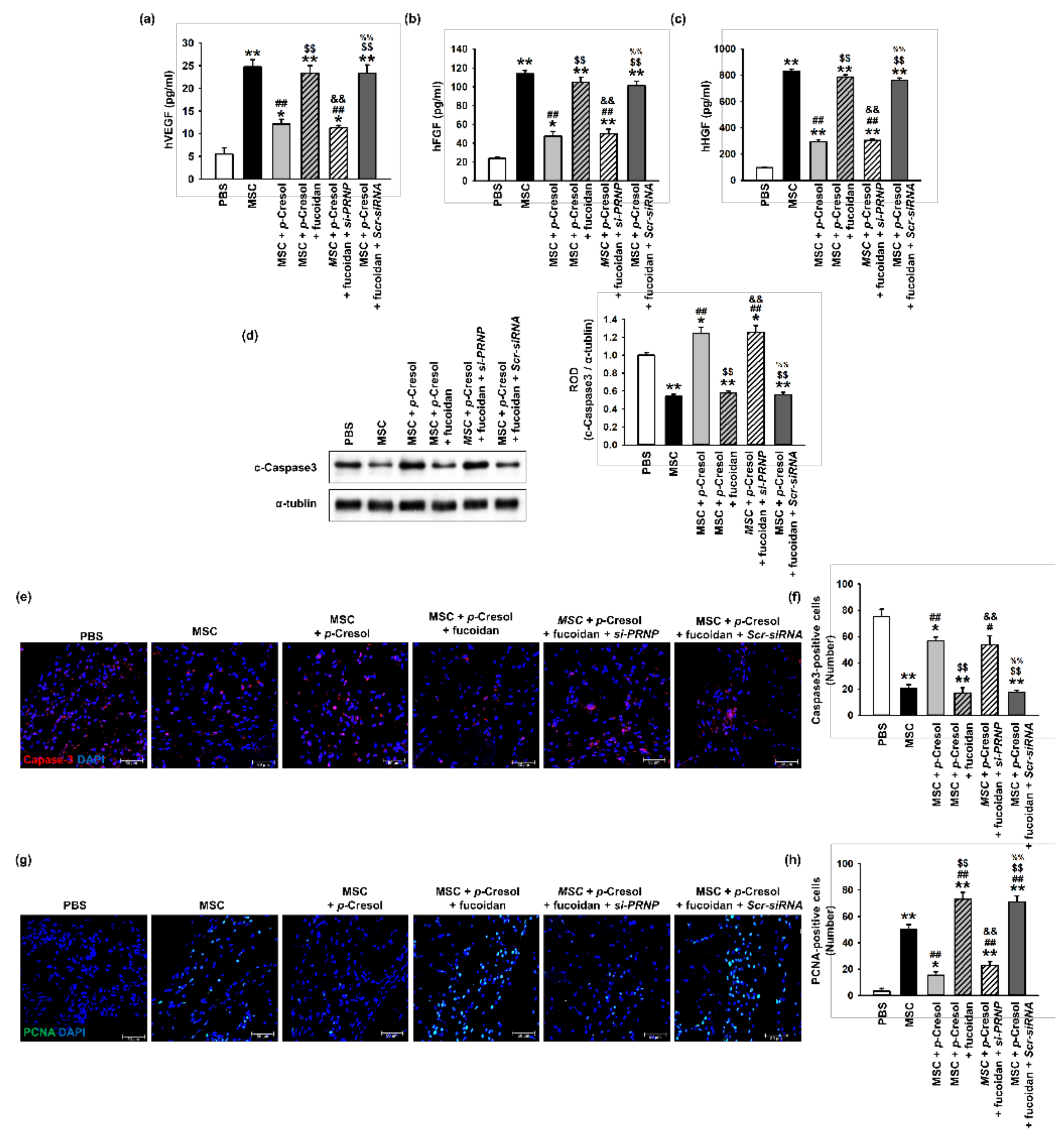
Figure 7.
Effect of fucoidan on angiogenic cytokine expression and protection from apoptosis in a murine hindlimb ischemia model. (a–c) The expression of (a) hVEGF, (b) hFGF, and (c) hHGF in ischemic limb tissue lysates was determined by ELISA. Values represent the mean ± S.E.M. * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol, && p < 0.01 vs. fucoidan pretreatment MSCs with p-cresol, %% p < 0.01 vs. fucoidan pretreatment si-PRNP transfected MSCs with p-cresol. (d) Western blot analysis of cleaved caspase-3 expression in ischemic sites of mice injected with PBS, MSCs, MSCs with p-cresol treatment, fucoidan-pretreated MSCs with p-cresol treatment, fucoidan-pretreated si-PRNP-transfected MSCs with p-cresol treatment, and fucoidan-pretreated si-Scramble transfection MSCs with p-cresol treatment at postoperative day 3. The level of cleaved caspase-3 was quantified by densitometry relative to α-tubulin levels (n = 3) Values represent mean ± SEM * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan pretreated si-PRNP-transfected MSCs with p-cresol treatment. (e) At postoperative day 3, the level of apoptosis was evaluated by immunofluorescence staining for cleaved caspase-3 in ischemic limb tissues. Scale bar = 50 μm. (f) The levels of apoptosis were quantified as the number of cleaved caspase-3 positive cells. Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. PBS, # p < 0.05 and ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan pretreated si-PRNP transfected MSCs with p-cresol treatment. (g) At postoperative day 3, cell proliferation was evaluated by immunofluorescent staining for PCNA in ischemic limb tissues. Scale bar = 50 μm. (h) The levels of cell proliferation were quantified as the number of PCNA positive cells. Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan-pretreated si-PRNP transfected MSCs with p-cresol treatment.
Figure 7.
Effect of fucoidan on angiogenic cytokine expression and protection from apoptosis in a murine hindlimb ischemia model. (a–c) The expression of (a) hVEGF, (b) hFGF, and (c) hHGF in ischemic limb tissue lysates was determined by ELISA. Values represent the mean ± S.E.M. * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol, && p < 0.01 vs. fucoidan pretreatment MSCs with p-cresol, %% p < 0.01 vs. fucoidan pretreatment si-PRNP transfected MSCs with p-cresol. (d) Western blot analysis of cleaved caspase-3 expression in ischemic sites of mice injected with PBS, MSCs, MSCs with p-cresol treatment, fucoidan-pretreated MSCs with p-cresol treatment, fucoidan-pretreated si-PRNP-transfected MSCs with p-cresol treatment, and fucoidan-pretreated si-Scramble transfection MSCs with p-cresol treatment at postoperative day 3. The level of cleaved caspase-3 was quantified by densitometry relative to α-tubulin levels (n = 3) Values represent mean ± SEM * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan pretreated si-PRNP-transfected MSCs with p-cresol treatment. (e) At postoperative day 3, the level of apoptosis was evaluated by immunofluorescence staining for cleaved caspase-3 in ischemic limb tissues. Scale bar = 50 μm. (f) The levels of apoptosis were quantified as the number of cleaved caspase-3 positive cells. Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. PBS, # p < 0.05 and ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan pretreated si-PRNP transfected MSCs with p-cresol treatment. (g) At postoperative day 3, cell proliferation was evaluated by immunofluorescent staining for PCNA in ischemic limb tissues. Scale bar = 50 μm. (h) The levels of cell proliferation were quantified as the number of PCNA positive cells. Values represent mean ± SEM. * p < 0.05 and ** p < 0.01 vs. PBS, ## p < 0.01 vs. MSC alone, $$ p < 0.01 vs. MSCs with p-cresol treatment, && p < 0.01 vs. fucoidan-pretreated MSCs with p-cresol treatment, %% p < 0.01 vs. fucoidan-pretreated si-PRNP transfected MSCs with p-cresol treatment.
![Marinedrugs 16 00121 g007]()
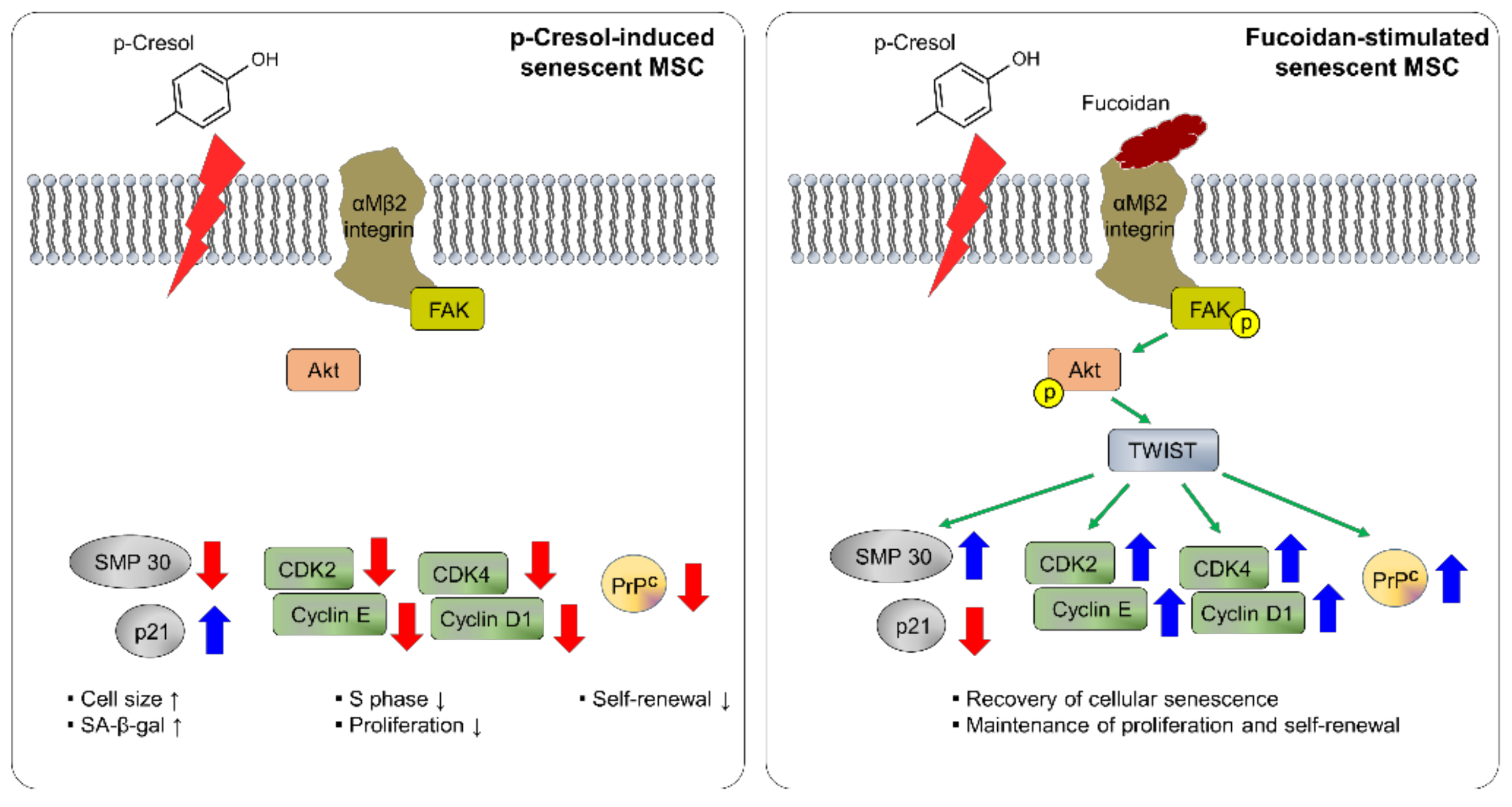
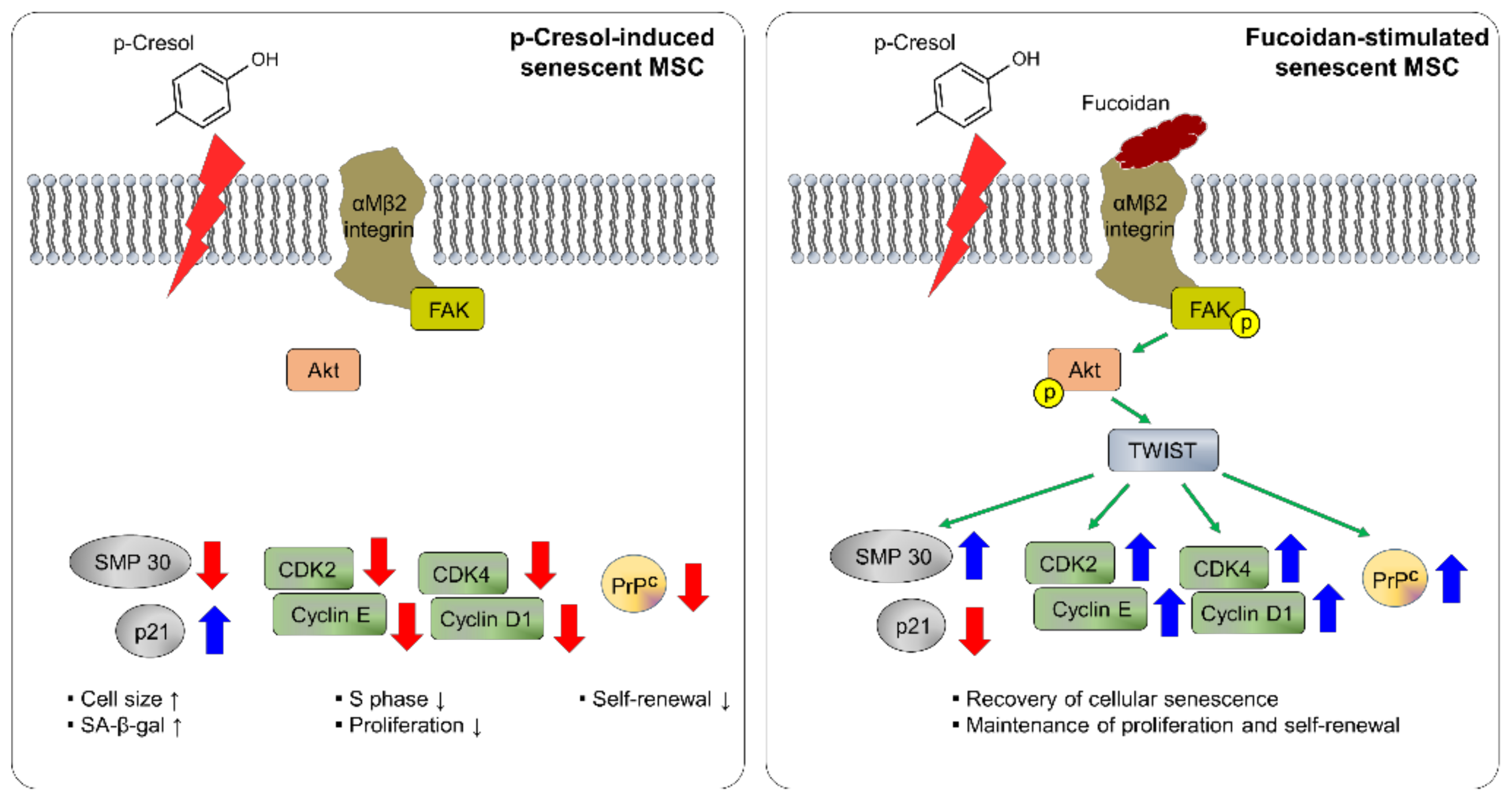
Figure 8.
Schematic representation of possible mechanisms by which fucoidan protects against p-cresol-induced senescence in MSCs through the FAK-Akt-TWIST signal pathway. Fucoidan rescues the p-cresol-mediated inhibition of senescence- and cell cycle-associated proteins via FAK-Akt-TWIST signal transduction. In addition, fucoidan-mediated maintenance of proliferation is dependent on the TWIST-PrPC axis. These effects of fucoidan protect against cellular senescence caused by p-cresol exposure in MSCs.
Figure 8.
Schematic representation of possible mechanisms by which fucoidan protects against p-cresol-induced senescence in MSCs through the FAK-Akt-TWIST signal pathway. Fucoidan rescues the p-cresol-mediated inhibition of senescence- and cell cycle-associated proteins via FAK-Akt-TWIST signal transduction. In addition, fucoidan-mediated maintenance of proliferation is dependent on the TWIST-PrPC axis. These effects of fucoidan protect against cellular senescence caused by p-cresol exposure in MSCs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}