AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity

Abstract
:1. Introduction
2. Results and Discussion
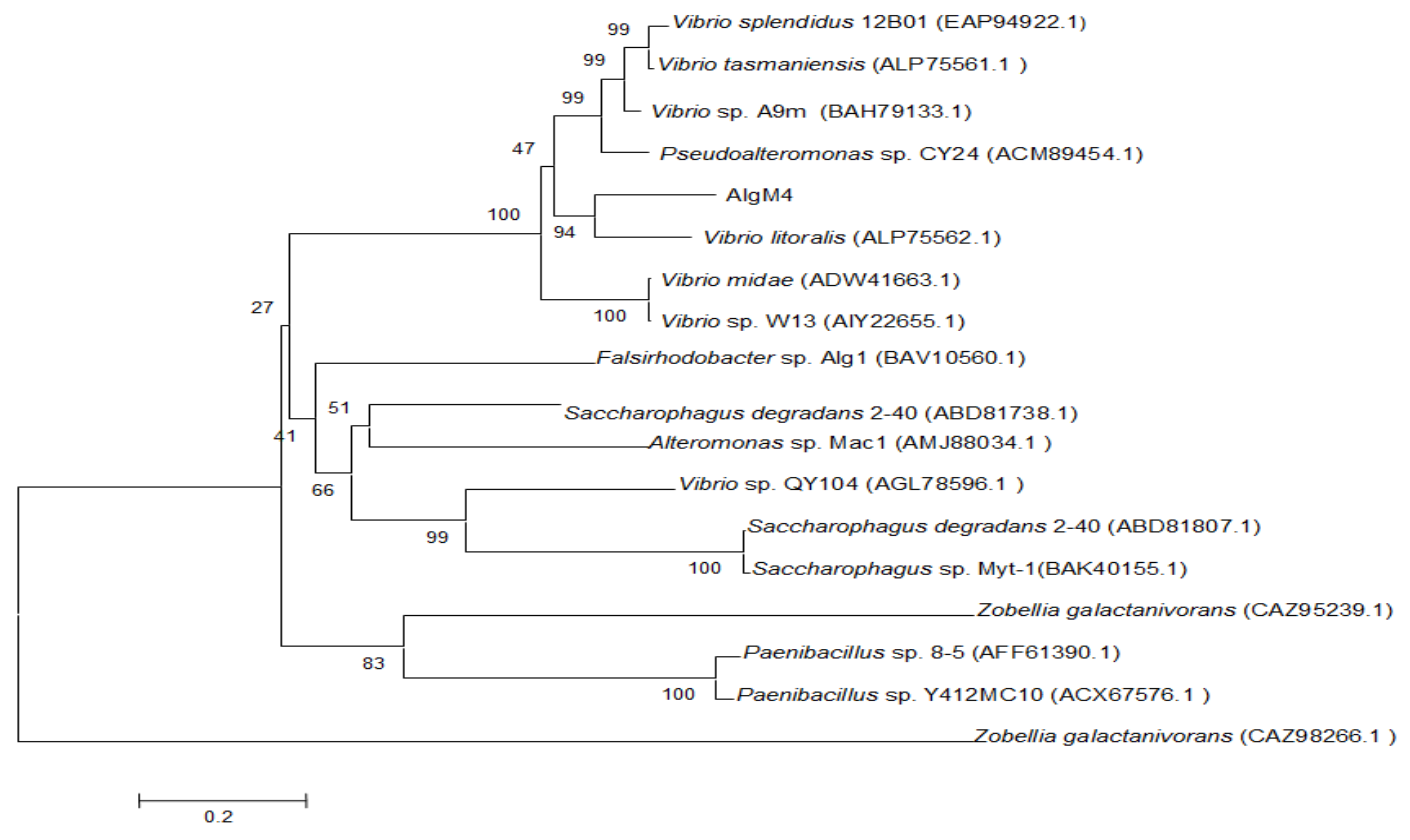
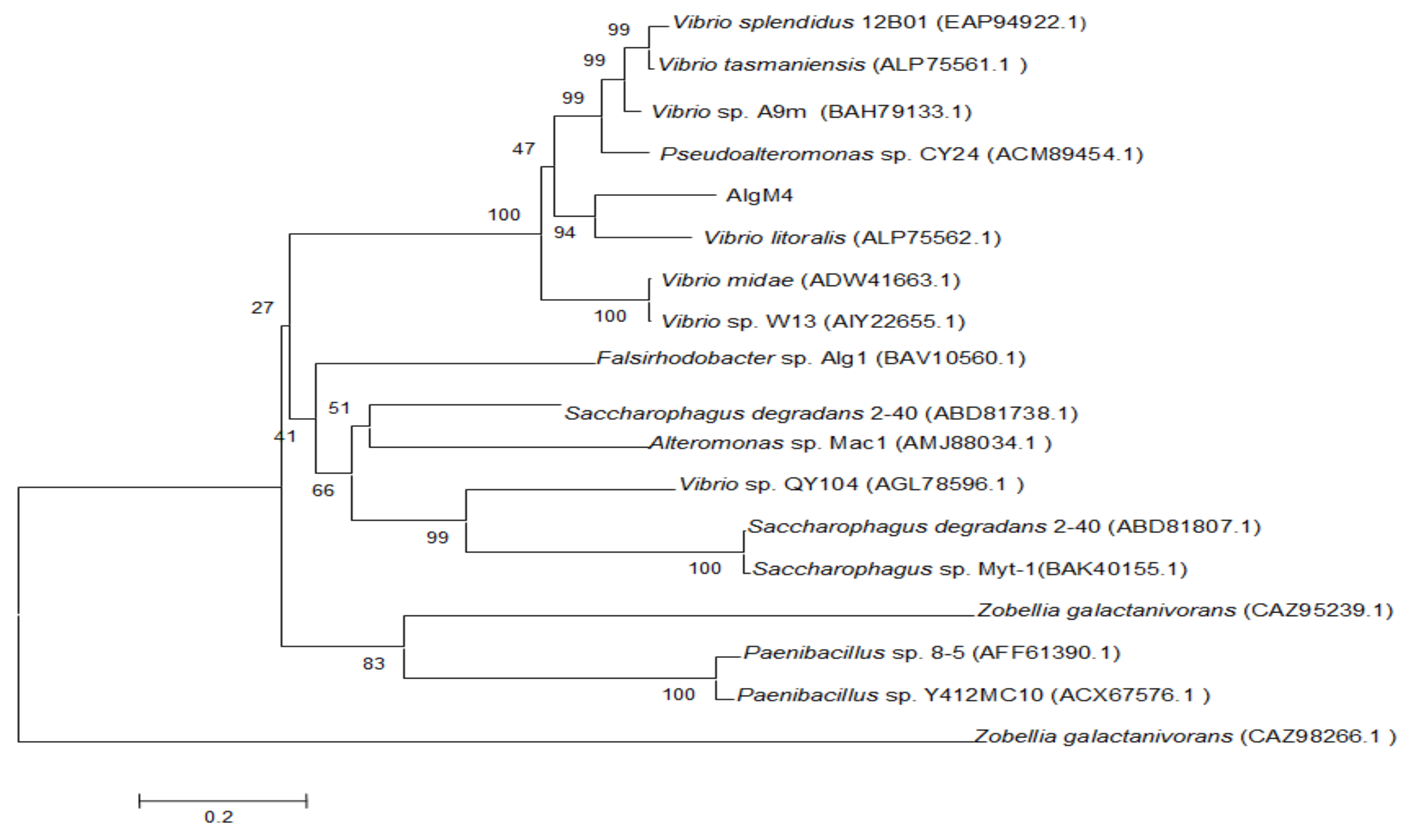
2.1. Analysis of the AlgM4 Sequence
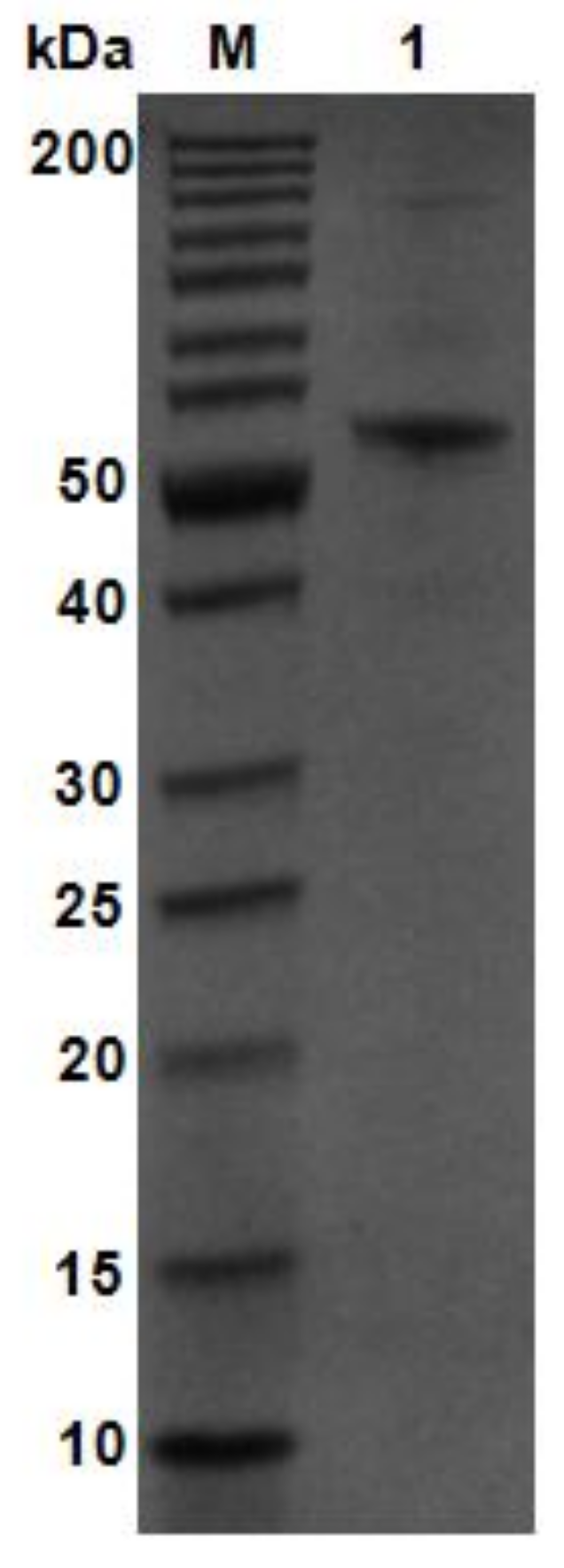
2.2. Purification and Enzymatic Activity of AlgM4
2.3. Vmax (Maximum Reaction Rate) and Km (Michaelis Constant) of AlgM4
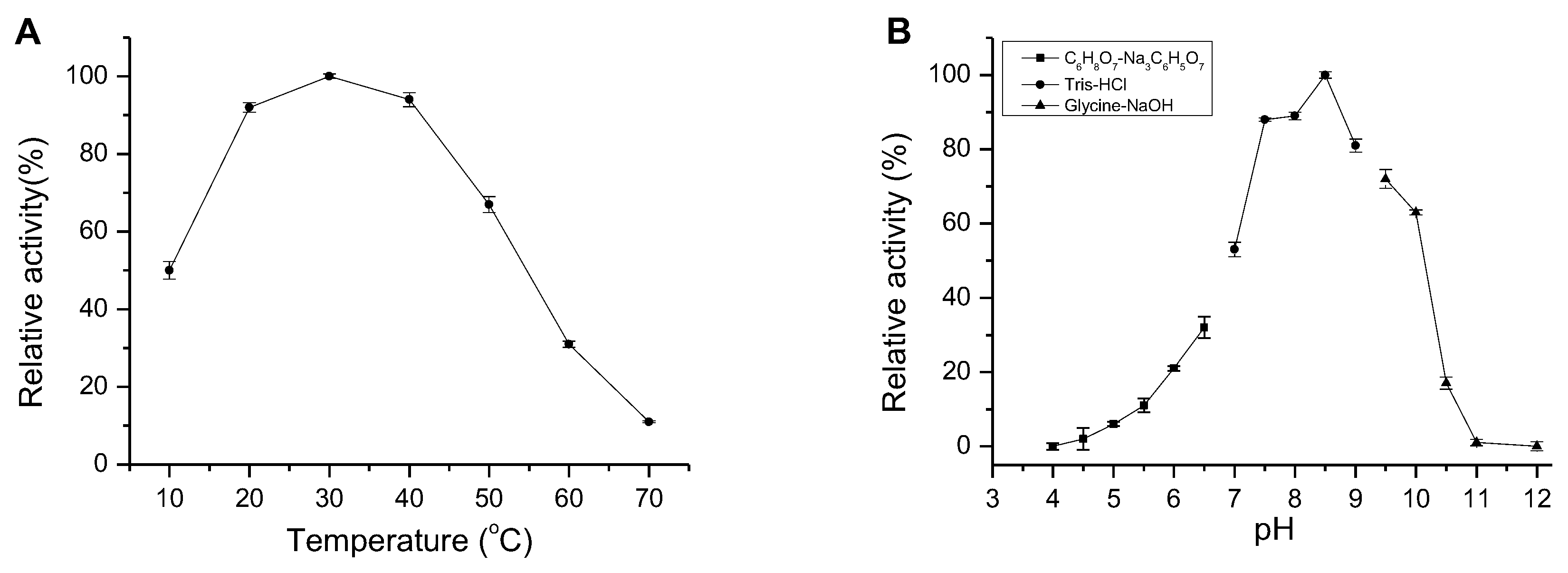
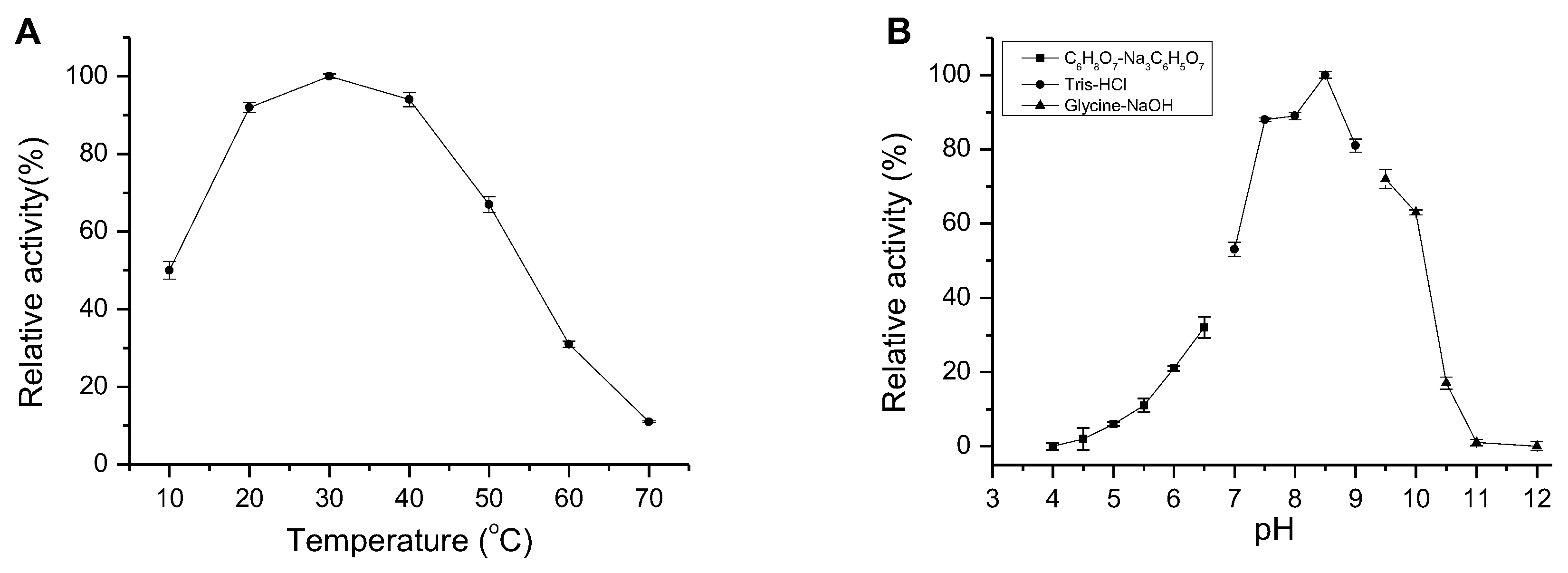
2.4. Enzymatic Characteristics of AlgM4
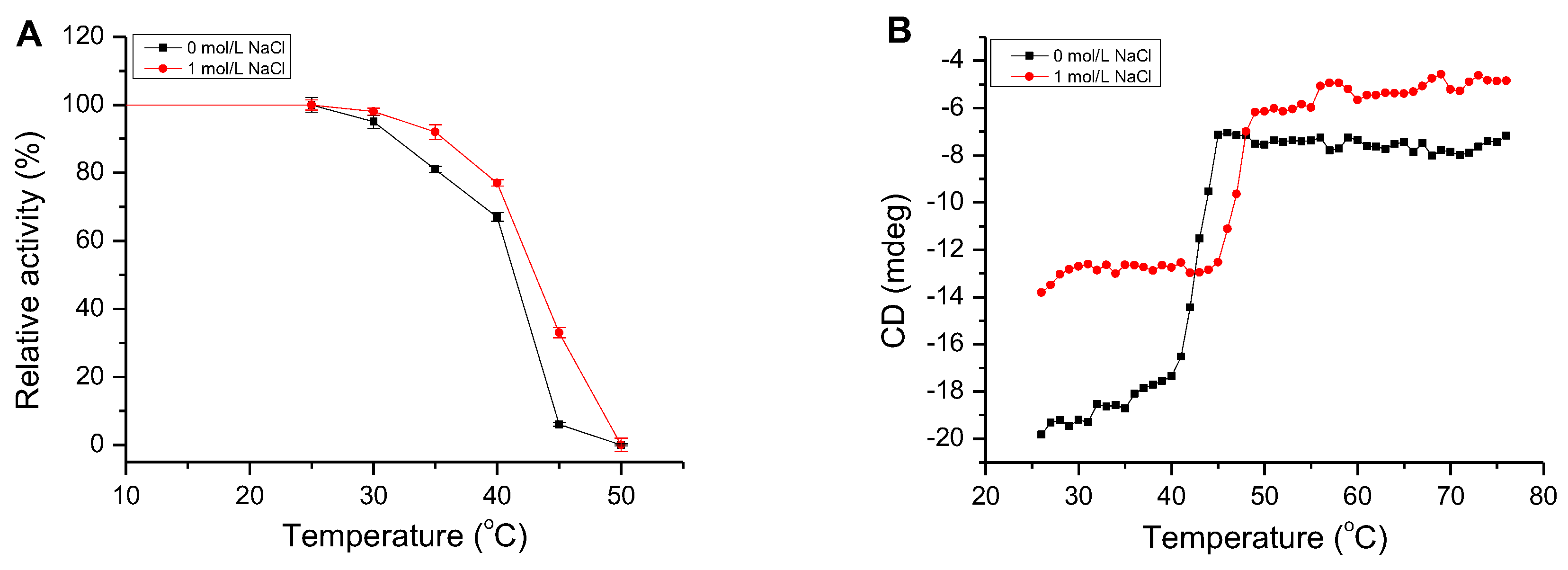
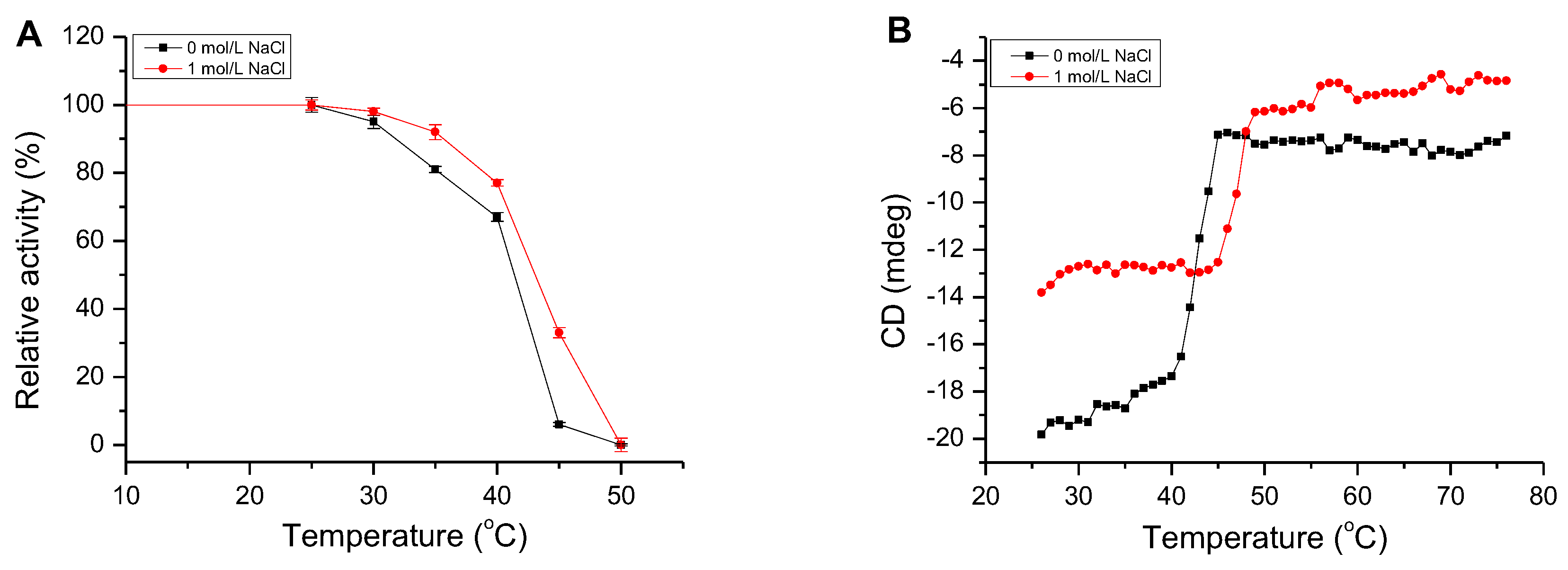
2.5. Effect of NaCl on AlgM4 Activity
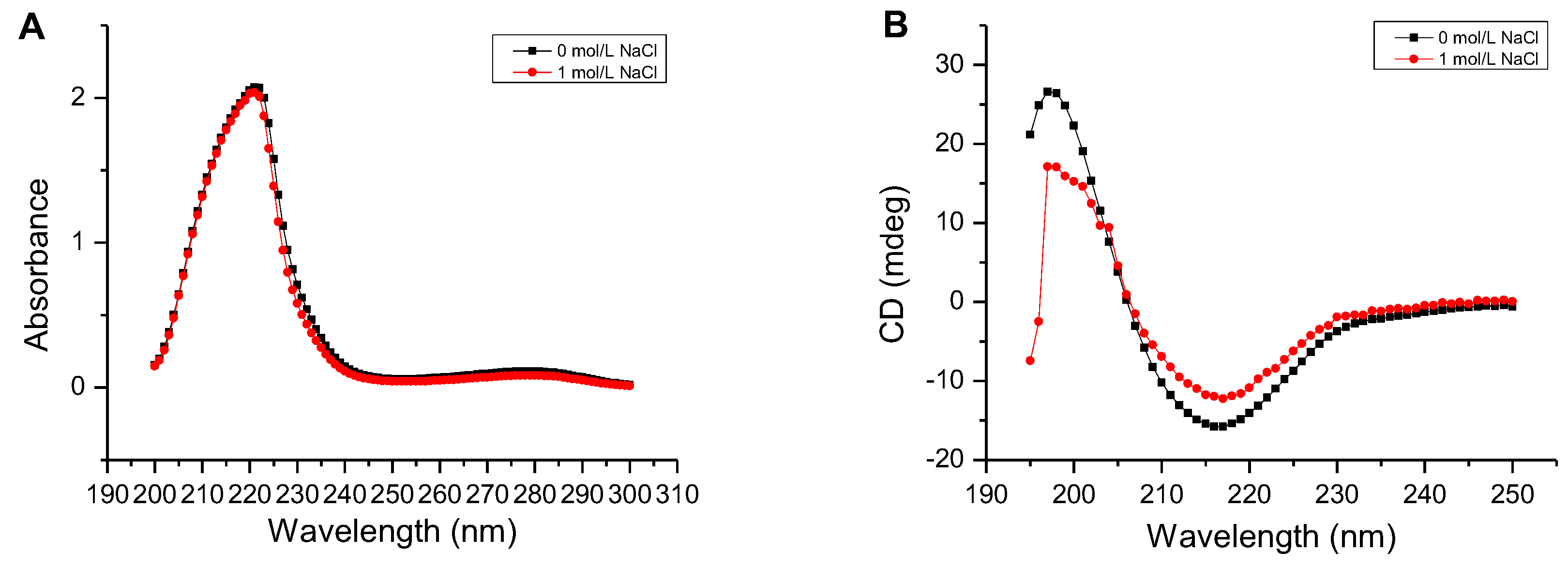
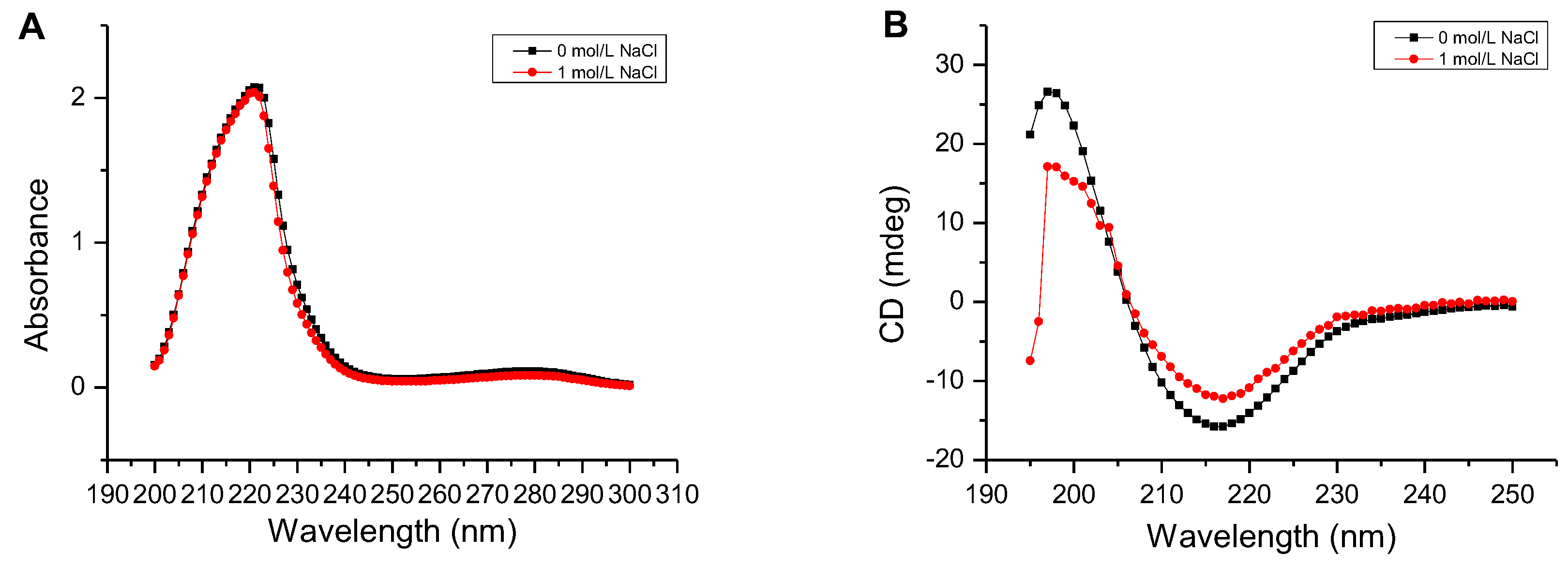
2.6. Determination of the Secondary Structure and Thermal Denaturation Temperature of AlgM4
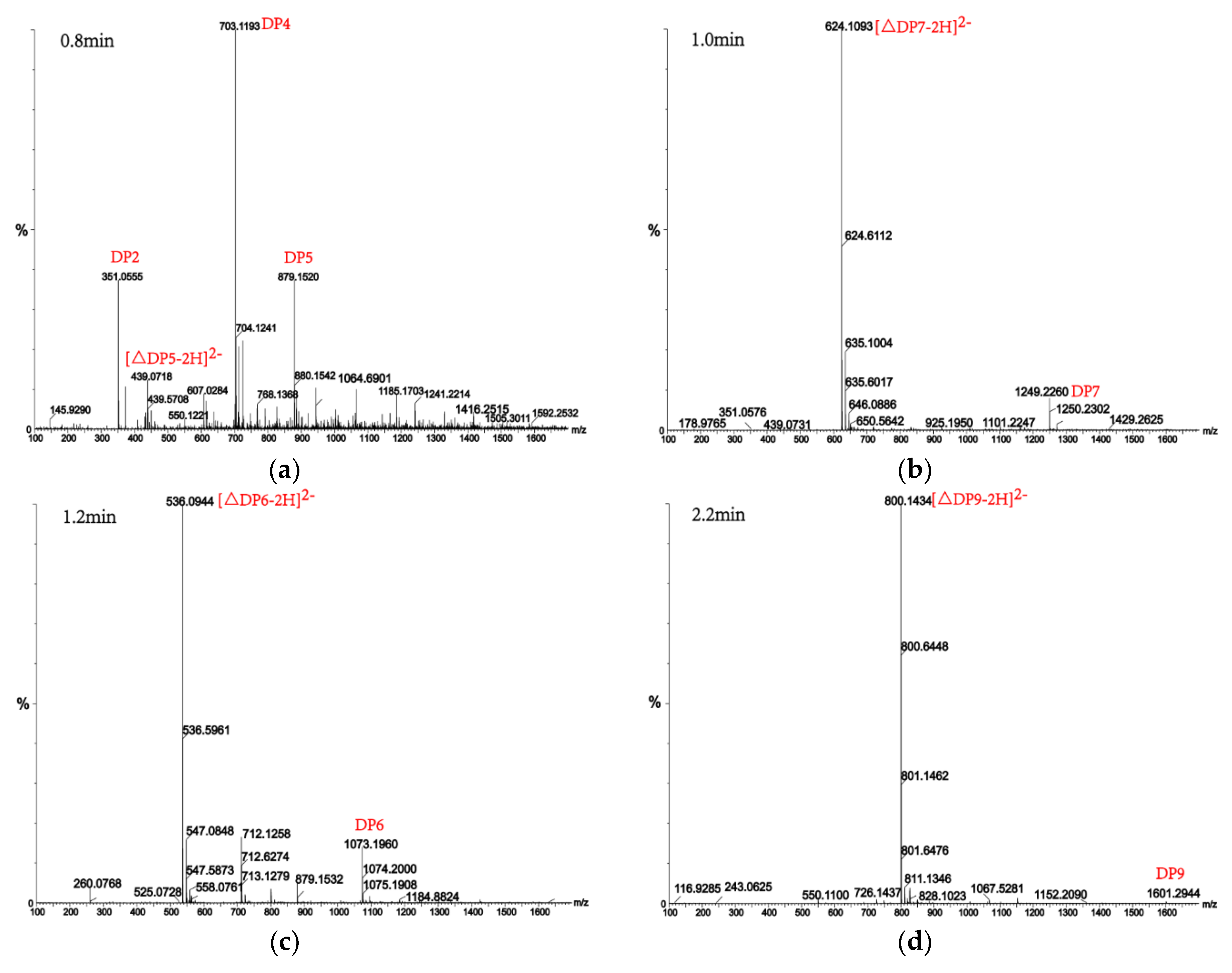
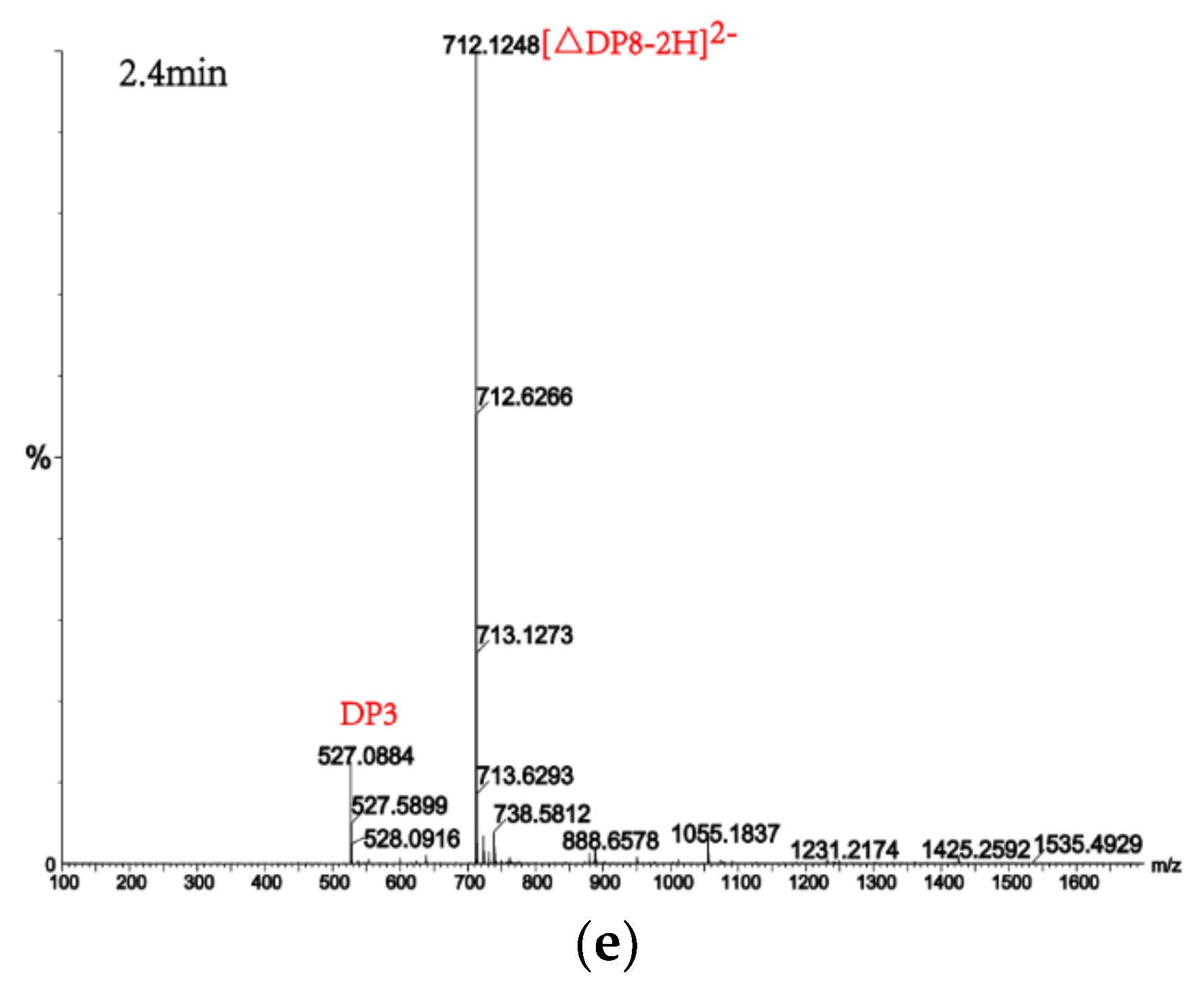
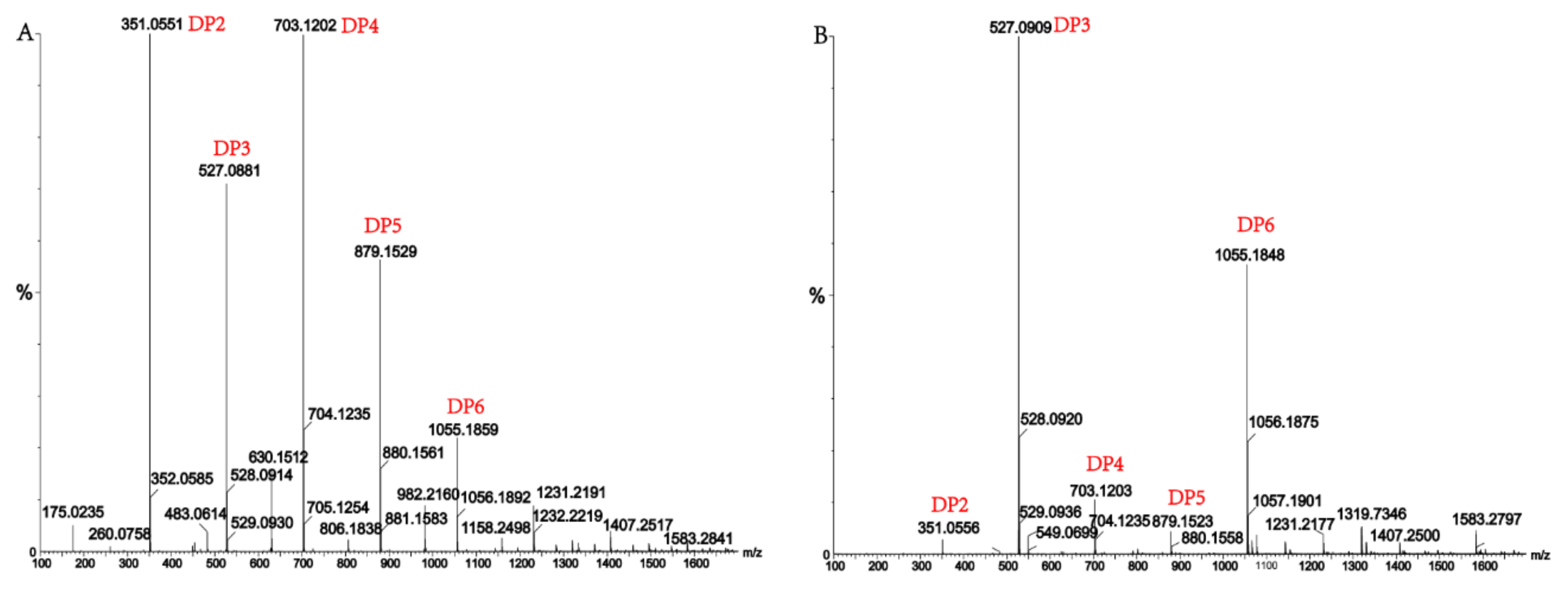
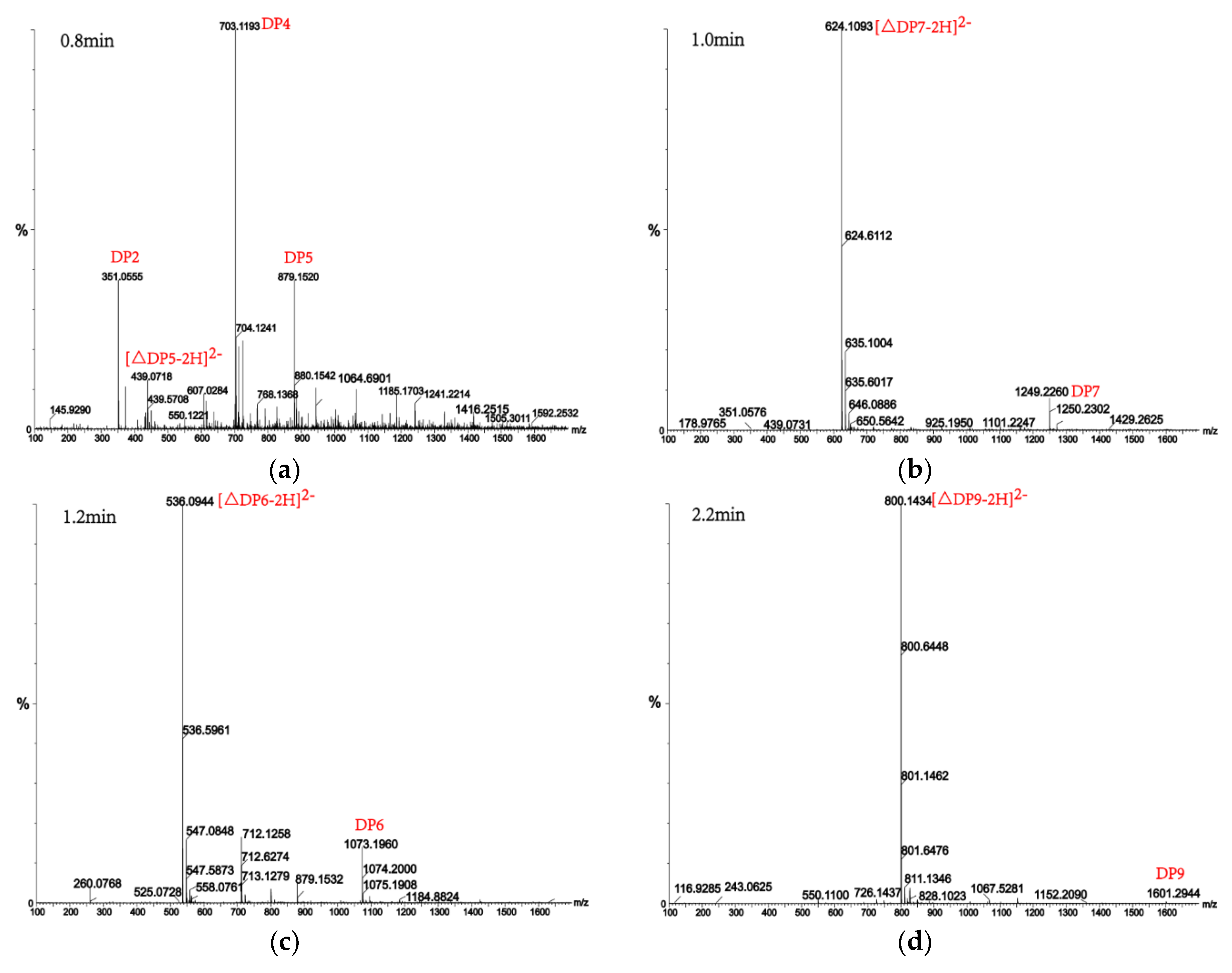
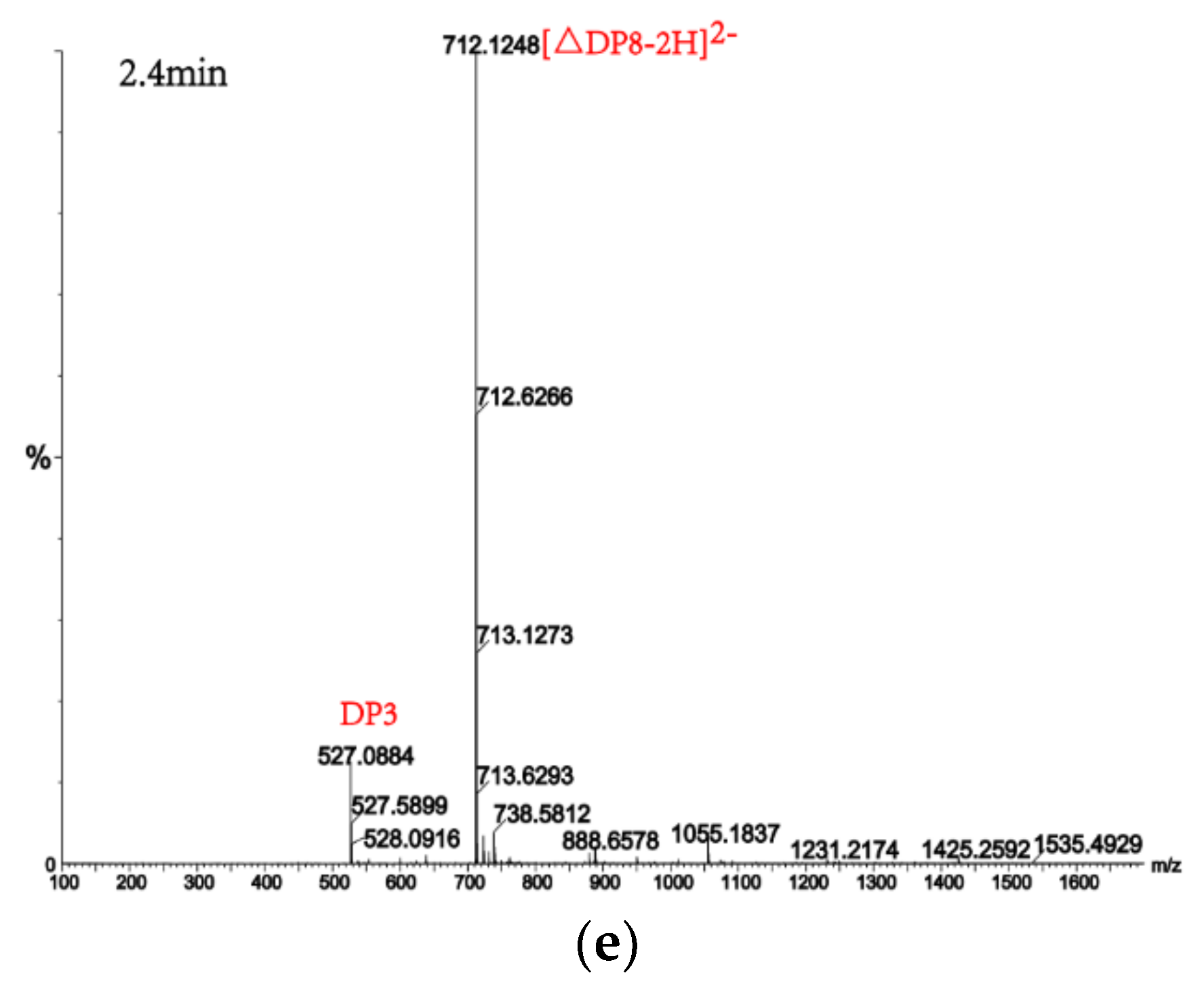
2.7. Analysis of the Products of AlgM4-Mediated Enzymolysis of Alginate by Ultra-Performance Liquid Chromatography (UPLC)–Quadrupole Time-of-Flight (QTOF)–Mass Spectrometry (MS)/MS
3. Materials and Methods
3.1. The Bacterium
3.2. Cloning and Expression of the algM4 Gene
3.3. Purification of AlgM4
3.4. Determination of the Enzymatic Properties of AlgM4
3.5. Determination of the Kinetic Constants for AlgM4 Activity
3.6. The Effects of NaCl on the Secondary Structure and Thermal Stability of AlgM4
3.7. Analysis of the Products of AlgM4-Mediated Enzymolysis Using UPLC–QTOF–MS/MS
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hernandez-carmona, G.; Mchugh, D.J.; Iopez-gutierrez, F. Pilot plant scale extraction of alginates from Macrocystis pyrifera. Appl. Phycol. 2000, 11, 493–502. [Google Scholar] [CrossRef]
- Liu, H.; Yin, H.; Zhang, Y.H.; Zhao, X.M. Research progress on biological activities of alginate oligosaccharides. Nat. Prod. Res. 2012, S1, 201–204. [Google Scholar]
- Gacesa, P. Alginate. Carbohydr. Polym. 1988, 8, 161–182. [Google Scholar] [CrossRef]
- Tako, M. Chemical characterization of acetyl fucoidan and alginate from commercially cultured cladosiphon okamuranus. Bot. Mar. 2000, 43, 393–398. [Google Scholar] [CrossRef]
- Tako, M.; Kiyuna, S.; Uechi, S.; Hongo, F. Isolation and characterization of alginic acid from commercially cultured Nemacystus decipiens (Itomozuku). Biosci. Biotechnol. Biochem. 2001, 65, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Tang, L.W.; Huang, S.S.; Chagan, I. Research progress of bioethanol from alginate fermentation. China Biotechnol. 2013, 33, 122–127. [Google Scholar]
- Schaumann, K.; Weide, G. Enzymatic degradation of alginate by marine fungi. Hydrobiologia 1990, 204, 589–596. [Google Scholar] [CrossRef]
- Lee, S.I.; Choi, S.H.; Lee, E.Y.; Kim, H.S. Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl. Microbiol. Biotechnol. 2012, 95, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Inoue, A.; Tanaka, H.; Ojima, T. cDNA cloning of an alginate lyase from a marine gastropod Aplysia kurodai and assessment of catalytically important residues of this enzyme. Biochimie 2011, 93, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Wang, L.; Inoue, A.; Ojima, T. cDNA cloning and bacterial expression of a PL-14 alginate lyase from a herbivorous marine snail Littorina brevicula. Carbohydr. Res. 2012, 360, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Jie, Y.; Zhang, X.Y.; Mei, S.; Song, X.Y.; Chen, X.L.; Zhang, Y.Z. Cultivable Alginate Lyase-Excreting Bacteria Associated with the Arctic Brown Alga Laminaria. Mar. Drugs 2012, 10, 2481–2491. [Google Scholar]
- Xiao, L.; Han, F.; Yang, Z.; Lu, X.Z.; Yu, W.G. A Novel Alginate Lyase with High Activity on Acetylated Alginate of Pseudomonas aeruginosa, FRD1 from Pseudomonas sp. QD03. World J. Microbio. Biotechnol. 2006, 22, 81–88. [Google Scholar] [CrossRef]
- Li, S.; Yang, X.; Zhang, L.; Yu, W.; Han, F. Cloning, Expression, and Characterization of a Cold-Adapted and Surfactant-Stable Alginate Lyase from Marine Bacterium Agarivorans sp. L11. J. Microbiol. Biotechnol. 2015, 25, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor sp. SB155-2 Thriving at Deep-sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Uchimura, K.; Miyazaki, M.; Nogi, Y.; Horikoshi, K. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles 2009, 13, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Sheng, D.; Fei, X.; Fang, D.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a New Cold-Adapted and Salt-Activated Polysaccharide Lyase Family 7 Alginate Lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, e30105. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Chung, J.H.; Wang, D.; Lee, J.; Woo, H.C.; Choi, I.G.; Kim, K.H. Depolymerization of alginate into a monomeric sugar acid using Alg17C, an exo-oligoalginate lyase cloned from Saccharophagus degradans 2-40. Appl. Microbiol. Biotechnol. 2012, 93, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, Y.; Iwasaki, K.; Muramatsu, T. Action of poly (alpha-l-guluronate)lyase from Corynebacterium sp. ALY-1 strain on saturated oligoguluronates. J. Agric. Chem. Soc. Jpn. 1998, 62, 1055–1060. [Google Scholar]
- Thiangyian, W.; Preston, L.A.; Schiller, N.L. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar]
- Sawabe, T.; Takahashi, H.; Ezura, Y.; Gacesa, P. Cloning, sequence analysis and expression of Pseudoalteromonas elyakovii IAM 14594 gene (alyPEEC) encoding the extracellular alginate lyase. Carbohydr. Res. 2001, 335, 11–21. [Google Scholar] [CrossRef]
- Miyake, O.; Hashimoto, W.; Murata, K. An exotype alginate lyase in Sphingomonas sp. A1: Overexpression in Escherichia coli, purification, and characterization of alginate lyase IV (A1-IV). Protein Expr. Purif. 2003, 29, 33–41. [Google Scholar] [CrossRef]
- Ochiai, A.; Yamasaki, M.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of exotype alginate lyase Atu3025 from Agrobacterium tumefaciens. J. Biol. Chem. 2010, 285, 24519–24528. [Google Scholar] [CrossRef] [PubMed]
- Garron, M.L.; Cygler, M. Structural and mechanistic classification of uronic acid-containing polysaccharide lyases. Glycobiology 2010, 20, 1547–1573. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, L.; Chen, Y.; Ni, H.; Xiao, A.; Cai, H. Characterization of an extracellular biofunctional alginate lyase from marine microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates. Microbiol. Res. 2016, 182, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Chen, M.; Yin, H.; Du, Y.; Ning, L. Enzymatic hydrolysis of alginate to produce oligosaccharides by a new purified endo-type alginate lyase. Mar. Drugs 2016, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, S.S.; Hehemann, J.H.; Polz, M.F.; Lee, J.K.; Zhao, H. Comparative biochemical characterization of three exolytic oligoalginate lyases from Vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl. Environ. Microbio. 2014, 80, 4207–4214. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and characterization of a novel oligoalginate lyase from a newly isolated bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Bao, M.; Wu, Y.; Yu, W.; Han, F. Family 13 carbohydrate-binding module of alginate lyase from Agarivorans sp. L11 enhances its catalytic efficiency and thermostability, and alters its substrate preference and product distribution. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Kurakake, M.; Kitagawa, Y.; Okazaki, A.; Shimizu, K. Enzymatic Properties of Alginate Lyase from Paenibacillus sp. S29. Appl. Biochem. Biotechnol. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tusi, S.K.; Khalaj, L.; Ashabi, G.; Kiaei, M.; Khodagholi, F. Alginate oligosaccharide protects against endoplasmic reticulum- and mitochondrial-mediated apoptotic cell death and oxidative stress. Biomaterials 2011, 32, 5438–5458. [Google Scholar] [CrossRef] [PubMed]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.I.; Matsubara, Y. Purification of Alginate Oligosaccharides with Root Growth-promoting Activity toward Lettuce. Biosci. Biotechnol. Biochem. 2000, 64, 1067–1070. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Recover (%) | Purification (Fold) |
|---|---|---|---|---|---|
| Crude enzyme AlgM4 | 58.352 | 73878 | 1266 | 100 | 1 |
| Purified AlgM4 | 11.55 | 53570 | 4638 | 72.5 | 3.7 |
| Reagent | Concentration (mmol/L) | Relative Activity (%) |
|---|---|---|
| None | - | 100 ± 0.3 |
| CaCl2 | 1 | 97 ± 0.1 |
| MgCl2 | 1 | 114 ± 1.1 |
| KCl | 1 | 91 ± 0.9 |
| CuCl2 | 1 | 75 ± 1.7 |
| MnCl2 | 1 | 77 ± 2.4 |
| ZnCl2 | 1 | 18 ± 4.5 |
| EDTA | 1 | 65 ± 6.2 |
| SDS | 1 | 3 ± 3.1 |
| NaCl Concentration (mol/L) | Relative Activity (%) |
|---|---|
| 0 | 100 ± 0.2 |
| 0.1 | 229 ± 5.6 |
| 0.2 | 286 ± 6.4 |
| 0.3 | 356 ± 3.8 |
| 0.4 | 447 ± 1.4 |
| 0.5 | 481 ± 0.8 |
| 0.6 | 528 ± 3.1 |
| 0.7 | 572 ± 0.4 |
| 0.8 | 628 ± 3.4 |
| 0.9 | 664 ± 1.9 |
| 1.0 | 741 ± 3.2 |
| 1.2 | 462 ± 5.8 |
| 1.4 | 236 ± 4.5 |
| Enzyme | α-Helix (%) | β-Sheet (%) | β-Turu (%) | Random Coil (%) |
|---|---|---|---|---|
| AlgM4 | 12.4 | 38.2 | 21.3 | 28 |
| AlgM4 + 1 mol/L NaCl | 10.8 | 36.5 | 22.9 | 29.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Wang, Q.; Lu, M.; Xu, C.; Li, F.; Zhang, R.; Liao, W.; Huang, S. AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity. Mar. Drugs 2018, 16, 120. https://doi.org/10.3390/md16040120
Huang G, Wang Q, Lu M, Xu C, Li F, Zhang R, Liao W, Huang S. AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity. Marine Drugs. 2018; 16(4):120. https://doi.org/10.3390/md16040120
Chicago/Turabian StyleHuang, Guiyuan, Qiaozhen Wang, Mingqian Lu, Chao Xu, Fei Li, Rongcan Zhang, Wei Liao, and Shushi Huang. 2018. "AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity" Marine Drugs 16, no. 4: 120. https://doi.org/10.3390/md16040120
APA StyleHuang, G., Wang, Q., Lu, M., Xu, C., Li, F., Zhang, R., Liao, W., & Huang, S. (2018). AlgM4: A New Salt-Activated Alginate Lyase of the PL7 Family with Endolytic Activity. Marine Drugs, 16(4), 120. https://doi.org/10.3390/md16040120