Abstract
Three new polyketides, lactomycins A (1)–C (3), were isolated from the culture broth of a marine-derived Streptomyces sp. ACT232 as cathepsin B inhibitors. Their structures were determined by a combination of NMR and MS data analyses to be the dephosphorylated derivatives of a phoslactomycin class of metabolites. Lactomycins exhibited cathepsin B inhibitory activity (IC50 0.8 to 4.5 μg/mL). Even though the biosynthetic gene clusters found in the genome of the current strain have high similarity to those of phoslactomycin, neither phoslactomycins nor leustroducsins were detected by LC-MS analyses of the crude extract.
1. Introduction
The majority of the structurally and functionally diverse anticancer agents have been derived from terrestrial naturel products [1]. Although marine organisms were reported to exhibit a high incidence of cytotoxic activity [2], it is only recently that a few anticancer agents derived from marine organisms have been approved [3]. We have been searching for anticancer compounds from marine organisms [4], and payed attention to cathepsin B as a target to discover anticancer agents with a mode of action distinct from cell cycle arrest. Cathepsin B is a lysosomal protease of the papain family, which has been considered as a superb target for cancer chemotherapy [5]. It is activated and secreted in the tumor microenvironment and enhances tumor metastasis and infiltration by cleaving extracellular matrix proteins and activating several prominent proteases. It is overexpressed in various cancers and its knockout retards cell proliferation and tumor growth. During our screening program it was found that the crude extract of the Streptomyces sp. ACT232, a marine-derived actinomycete, showed cathepsin B inhibitory activity [6]. We isolated and studied the structures and biosynthesis of the active constituents.
2. Results
2.1. Isolation
The mycelia from the culture of Streptomyces sp. ACT232 were collected by vacuum filtration. Resins (Amberlite XAD 16N) were added to the filtrate to allow adsorption of the metabolites. The acetone extracts of the mycelia and resins were combined and partitioned between n-BuOH and H2O. The n-BuOH fraction was separated by ODS flash chromatography to give five fractions. The active fraction was separated by reversed-phase HPLC to afford three metabolites termed lactomycins A–C (Figure 1, 1–3).
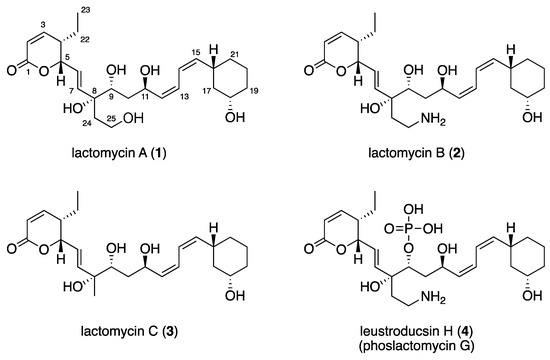
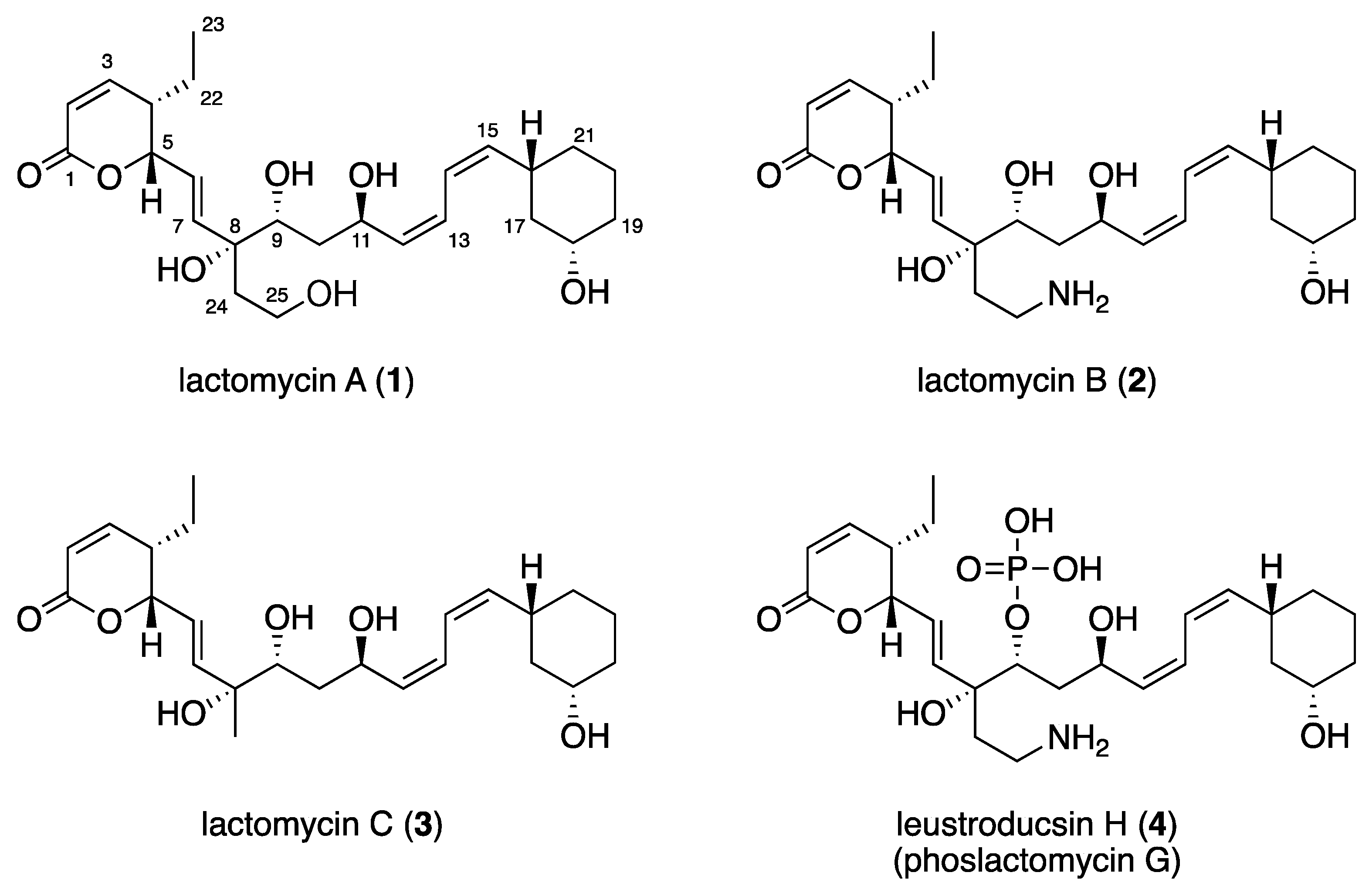
Figure 1.
Structures of lactomycins A (1)–C (3) and phoslactomycin G (4).
2.2. Structure Elucidation
The molecular formula of lactomycin A (1), C25H38O7, was established by HRESIMS. The analyses of 1H NMR and HSQC spectra of 1 indicated the presence of eight protonated sp2 carbons (δH/δC 7.03/151.5, 6.10/122.5, 6.09/121.8, 5.96/120.5, 5.84/137.4, 5.69/123.7, 5.40/137.6, 5.25/137.8), four oxymethines (δH/δC 5.02/80.7, 4.58/63.5, 3.53/72.6, 3.37/68.5), one oxygenated methylene (δH/δC 3.53, 3.46/57.9), six methylenes, and one terminal methyl (Table 1). The 13C NMR spectrum revealed the presence of two additional non-protonated carbons at δC 164.0 and 76.6 (Table 1). Three partial structures of C2–C7 plus C22–C23, C9–C21, and C24–C25 were established by interpretation of the 1H-1H COSY spectrum. HMBC correlations from H2, H3, and H5 to the carbon at δC 164.0 (C1) indicated that C2 to C5 comprised an α,β-unsaturated δ-lactone moiety. There was a 1,3-disubstituted cyclohexane moiety (C16 to C21) in the second partial structure, in which C18 was substituted by an oxygen atom judging from the 1H and 13C NMR chemical shifts (δH/δC 3.37/68.5). The planar structure of the C9 to C15 portion was elucidated by analysis of the COSY spectrum. The third partial structure was composed of disubstituted ethane in which a substituent at C25 was assigned as an oxygen atom on the basis of the 1H and 13C NMR data (δH/δC 3.46, 3.53/57.9). The three partial structures were all attached to the tetrasubstituted carbon bearing an oxygen atom (C8), as demonstrated by HMBC correlations from H7, H9, and H2-24 to C8. Considering the molecular formula, the oxygen atoms at C8, C9, C11, C18, and C25 were all assigned as free hydroxyl groups.
Table 1.
1H NMR (600 MHz) and 13C NMR (150 MHz) Data for 1 in DMSO-d6 and 2 and 3 in CD3OD.
The coupling constant of 9.7 Hz between H2 and H3 suggested that they are cis to each other, which was supported by a prominent NOESY cross peak between these protons. The E-geometry of the C6 to C7 double bond was assigned from the coupling constant of 15.6 Hz between H6 and H7. Methylene protons on C10 were diastereotopic and coupling between H10a (δH 0.98)/H9 (10 Hz), H10a/H11(1.4 Hz), and H10b (δH 1.58)/H11 (10 Hz) was observed, whereas coupling between H10b/H9 was not observed. Analysis of these coupling constant values suggested the 9R*, 11R* relative configuration. The coupling constants between H12 and H13 and between H14 and H15, as well as between H13 and H14, were determined by analyzing the AB-multiplet for H13 and H14, which suggested that the three coupling constants were all 11 Hz, indicating that the C12 to C13 and C14 to C15 double bonds were both Z. This assignment was supported by intense NOESY cross peaks H12/H13, H14/H15, H11/H14, and H13/H16. The C16 and C18 substituents of the cyclohexane ring were both equatorial, as shown by the large coupling constants observed for H16 and H18 to the adjacent axial methylene protons. The structural assignment conducted so far suggested 1 to be a dephospho-25-deamino-25-hydroxy-leustroducsin H (phoslactomycins G) (4) derivative [7,8]. Although the relative stereochemistry within the three partial structures was identical with that of leustroducsin H [9], due to the insulation by double bonds and the possible free rotation of the bond between C7 and C8, it was not possible to correlate the relative configurations between the partial structures nor assign that of C8 on the basis of the NOESY data. The NMR data of 1 was very close to those reported for phoslactomycins [10,11,12,13] and leustroducsins [14,15], except for those around C9 and C18, which were phosphorylated and esterified, respectively, in the latter compounds. The presence of an almost identical biosynthetic gene cluster with those previously reported for phoslactomycins (vide infra) suggests that the stereochemistry is shared between lactomycin A and phoslactomycins.
Lactomycin B (2) had a molecular formula of C25H39NO6 as determined by HRESIMS. Extensive 1D and 2D NMR analyses suggested that 2 had a carbon backbone identical to that of 1. Chemical shift values of C25 methylene (δH/δC 3.03, 2.97/37.4) in 2 were significantly different from those in 1 (δH/δC 3.53, 3.46/57.9) and suggested that C25 was substituted by an amino group. Therefore, compound 2 was a dephosphorylated derivative of leustroducsin H.
The molecular formula of lactomycin C (3) was determined to be C24H36O6, which is smaller than 1 by a CH2O unit. 1H and 13C data of 3 were almost identical to those of 2, except for the replacement of the aminoethyl group by a methyl group (δH/δC 1.27/24.8). This substitution was confirmed by interpretation of the 1D and 2D NMR data.
2.3. Biosynthetic Gene Cluster
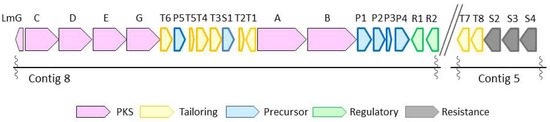
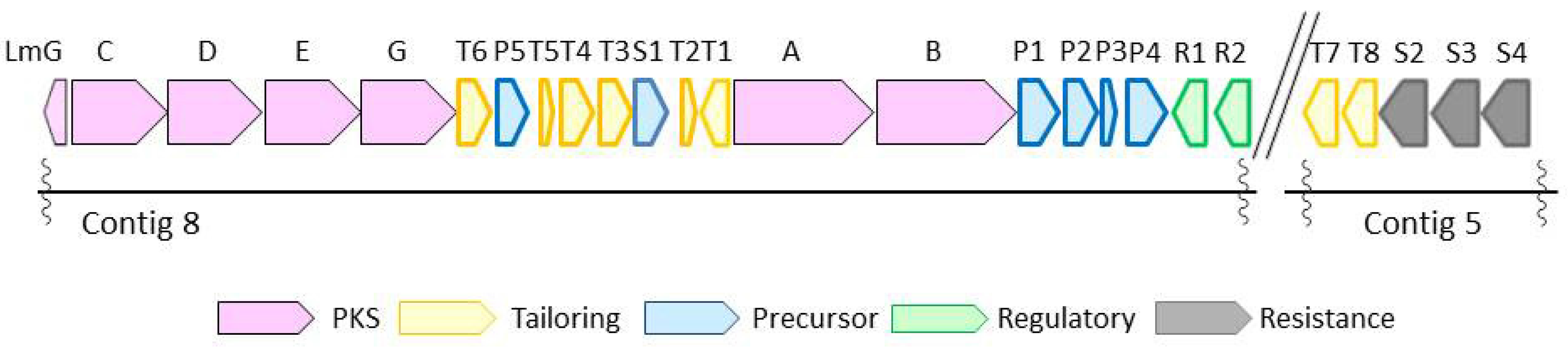
With structural difference only being exhibited in the state of phosphorylation and acylation, lactomycins can be considered as biosynthetic intermediates of phoslactomycins. Because dephospho-25-deamino-25-hydroxy phoslactomycin B (NPLM), which is closely related to lactomycin A, has been isolated from the streptomycete that produces leustroducsins [16], we suspected that our streptomycete has the same sets of biosynthetic genes to produce phoslactomycins. We set out to reveal the biosynthetic gene cluster of lactomycins by the genome analysis of Streptomyces sp. ACT232. The genome was sequenced using Illumina Hiseq, followed by assembly with CLC Genomics Workbench to give 15 contigs which cover 7.5 Mbp in total. The sequence data in this study have been deposited in GenBank under Genbank ID LC364194 and LC365285. Analyses of contigs by antiSMASH 3.0 [17] implied the presence of biosynthetic gene clusters of 32 secondary metabolites, including a gene cluster parallel to those reported for the biosynthetic gene cluster of phoslactomycin. Biosynthetic gene clusters of phoslactomycins were previously identified in two species of streptomycetes, namely Streptomyces sp. HK-803 [18,19,20] and Streptomyces platensis SAM-0654 [21]. Whilst they contain almost the same sets of genes, their architectures are different; the gene cluster of phoslactomycin in Streptomyces sp. HK-803 is present in one cluster, whereas the gene cluster in Streptomyces platensis SAM-0654 is dispersed into two clusters. The lactomycin cluster is composed of six genes for the biosynthesis of cyclohexanecarboxyl-CoA (CHC) as the starter unit, six PKS genes, eight genes for post PKS modifications, four transporters, and two regulatory genes (Figure 2). The biosynthetic gene cluster of our strain was separately located in two contigs, as found in Streptomyces sp. SAM-0654, and all genes required for the biosynthesis of phoslactomycins exist in the genome. The deduced function of each lactomycin biosynthetic gene and identity with the previously reported corresponding genes are listed in Table 2. Our genes shared identity ranging from 83% to 98% with those of the HK803 strain and identity ranging from 83% to 97% with those of the SAM-0654 strain. The analysis suggested that the strain ACT232 should also produce a phoslactomycin class of metabolites. Nevertheless, we were not able to detect these metabolites in our culture by LC-MS, implying that the putative kinase (lmT4) and acyltranferase (lmT8) [22] are not functional for some reason.
Figure 2.
Biosynthetic gene cluster of lactomycins from Streptomyces sp. ACT232.
Table 2.
Proposed funtion of ORFs in lactomycin biosynthetic gene cluster from Streptomyces sp. ACT232.
2.4. Biological Activity
Lactomycin A, B, and C inhibit cathepsin B with IC50 values of 4.5 μM, 0.8 μM, and 1.6 μM, respectively. Their inhibitory activities are moderate compared with those of known cathepsin B inhibitors, such as E-64, which are active at a nM range [23]. Additionally, the cytotoxic activity of lactomycins A–C was examined, but none of them showed activity against HeLa cells at concentrations lower than 40 μM.
3. Discussion
Phoslactomycins [10,11,12,13] and leustroducsins [14,15] are polyketides that possess α,β-unsaturated δ-lactone and cyclohexane moieties at both ends, which are tethered through a highly unsaturated chain with a phosphate ester. This class of compounds shows a plethora of biological activities, such as antifungal activity, inhibitory activity against protein phosphatase 2A, increasing the level of cytokines such as colony stimulating factors, and induction of myeloid differentiation in HL-60 cells. However, their inhibitory activity against cathepsin B has not been reported so far. We noticed the structural similarity of lactomycins and leptomycins [24,25], polyketides with an α,β-unsaturated δ-lactone moiety and an unsaturated chain. Leptomycins are potent cytotoxins and the active site is the α,β-unsaturated δ-lactone moiety. However, lactomycins do not show cytotoxic activity, demonstrating that the presence of α,β-unsaturated δ-lactone is not sufficient for cytotoxicity.
4. Materials and Methods
4.1. General Experimental Procedures
UV spectra were measured on a Shimadzu Biospec 1600 (Shimadzu, Kyoto, Japan). NMR spectra were recorded on a JEOL alpha 600 NMR spectrometer (JEOL, Tokyo, Japan). Chemical shifts were referenced to a solvent peak: δH 2.49 and δC 40.0 (DMSO-d6) and δH 3.30 and δC 49.0 (CD3OD). HRESI mass spectra were measured on a JEOL JMS-T100LC (JEOL, Tokyo, Japan). HPLC purification was carried out on a Shimadzu LC 20AT (Shimadzu, Kyoto, Japan) with an SCL-10 Avp controller and an SPD-10Avp detector.
4.2. Collection and Identification of the Microorganism
Deep-sea sediments were collected by the manned submersible “SHINKAI 2000” system off Hatsu-shima, Sagami-Bay, Japan, at a depth of 1174 m, in December 2001. The sediment sample was stored in a sterilized sampler, frozen with liquid nitrogen, and transported to the laboratory, where it was kept frozen until processed. The Streptomyces sp. ACT232 was isolated from this sample. The taxonomy of the strain was determined by 16S rRNA phylogenetic analysis using 27F and 1492R primers, and the sequence was deposited in the DNA Data Bank of Japan (DDBJ, accession no. AB968434).
4.3. Fermentation, Extraction, and Isolation
Streptomyces sp. ACT232 was cultured in 120 × 500 mL Erlenmeyer flasks, each containing 250 mL of ISP2 medium (yeast extract 1.0 g, malt extract 2.5 g, glucose 1.0 g) at 28 °C on rotary shakers at 150 rpm. After 10 days of culture, the mycelia were separated by vacuum filtration. Resins (Amberlite XAD 16N, Sigma-Aldrich, MO, USA) were added to the filtrate to allow the adsorption of metabolites. The acetone extract of both the mycelia and resins was combined and then partitioned between n-BuOH and H2O. The extract was concentrated in vacuo and separately subjected to ODS flash column chromatography eluting with 20%, 40%, 60%, 80%, and 100% (v/v) MeOH in H2O. The fractions that eluted with 40% and 60% MeOH were purified by ODS-HPLC (Cosmosil MSII φ 4.6 × 250 mm, Nacalai tesque, Tokyo, Japan) with gradient elution from 45% to 60% aqueous MeOH with 0.2% AcOH to afford lactomycins A (1.5 mg) and C (0.3 mg). The fraction that eluted with 80% MeOH was purified by ODS-HPLC with gradient elution from 10% to 40% aqueous MeOH with 0.5% AcOH to afford lactomycins B (1.5 mg)
Lactomycin A (1): white solid; [α]20D (c 0.09, MeOH) +30; UV (MeOH) λmax 230 nm (log ε 4.12); 1H and 13C NMR, see Table 1; HRESIMS m/z 473.2510 [M + Na]+ (calcd. for C25H38NaO7, 473.2515).
Lactomycin B (2): white solid; [α]24D (c 0.06, MeOH) +32; UV (MeOH) λmax 230 nm (log ε 2.92); 1H and 13C NMR, see Table 1; HRESIMS m/z 472.2656 [M + Na]+ (calcd. for C25H39N Na O6, 472.2675).
Lactomycin C (3): white solid; [α]20D (c 0.02, MeOH) +1; UV (MeOH) λmax 230 nm (log ε 3.80); 1H and 13C NMR, see Table 1; HRESIMS m/z 443.2432 [M + Na]+ (calcd. for C24H36NaO6, 443.2410).
4.4. Identification of the Biosynthetic Gene Cluster
Streptomyces sp. ACT232 was cultured in ISP2 medium for three days at 30 °C with agitation and aeration. The genomic DNA was extracted from mycelia and isolated using QIAGEN Genomic-tip 20/G. The genome of Streptomyces sp. ACT232 was sequenced by Ilumina Hiseq to afford the data set consisting of 53,907,834 single 100 bp reads. These reads were subjected to de novo assembly with CLC Genomics Workbench (ver8.5) to afford 15 contigs as draft genome sequences. The function of each gene was identified by antiSMASH 3.0 [17] and Blast searches. (accession number LC364194 and LC365285)
4.5. Cathepsin B Inhibitory Assay
A Cathepsin B inhibitory assay was performed according to a modification of the method of Hiwasa et al. [26] The enzyme (cathepsin B from bovine spleen, Sigma C6286) was stocked at 1 unit/mL in 50 mM MES pH 6.0 and 0.1% Brij-35. The enzyme solution was diluted by 100 times with the buffer before use. The mixture of 4 μL test sample solution, 100 μL of the enzyme solution, and 50 μL of 25 μM fluorescent substrate (Z-Arg-Arg-AMC, Peptide Institute, Inc.) in DMSO was incubated at 37 °C for 30 min. The fluorescence of the liberated AMC was measured with an excitation at 345 nm and emission at 440 nm.
Supplementary Materials
The following are available online at http://www.mdpi.com/1660-3397/16/2/70/s1, 1D and 2D NMR spectra of lactomycins A (1)–C (3).
Acknowledgments
This work was partly supported by JSPS KAKENHI Grant Numbers 25252037, 16H04980 from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Author Contributions
K.T., S.O. and S.M. conceived and designed the experiments; Y.S., R.R.C. and Y.H. performed the experiments; K.T., K.Y. and S.A. performed genome analyses; Y.N. isolated the Streptomyces sp. ATC232; K.T. and S.M. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Munro, M.H.G.; Blunt, J.W.; Dumdei, E.J.; Hickford, S.J.H.; Lill, R.E.; Li, S.; Battershill, C.N.; Duckworth, A.R. The discovery and development of marine compounds with pharmaceutical potential. J. Biotechnol. 1999, 70, 15–25. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Irie, R.; Takda, K.; Ise, Y.; Ohtsuka, S.; Okada, S.; Gustafson, K.; Matsunaga, S. Structure revision of poecillastrin C and the absolute configuration of the β-hydroxyaspartic acid residue. Org. Lett. 2017, 19, 5395–5397. [Google Scholar] [CrossRef] [PubMed]
- Gondi, C.S.; Rao, J.S. Cathepsin B as a cancer target. Expert Opin. Ther. Targets 2013, 17, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Takada, K.; Nogi, Y.; Okada, S.; Matsunaga, S. Lower homologues of ahpatinin, aspartic protease inhibitores, from a marine Streptomyces sp. J. Nat. Prod. 2014, 77, 1749–1752. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kurihara, S.; Oikawa, T.; Ohkawa, N.; Shimazaki, N.; Sasagawa, K.; Kobayashi, T.; Kohama, T.; Asai, F.; Shiraishi, A.; Sugimura, Y. Preparation of leustroducsin H and the structure-activity relationship of its derivatives. J. Antibiot. 1995, 48, 1518–1520. [Google Scholar] [CrossRef] [PubMed]
- Trost, B.M.; Knopf, J.D.; Brindle, C.S. Synthetic strategies employed for the construction of fostriecin and related natural products. Chem. Rev. 2016, 116, 15035–15088. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, C.R.; Trager, W.F. Absolute configuration of acenocoumarin. J. Med. Chem. 1979, 22, 1122–1124. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, S.; Nishikawa, S.; Shimazu, A.; Seto, H. Studies on new phosphate ester antifungal antibiotics phoslactomycins. I. Taxonomy, fermentation, purification and biological activities. J Antibiot. 1989, 42, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, S.; Furihata, K.; Seto, H. Studies of new phosphate ester antifungal antibiotics phoslactomycins. II. Structure elucidation of phoslactomycins A to F. J. Antibiot. (Tokyo) 1989, 42, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Mizuhara, N.; Usuki, Y.; Ogita, M.; Fujita, K.I.; Kuroda, M.; Doe, M.; Iio, H.; Tanaka, T. Identification of phoslactomycin E as a metabolite inducing hyphal morphological abnormalities in Aspergillus fumigatus IFO 5840. J. Antibiot. (Tokyo) 2007, 60, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Fotso, S.; Graupner, P.; Xiong, Q.; Hahn, D.; Avila-Adame, C.; Davis, G. Phoslactomycins from Streptomyces sp. MLA1839 and their biological activities. J. Nat. Prod. 2013, 76, 1509–1513. [Google Scholar] [CrossRef] [PubMed]
- Kohama, T.; Nakamura, T.; Kinoshita, T.; Kaneko, I.; Shiraishi, A. Novel microbial metabolites of the phoslactomycins family induce production of colony-stimulating factors by bone marrow stromal cells. II. Isolation, physico-chemical properties and structure determination. J. Antibiot. (Tokyo) 1993, 46, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Kohama, T.; Maeda, H.; Sakai, J.I.; Shiraishi, A.; Yamashita, K. Leustroducsin B, a New Cytokine Inducer Derived from an Actinomycetes, Induces Thrombocytosis in Mice. J. Antibiot. (Tokyo) 1996, 49, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Simizu, S.; Teruya, T.; Nogawa, T.; Aono, H.; Ueki, M.; Uramoto, M.; Kobayashi, Y.; Osada, H. Deamino-hydroxy-phoslactomycin B, a biosynthetic precursor of phoslactomycin, induces myeloid differentiation in HL-60 cells. Biochem. Biophys. Res. Commun. 2009, 383, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. AntiSMASH 3.0―A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, N.; Kim, B.S.; Sekiyama, Y.; Osada, H.; Reynolds, K.A. Enhancement and selective production of phoslactomycin B, a protein phosphatase IIa inhibitor, through identification and engineering of the corresponding biosynthetic gene cluster. J. Biol. Chem. 2003, 278, 35552–35557. [Google Scholar] [CrossRef] [PubMed]
- Alhamadsheh, M.M.; Palaniappan, N.; DasChouduri, S.; Reynolds, K.A. Modular polyketide synthases and cis double bond formation: Establishment of activated cis-3-cyclohexylpropenoic acid as the diketide intermediate in phoslactomycin biosynthesis. J. Am. Chem. Soc. 2007, 129, 1910–1911. [Google Scholar] [CrossRef] [PubMed]
- Ghatge, M.S.; Reynolds, K.A. The plmS2-Encoded Cytochrome P450 Monooxygenase Mediates Hydroxylation of Phoslactomycin B in Streptomyces sp. Strain HK803. J. Bacteriol. 2005, 187, 7970–7976. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Zhao, J.; Liu, W.; Gao, J.F.; Tao, L.M.; Pan, H.X.; Tang, G.L. Identification of phoslactomycin biosynthetic gene clusters from Streptomyces platensis SAM-0654 and characterization of PnR1 and PnR2 as positive transcriptional regulators. Gene 2012, 509, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ghatge, M.S.; Palaniappan, N.; Alhamadsheh, M.M.; DiBari, J.; Reynolds, K.A. Application of a newly identified and characterized 18-O-acyltransferase in chemoenzymatic synthesis of selected natural and nonnatural bioactive derivatives of phoslactomycins. Appl. Environ. Microbiol. 2009, 75, 3469–3476. [Google Scholar] [CrossRef] [PubMed]
- Lecaille, F.; Kaleta, J.; Bpoemme, D. Human and parasitic papain-like cysteine proteases: Their role in physiology and pathology and recent developments in inhibitor design. Chem. Rev. 2002, 102, 4459–4488. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, T.; Seto, H.; Beppu, T. Leptomycins A and B, new antifungal antibiotics. I. Taxonomy of the producing strain and their fermentation, purification and characterization. J. Antibiot. (Tokyo) 1983, 36, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, T.; Seto, H.; Beppu, T. Leptomycins A and B, new antifungal antibiotics II Structure elucidation. J. Antibiot. (Tokyo) 1983, 36, 646–650. [Google Scholar] [PubMed]
- Hiwasa, T.; Fujita-Yoshigaki, J.; Shirouzu, M.; Koide, H.; Sawada, T.; Sakiyama, S.; Yokoyama, S. c-Ha-Ras mutants with point mutations in Gln-Val-Val region have reduced inhibitory activity toward cathepsin B. Cancer Lett. 1993, 69, 161–165. [Google Scholar] [CrossRef]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).