Diversity and Ecology of Marine Algicolous Arthrinium Species as a Source of Bioactive Natural Products

 ,
,
Abstract
1. Introduction
2. Results and Discussion
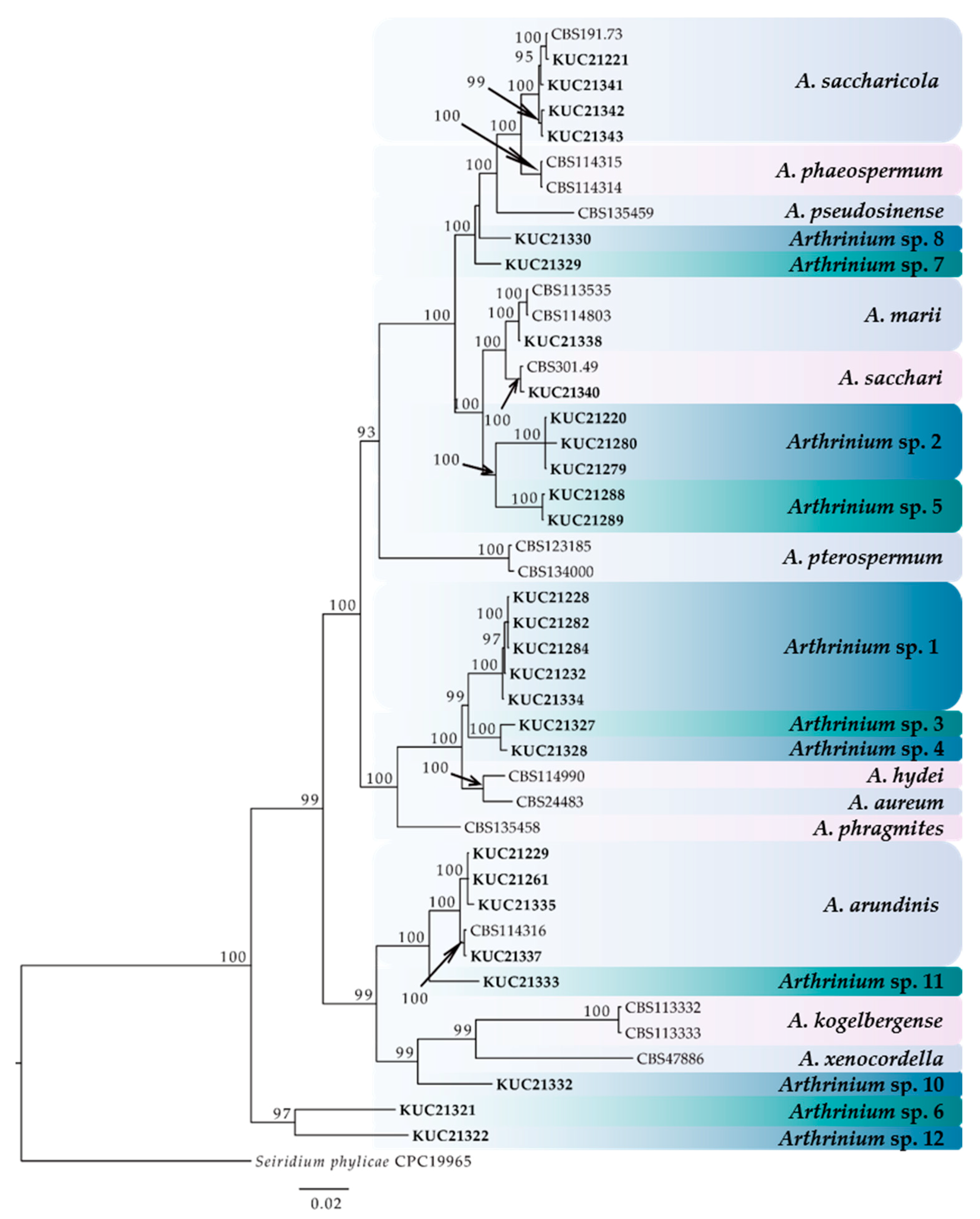
2.1. Identification and Multi-Gene Phylogeny
2.2. Marine Habitat of Arthrinium
2.3. Biological Activities of Marine Arthrinium spp.
2.3.1. Antioxidant Activity
2.3.2. Antifungal Activity
2.3.3. Tyrosinase Inhibition Activity
2.3.4. Quorum Sensing Inhibition Activity
2.4. Antioxidant Compound from Arthrinium sp. 10
3. Materials and Methods
3.1. Microorganisms
3.1.1. Fungal Resources
3.1.2. DNA Extraction, PCR, and Identification
3.1.3. Phylogenetic Analysis
3.2. Preparation of Fungal Extracts
3.3. Biological Assays
3.3.1. Antioxidant Assay
ABTS Radical-Scavenging Assay
DPPH Radical-Scavenging Assay
3.3.2. Antifungal Assay
3.3.3. Tyrosinase Inhibition Assay
3.3.4. Quorum Sensing Inhibition Assay
3.4. Isolation and Identification of the Bioactive Compound
3.4.1. Experimental Procedures
3.4.2. Bioassay-Guided Isolation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hong, J.-H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.M.; Lee, H.; Kim,, J.-J. Investigation of marine-derived fungal diversity and their exploitable biological activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.B. Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Sharma, R.; Kulkarni, G.; Sonawane, M.S.; Shouche, Y.S. A new endophytic species of arthrinium (apiosporaceae) from Jatropha podagrica. Mycoscience 2014, 55, 118–123. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Seifert, K.A.; Gams, W. The genera of hyphomycetes. Persoonia 2011, 27, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M. More Dematiaceous Hypomycetes; CABI Publishing: Kew, UK, 1976; p. 507. [Google Scholar]
- Wang, M.; Tan, X.-M.; Liu, F.; Cai, L. Eight new arthrinium species from china. MycoKeys 2018, 1–24. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 6969–6985. [Google Scholar] [CrossRef]
- Mousa, W.K.; Raizada, M.N. The diversity of anti-microbial secondary metabolites produced by fungal endophytes: An interdisciplinary perspective. Front. Microbiol. 2013, 4, 65. [Google Scholar] [CrossRef]
- Tsukada, M.; Fukai, M.; Miki, K.; Shiraishi, T.; Suzuki, T.; Nishio, K.; Sugita, T.; Ishino, M.; Kinoshita, K.; Takahashi, K.; et al. Chemical constituents of a marine fungus, Arthrinium sacchari. J. Nat. Prod. 2011, 74, 1645–1649. [Google Scholar] [CrossRef]
- Ebada, S.S.; Schulz, B.; Wray, V.; Totzke, F.; Kubbutat, M.H.; Müller, W.E.; Hamacher, A.; Kassack, M.U.; Lin, W.; Proksch, P. Arthrinins a–d: Novel diterpenoids and further constituents from the sponge derived fungus arthrinium sp. Bioorg. Med. Chem. 2011, 19, 4644–4651. [Google Scholar] [CrossRef] [PubMed]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; Özkaya, F.C.; Ebrahim, W.; Singab, A.B.; Proksch, P. Spiroarthrinols a and b, two novel meroterpenoids isolated from the sponge-derived fungus arthrinium sp. Phytochem. Lett. 2017, 20, 246–251. [Google Scholar] [CrossRef]
- Wei, M.-Y.; Xu, R.-F.; Du, S.-Y.; Wang, C.-Y.; Xu, T.-Y.; Shao, C.-L. A new griseofulvin derivative from the marine-derived arthrinium sp. Fungus and its biological activity. Chem. Nat. Compd. 2016, 52, 1011–1014. [Google Scholar] [CrossRef]
- Bao, J.; Zhai, H.; Zhu, K.; Yu, J.-H.; Zhang, Y.; Wang, Y.; Jiang, C.-S.; Zhang, X.; Zhang, Y.; Zhang, H. Bioactive pyridone alkaloids from a deep-sea-derived fungus arthrinium sp. Ujnmf0008. Mar. Drugs 2018, 16, 174. [Google Scholar] [CrossRef]
- Bao, J.; He, F.; Yu, J.-H.; Zhai, H.; Cheng, Z.-Q.; Jiang, C.-S.; Zhang, Y.; Zhang, Y.; Zhang, X.; Chen, G. New chromones from a marine-derived fungus, arthrinium sp., and their biological activity. Molecules 2018, 23, 1982. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Ju, Z.; Wan, J.; Liao, S.; Lin, X.; Zhang, T.; Zhou, X.; Chen, H.; Tu, Z. Cytotoxic cytochalasins from marine-derived fungus Arthrinium arundinis. Planta Med. 2015, 81, 160–166. [Google Scholar] [CrossRef]
- Park, M.S.; Oh, S.-Y.; Lee, S.; Eimes, J.A.; Lim, Y.W. Fungal diversity and enzyme activity associated with sailfin sandfish egg masses in korea. Fungal Ecol. 2018, 34, 1–9. [Google Scholar] [CrossRef]
- Park, J.Y.; Cho, J.K.; Son, M.H.; Kim, K.M.; Han, K.H.; Park, J.M. Artificial spawning behavior and development of eggs, larvae and juveniles of the red spotted grouper, epinephelus akaara in korea. Dev. Reprod. 2016, 20, 31–40. [Google Scholar] [CrossRef]
- Miao, L.; Kwong, T.F.N.; Qian, P.-Y. Effect of culture conditions on mycelial growth, antibacterial activity, and metabolite profiles of the marine-derived fungus arthrinium c.F. Saccharicola. Appl. Microbiol. Biotechnol. 2006, 72, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-Y.; Cai, Y.-Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Wang, H.-M.D.; Chen, C.-C.; Huynh, P.; Chang, J.-S. Exploring the potential of using algae in cosmetics. Bioresour. Technol. 2015, 184, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Machu, L.; Misurcova, L.; Vavra Ambrozova, J.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.; Doyle, S.; Murphy, R. Filamentous fungi as a source of natural antioxidants. Food Chem. 2015, 185, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Bartasiute, A.; Westerink, B.H.; Verpoorte, E.; Niederländer, H.A. Improving the in vivo predictability of an on-line hplc stable free radical decoloration assay for antioxidant activity in methanol–buffer medium. Free Radic. Biol. Med. 2007, 42, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Skriptsova, A.V. Fucoidans of brown algae: Biosynthesis, localization, and physiological role in thallus. Russ. J. Mar. Biol. 2015, 41, 145–156. [Google Scholar] [CrossRef]
- Raghukumar, S. Fungi in Coastal and Oceanic Marine Ecosystems; Springer: New York, NY, USA, 2017. [Google Scholar]
- Gomaa, M.; Hifney, A.F.; Fawzy, M.A.; Issa, A.A.; Abdel-Gawad, K.M. Biodegradation of Palisada perforata (rhodophyceae) and sargassum sp.(phaeophyceae) biomass by crude enzyme preparations from algicolous fungi. J. Appl. Phycol. 2015, 27, 2395–2404. [Google Scholar] [CrossRef]
- Yap, P.S.X.; Yiap, B.C.; Ping, H.C.; Lim, S.H.E. Essential oils, a new horizon in combating bacterial antibiotic resistance. Open Microbiol. J. 2014, 8, 6–14. [Google Scholar] [CrossRef]
- Chang, T.-S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef]
- Kang, H.S.; Choi, J.H.; Cho, W.K.; Park, J.C.; Choi, J.S. A sphingolipid and tyrosinase inhibitors from the fruiting body of Phellinus linteus. Arch. Pharmacal. Res. 2004, 27, 742–750. [Google Scholar] [CrossRef]
- Li, X.; Kim, M.K.; Lee, U.; Kim, S.-K.; Kang, J.S.; Choi, H.D.; Son, B.W. Myrothenones a and b, cyclopentenone derivatives with tyrosinase inhibitory activity from the marine-derived fungus myrothecium sp. Chem. Pharm. Bull. 2005, 53, 453–455. [Google Scholar] [CrossRef]
- Chen, J.S.; Wei, C.I.; Marshall, M.R. Inhibition-mechanism of kojic acid on polyphenol oxidase. J. Agric. Food Chem. 1991, 39, 1897–1901. [Google Scholar] [CrossRef]
- Olson, M.E.; Ceri, H.; Morck, D.W.; Buret, A.G.; Read, R.R. Biofilm bacteria: Formation and comparative susceptibility to antibiotics. Can. J. Vet. Res. 2002, 66, 86–92. [Google Scholar] [PubMed]
- Rasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, S.A.; Allain, E.J.; Michels, J.J.; Stearns, G.W.; Kelly, R.F.; McCoy, W.F. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Appl. Environ. Microbiol. 2001, 67, 3174–3179. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, Y.H.; Seo, Y.W.; Park, S. Quorum sensing inhibitors from the red alga, Ahnfeltiopsis flabelliformis. Biotechnol. Bioprocess Eng. 2007, 12, 308–311. [Google Scholar] [CrossRef]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef] [PubMed]
- Baldry, M.; Nielsen, A.; Bojer, M.S.; Zhao, Y.; Friberg, C.; Ifrah, D.; Heede, N.G.; Larsen, T.O.; Frøkiær, H.; Frees, D.; et al. Norlichexanthone reduces virulence gene expression and biofilm formation in Staphylococcus aureus. PLoS ONE 2016, 11, e0168305. [Google Scholar] [CrossRef]
- Skandamis, P.N.; Nychas, G.-J.E. Quorum sensing in the context of food microbiology. Appl. Environ. Microbiol. 2012, 78, 5473–5482. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef]
- Chen, L.; Fang, Y.; Zhu, T.; Gu, Q.; Zhu, W. Gentisyl alcohol derivatives from the marine-derived fungus Penicillium terrestre. J. Nat. Prod. 2008, 71, 66–70. [Google Scholar] [CrossRef]
- Nenkep, V.N.; Yun, K.; Li, Y.; Choi, H.D.; Kang, J.S.; Son, B.W. New production of haloquinones, bromochlorogentisylquinones a and b, by a halide salt from a marine isolate of the fungus phoma herbarum. J. Antibiot. 2010, 63, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, J.H.; Lee, C.H.; Kwon, H.J. Gentisyl alcohol, an antioxidant from microbial metabolite, induces angiogenesis in vitro. J. Microbiol. Biotechnol. 2006, 16, 475–479. [Google Scholar]
- Kim, J.; Kim, D.; Kim, M.; Kwon, H.; Oh, T.; Lee, C. Gentisyl alcohol inhibits apoptosis by suppressing caspase activity induced by etoposide. J. Microbiol. Biotechnol. 2005, 15, 532–536. [Google Scholar]
- Lernoux, M.; Schnekenburger, M.; Dicato, M.; Diederich, M. Anti-cancer effects of naturally derived compounds targeting histone deacetylase 6-related pathways. Pharmacol. Res. 2018, 129, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Zwick, V.; Allard, P.M.; Ory, L.; Simões-Pires, C.A.; Marcourt, L.; Gindro, K.; Wolfender, J.L.; Cuendet, M. Uhplc-ms-based hdac assay applied to bio-guided microfractionation of fungal extracts. Phytochem. Anal. 2017, 28, 93–100. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Son, B.-W. Antibacterial and radical scavenging epoxycyclohexenones and aromatic polyols from a marine isolate of the fungus aspergillus. Nat. Prod. Sci. 2005, 11, 136–138. [Google Scholar]
- Gupta, S.; Kaul, S.; Singh, B.; Vishwakarma, R.A.; Dhar, M.K. Production of gentisyl alcohol from Phoma herbarum endophytic in Curcuma longa L. And its antagonistic activity towards leaf spot pathogen Colletotrichum gloeosporioides. Appl. Biochem. Biotechnol. 2016, 180, 1093–1109. [Google Scholar] [CrossRef] [PubMed]
- Malak, L.G.; Ibrahim, M.A.; Bishay, D.W.; Abdel-baky, A.M.; Moharram, A.M.; Tekwani, B.; Cutler, S.J.; Ross, S.A. Antileishmanial metabolites from Geosmithia langdonii. J. Nat. Prod. 2014, 77, 1987–1991. [Google Scholar] [CrossRef]
- Ali, T.; Inagaki, M.; Chai, H.-B.; Wieboldt, T.; Rapplye, C.; Rakotondraibe, L.H. Halogenated compounds from directed fermentation of penicillium concentricum, an endophytic fungus of the liverwort Trichocolea tomentella. J. Nat. Prod. 2017, 80, 1397–1403. [Google Scholar] [CrossRef]
- Smetanina, O.; Kalinovskii, A.; Khudyakov, Y.V.; Moiseenko, O.; Pivkin, M.; Menzorova, N.; Sibirtsev, Y.T.; Kuznetsova, T. Metabolites of the marine fungus Asperigillus varians kmm 4630. Chem. Nat. Compd. 2005, 41, 243–244. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. Its primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. Its as an environmental DNA barcode for fungi: An in silico approach reveals potential pcr biases. BMC Microbiol 2010, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rdna its2 types within a monophyletic lineage of the fungus fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the pcr to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [PubMed]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Maddison, D.R.; Maddison, W.P. Macclade 4: Analysis of Phylogeny and Character Evolution. Version 4.08 a; Sinauer Associates: Sunderland, MA, USA, 2005. [Google Scholar]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nylander, J.A.A. Mrmodeltest v2. Program Distributed by the Author; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Kim, J.D.; Han, J.W.; Hwang, I.C.; Lee, D.; Kim, B.S. Identification and biocontrol efficacy of Streptomyces miharaensis producing filipin iii against Fusarium wilt. J. Basic Microbiol. 2012, 52, 150–159. [Google Scholar] [CrossRef]
- Lai, H.Y.; Lim, Y.Y.; Tan, S.P. Antioxidative, tyrosinase inhibiting and antibacterial activities of leaf extracts from medicinal ferns. Biosci. Biotechnol. Biochem. 2009, 73, 1362–1366. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.E.; Han, J.W.; Jeon, B.J.; Kim, B.S. Efficacies of quorum sensing inhibitors, piericidin a and glucopiericidin a, produced by Streptomyces xanthocidicus kpp01532 for the control of potato soft rot caused by Erwinia carotovora subsp. atroseptica. Microbiol. Res. 2016, 184, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Ikawa, T.; Nisizawa, K. Pathway of mannitol formation during photosynthesis in brown algae. PCPhy 1969, 10, 425–440. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Fungal Identity | Strain ID | Isolation Source | GenBank Accession Number | Antioxidant Activity (IC50, μg/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | TUB | EF-1α | ABTS a | DPPH b | |||
| A. arundinis | KUC21229 | Sargassum fulvellum | KT207747 | MH498470 | MH498512 | MH544684 | 40.55 | >1000 |
| KUC21261 | Sargassum fulvellum | KT207779 | MH498469 | MH498511 | MH544683 | 52.33 | >1000 | |
| KUC21335 | Unknown seaweed | MH498552 | N.D. | MH498510 | N.D. | 64.89 | N.D. | |
| KUC21337 | Beach sand | MH498551 | MH544659 | MH498509 | MH544682 | >100 | >1000 | |
| A. marii | KUC21338 | Unknown seaweed | MH498549 | MH498467 | MH498507 | MH544681 | 74.77 | N.D. |
| A. sacchari | KUC21340 | Egg masses of Arctoscopus japonicus | MH498548 | MH498466 | MH498506 | MH544680 | 17.45 | 601.34 |
| A. saccharicola | KUC21221 | Sargassum fulvellum | KT207737 | KT207687 | KT207637 | MH544679 | 14.09 | 59.19 |
| KUC21341 | Egg masses of Arctoscopus japonicus | MH498547 | MH498465 | MH498505 | N.D. | >100 | N.D. | |
| KUC21342 | Egg masses of Arctoscopus japonicus | MH498546 | MH498464 | MH498504 | N.D. | 58.27 | N.D. | |
| KUC21343 | Egg masses of Arctoscopus japonicus | MH498545 | MH498463 | MH498503 | MH544678 | 38.81 | 744.32 | |
| Arthrinium sp. 1 | KUC21228 | Sargassum fulvellum | KT207746 | KT207696 | KT207644 | MH544677 | 55.29 | N.D. |
| KUC21232 | Sargassum fulvellum | KT207750 | KT207700 | KT207648 | MH544676 | >100 | >1000 | |
| KUC21282 | Sargassum fulvellum | MH498544 | MH498462 | MH498502 | MH544675 | 45.37 | >1000 | |
| KUC21284 | Sargassum fulvellum | MF615228 | MF615215 | MF615233 | MH544674 | 100.04 | >1000 | |
| KUC21334 | Egg masses of Arctoscopus japonicus | MH498543 | MH544661 | MH498501 | MH544673 | 50.89 | >1000 | |
| Arthrinium sp. 2 | KUC21220 | Sargassum fulvellum | KT207736 | KT207786 | KT207636 | MH544672 | 23.23 | >1000 |
| KUC21279 | Sargassum fulvellum | MF615229 | MF615216 | MF615234 | MH544671 | 41.71 | >1000 | |
| KUC21280 | Sargassum fulvellum | MH498542 | MH544660 | MH498500 | N.D. | 61.43 | >1000 | |
| Arthrinium sp. 3 | KUC21327 | Egg masses of Arctoscopus japonicus | MH498541 | MH498461 | MH498499 | MH544670 | 10.32 | >1000 |
| Arthrinium sp. 4 | KUC21328 | Unknown seaweed | MH498538 | MH498458 | MH498496 | MH544669 | 19.22 | >1000 |
| Arthrinium sp. 5 | KUC21288 | Unknown seaweed | MF615230 | MF615217 | MF615235 | MH544668 | 50.32 | >1000 |
| KUC21289 | Unknown seaweed | MF615226 | MF615213 | MF615231 | MH544668 | 61.63 | >1000 | |
| Arthrinium sp. 6 | KUC21321 | Unknown seaweed | MH498533 | MH498453 | MH498491 | N.D. | 46.98 | >1000 |
| Arthrinium sp. 7 | KUC21329 | Egg masses of Arctoscopus japonicus | MH498531 | MH498451 | MH498489 | MH544666 | 39.05 | >1000 |
| Arthrinium sp. 8 | KUC21330 | Egg masses of Arctoscopus japonicus | MH498530 | MH498450 | MH498488 | MH544665 | 72.56 | > 1000 |
| Arthrinium sp. 10 | KUC21332 | Egg masses of Arctoscopus japonicus | MH498524 | MH498444 | MH498482 | MH544664 | 14.87 | 73.22 |
| Arthrinium sp. 11 | KUC21333 | Agarum cribrosum | MH498520 | MH498440 | MH498478 | MH544663 | 32.62 | >1000 |
| Arthrinium sp. 12 | KUC21322 | Unknown seaweed | MH498515 | MH498435 | MH498473 | MH544662 | >100 | >1000 |
| Ascorbic acid * | 13.70 | 6.80 | ||||||
| Compound | Antioxidant Activity (IC50, μM) | |
|---|---|---|
| ABTS a | DPPH b | |
| Gentisyl alcohol (1) | 26.43 | 28.74 |
| Ascorbic acid * | 77.79 | 38.61 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, Y.M.; Kim, K.; Ryu, S.M.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Hong, J.-H.; Lim, Y.W.; Kim, C.; Kim, B.S.; et al. Diversity and Ecology of Marine Algicolous Arthrinium Species as a Source of Bioactive Natural Products. Mar. Drugs 2018, 16, 508. https://doi.org/10.3390/md16120508
Heo YM, Kim K, Ryu SM, Kwon SL, Park MY, Kang JE, Hong J-H, Lim YW, Kim C, Kim BS, et al. Diversity and Ecology of Marine Algicolous Arthrinium Species as a Source of Bioactive Natural Products. Marine Drugs. 2018; 16(12):508. https://doi.org/10.3390/md16120508
Chicago/Turabian StyleHeo, Young Mok, Kyeongwon Kim, Seung Mok Ryu, Sun Lul Kwon, Min Young Park, Ji Eun Kang, Joo-Hyun Hong, Young Woon Lim, Changmu Kim, Beom Seok Kim, and et al. 2018. "Diversity and Ecology of Marine Algicolous Arthrinium Species as a Source of Bioactive Natural Products" Marine Drugs 16, no. 12: 508. https://doi.org/10.3390/md16120508
APA StyleHeo, Y. M., Kim, K., Ryu, S. M., Kwon, S. L., Park, M. Y., Kang, J. E., Hong, J.-H., Lim, Y. W., Kim, C., Kim, B. S., Lee, D., & Kim, J.-J. (2018). Diversity and Ecology of Marine Algicolous Arthrinium Species as a Source of Bioactive Natural Products. Marine Drugs, 16(12), 508. https://doi.org/10.3390/md16120508